1. Introduction
Chestnuts are the main source of income for farmers in the upland areas of the north of Portugal. However, farmers are facing a quite complex situation due to a set of pests and diseases that weaken the trees, thereby reducing their productivity and, in some cases, causing their death. Currently, ink disease (
Phytophthora sp.pl.), chestnut blight (
Cryphonectria parasitica (Murrill) Barr.) and the Asian gall wasp (
Dryocosmus kuriphilus Yasumatsu) are the main health problems affecting chestnut trees [
1,
2,
3]. Notwithstanding this, chestnuts have maintained very good market prices [
4], which has led farmers to devote great attention and care to their crops, either replacing dead trees or establishing new orchards [
5].
Chestnut is grown all over the world as part of agroforestry systems with little phytotechnical intensification [
6]. In the mountainous areas of the north of Portugal, the lack of other crop options has raised chestnut to the status of the main crop, having been grown in monoculture and integrated into increasingly intensive farming systems, in a similar way to orchards of other important fruit trees [
7,
8,
9]. One of the practices that has received greater attention from producers is fertilisation, with trees currently being fertilised regularly [
5,
10,
11].
Crop fertilisation, being essential for obtaining high productivity in any species [
12,
13,
14], can also be associated with high risks of environmental contamination, especially the use of N fertilisers that can lead to the eutrophication of ground water [
15,
16] and the emission of greenhouse gases into the atmosphere, in particular, N oxides [
17,
18]. Thus, crop fertilisation must be managed judiciously, in order to apply the appropriate nutrient rates, thereby reducing the risk of environmental damage [
19,
20,
21,
22].
The prospects of a growing global population and the need to feed it, associated with the risks of environmental contamination, have led to the need to develop farming practices based on the concept of ecological intensification [
23,
24], which, in practice, means maintaining high productivity, but by using production factors in a more rational way. Thus, as with the main world crops, but also with chestnut, it is necessary to manage resources properly, using them in the smallest amounts necessary to maintain productivity.
Previous work carried out in NE Portugal has shown that in chestnut groves, nutrients are often below the lower limit of the sufficiency range [
25] and that trees generally tend to respond to fertiliser applications [
11,
26], although in some studies, they did not [
5]. However, there are still only a few studies on chestnut fertilisation, and the use of fertilisers is far from being optimized, with more data being required to establish adequate fertilisation programmes. It is therefore necessary to establish better guidelines for the fertilisation of these trees, to try to keep them healthy and productive, so that these magnificent ecosystems may persist, allowing man to continue to occupy these mountain territories which are showing concerning signs of depopulation [
27]. This study reports the results from a field experiment of chestnut fertiliser application using NPK fertilisers with different combinations of macronutrients, trying as best as possible to replicate the diversity of fertilisers found on the market, which farmers have access to. The objectives of the study are to understand better how these huge trees respond to fertiliser application so as to help farmers make better decisions when they need to acquire them.
2. Materials and Methods
2.1. Experimental Conditions
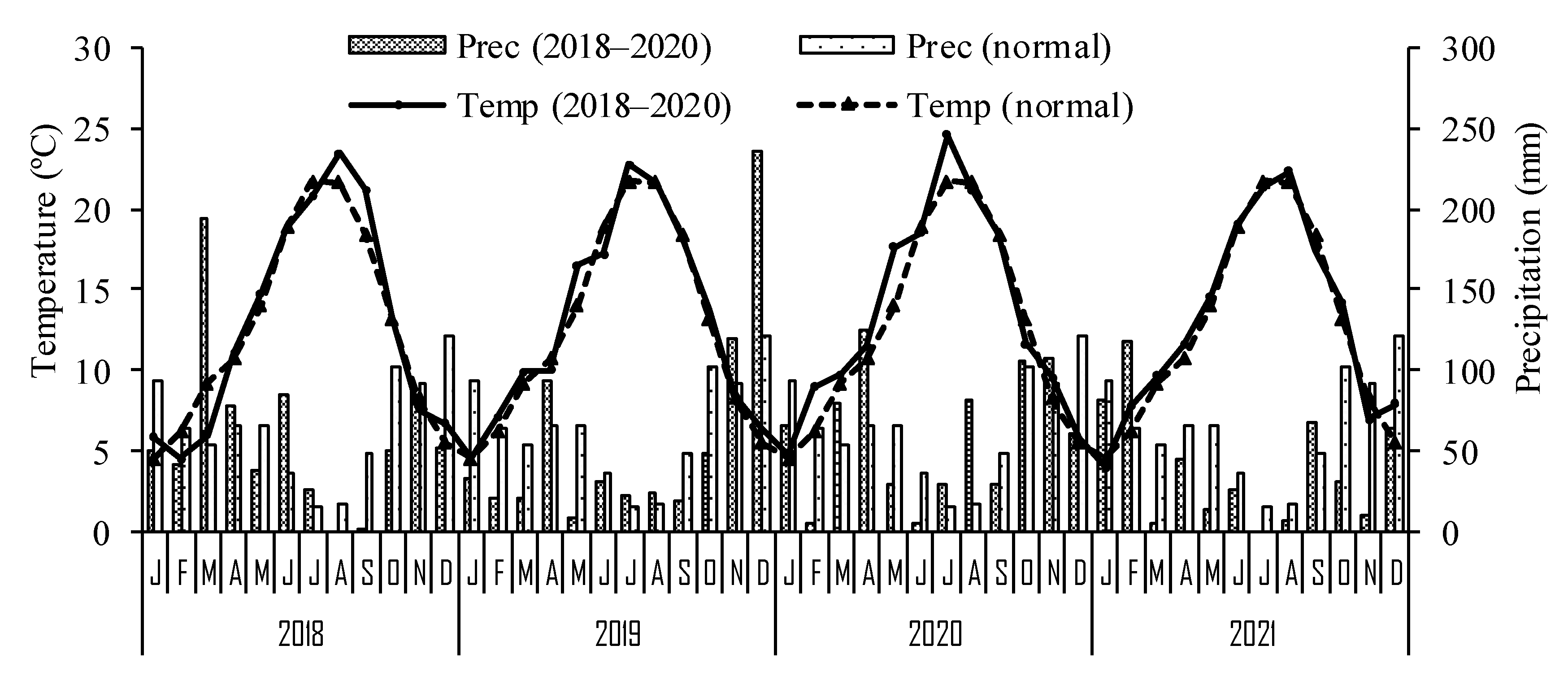
The field experiment took place in Vinhais (41°50′15.8″ N; 7°03′40.4″ W, 800 m above sea level), northeastern Portugal, in a 50-year-old chestnut orchard of the cultivar Judia with trees spaced at 10 m × 10 m. The region benefits from a warm-summer Mediterranean climate (Csb), according to the Köppen–Geiger classification [
28]. The annual mean temperature and the accumulated annual precipitation are 11.9 °C and 880 mm, respectively. Average monthly temperatures and precipitation of the climatological normal (1981–2010), together with those recorded during the experimental period (2018–2021), are presented in
Figure 1.
The soil where the chestnut orchard is planted is a Leptosol, sandy-loam textured. It is a very shallow soil (~0.20 m deep), which separates, determined from composite soil samples (n = 3) taken from the 0.0–0.2 m soil layer at the beginning of the study, were 11.8% clay, 17.3% silt and 70.9% sand. Soil organic C was low, pH acidic and extractable P and K were medium and very high, respectively. Some other soil properties determined at the beginning of the field trial are presented in
Table 1.
2.2. Experimental Design and Management of the Field Trial
Sixteen large-sized trees with similar, spherical canopies (~270 m3) were selected for the study. They were randomly distributed into four groups, corresponding to four fertilisation treatments, with four trees (replicates) in each treatment, in a completely randomized design. The treatments consisted of three compound NPK fertilisers with different levels of N, P and K, and an unfertilised control.
One of the treatments, named 7:14:14, consisted of the application of a 7:14:14 NPK compound fertiliser that doses 7% N (5% ammoniacal-N and 2% ureic-N), 14% P2O5 (11% water soluble) and 14% K2O. This fertiliser also contains 4% CaO, 2% MgO, 15% SO3, and 0.02% B. Another treatment named YA20:7:10, corresponds to the application of the commercial fertiliser Yara MilaTM Actyva 20:7:10, with 20% N (9.4% nitric-N, 10.6% ammoniacal-N), 7% P2O5 (25 to 30% as polyphosphates) and 10% K2O. The fertiliser also contains other important nutrients, namely S (10% SO3) and Mg (3% MgO). The third treatment, named YS13:11:21, consisted of the application of the NPK compound fertiliser Yara MilaTM Solán 13:11:21, which doses 13% N (5.5% nitric-N, 7.7% ammoniacal-N), 11% P2O5 (20 to 30% as polyphosphates) and 21% K2O. This fertiliser also contains relevant amounts of Mg (2% MgO) and B (0.2%).
All fertilisers were applied at a rate of 4 kg per tree (~400 kg ha−1). Thus, the YA20:7:10 treatment corresponded to an application of 80, 28 and 40 kg ha−1 of N, P2O5 and K2O, respectively; the 7:14:14 treatment to an application of 28, 56 and 56 kg ha−1 of N, P2O5 and K2O, respectively, and the YS13:11:21 to an application of 52, 44, 84 kg ha−1 of N, P2O5 and K2O, respectively. Similar fertilisations are commonly used by local farmers, and these particular fertilisers were chosen for the trial because they present a good variation in the levels of macronutrients.
The fertilisers were evenly applied beneath the canopy of the trees in the first week of April over the four years of the study and incorporated with a cultivator. The orchard was tilled a second time every year at the end of May to control the weeds. No further cropping practices were carried out in the orchard during the four years of the study.
2.3. Measurements in the Field
The effect of the treatments was assessed in the field by measuring the greenness of the leaves and by chlorophyll a fluorescence analysis.
The SPAD (Soil and Plant Analysis Development)-502 Plus chlorophyll meter (Spectrum Technologies, Inc., Aurora, IL, USA) was used to measure leaf greenness. SPAD-502 provides dimensionless readings, proportional to the chlorophyll content of the leaves, by measuring the transmittance of light through the leaves at 650 nm (red light, absorbed by chlorophyll) and 940 nm (infrared light, non-absorbed by chlorophyll). Each mean value was obtained after 30 individual readings taken around the crown on fully expanded young leaves.
Chlorophyll a fluorescence was assessed using the dark adaptation protocols with the OS-30p+ fluorometer (Opti-sciences, Inc., Hudson, NH, USA). FM, F0 and FV are, respectively, maximum, minimum and variable fluorescence from dark-adapted leaves. FV/FM is estimated as (FM − F0)/FM.
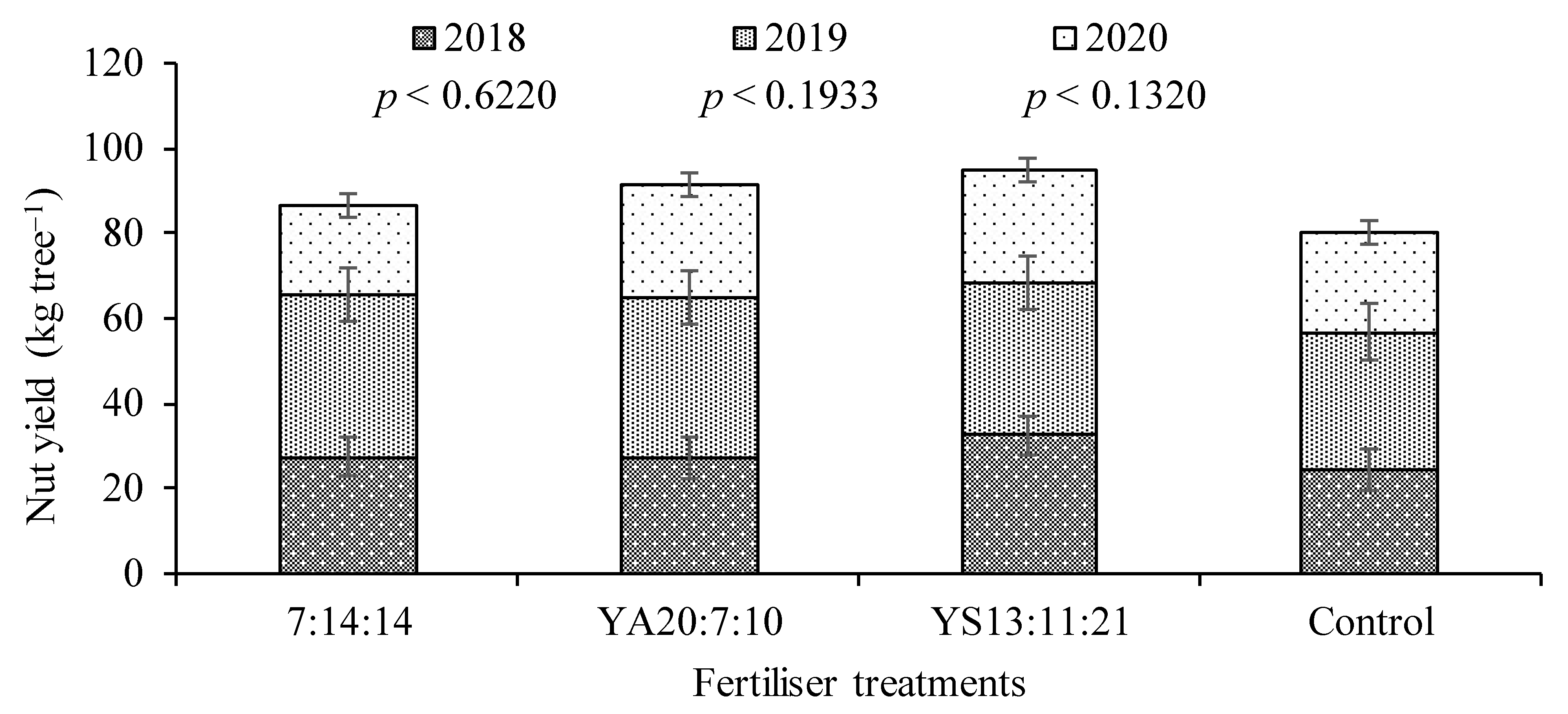
To harvest the chestnuts, it is necessary to wait for them to fall to the ground and then pick them up manually or mechanically. In this experiment, the fruits were harvested manually, in three passes during the autumn, to allow individual weighing per tree. In 2021, the COVID-19 pandemic restrictions did not allow the completion of harvest records, and therefore, only the values for 2018–2020 are available.
2.4. Soil and Plant Tissue Sampling and Analytical Determinations
Three composite samples were taken at the beginning of the experiment to characterize the experimental plot. The soil was sampled again in October 2021 to evaluate the effect of the treatments on soil properties. All soil samples taken to the laboratory were composite samples, taken at six different sampling points. Sampling was carried out in the 0.0–0.20 m soil layer, beneath the canopy of the trees, where the fertilisers had been applied.
Soil samples were oven-dried at 40 °C and sieved (2 mm mesh). Thereafter, the samples were analysed for pH (H
2O and KCl) (soil: solution, 1:2.5), cation-exchange capacity (ammonium acetate, pH 7.0), organic C (wet digestion, Walkley-Black method) and extractable P and K (Egner–Riehm method, ammonium lactate extract). Soil B was extracted by hot water and determined by the method of azomethine-H. For more details on these analytical procedures, the reader is referred to van Reeuwijk [
29]. The availability of other micronutrients (Cu, Fe, Zn, and Mn) in the soil was determined by atomic absorption spectrometry after extraction with ammonium acetate and EDTA, according to the method described by Lakanen and Erviö [
30]. Soil inorganic-N was determined in soil extracts prepared from 20 g of soil and 40 mL 2 M KCl. The suspension was shaken for 1 h and filtered through Watmann No. 42 filter paper. Nitrate and ammonium concentrations in the extracts were analysed in an UV–Vis spectrophotometer [
31].
By the end of July, in each of the four years, samples of young, fully developed leaves were taken for elemental analysis. Following each harvest, samples of 50 nuts per tree were randomly taken to evaluate their size and also for elemental analysis. After counting and weighing, the kernel was separated from shell and pellicle, and the two parts analysed separately. In April 2022, the spontaneous vegetation which had developed beneath the canopy of the trees was mowed to serve as a biological index of soil-available nutrients. The samples were collected by randomly placing a grid of 0.5 m × 0.5 m on the vegetation.
The samples of leaves, fruit kernels, shells and pellicles and spontaneous vegetation, were oven-dried at 70 °C until they reached a constant weight and ground (1 mm mesh). Elemental analyses of tissue samples were performed by Kjeldahl (N), colorimetry (B and P) and atomic absorption spectrophotometry (K, Ca, Mg, Fe, Mn, Cu, Zn) methods [
32] after tissue samples had been previously digested with nitric acid in a microwave.
2.5. Data Analysis
The data was analysed for normality and homogeneity of variance using the Shapiro–Wilk and Bartlett’s test, respectively. The analysis of variance was performed as a one-way ANOVA, using the Statistical Package for the Social Sciences (SPSS) version 25 (IBM Corporation, New York, NY, USA). When significant differences were found, the means were separated by the Tukey HSD post hoc test (α = 0.05).
4. Discussion
The annual and accumulated (2018–2020) chestnut yields did not vary significantly with fertilisation treatments. However, the average accumulated yield showed a clear tendency of reduction in the control in relation to the fertilised treatments. Chestnut trees are particularly large, with a huge perennial structure and canopy. In previous studies, it has already been observed that chestnut tends to respond poorly to fertilisers applied to the soil, probably due to the buffering effect that the perennial structure exerts in regulating the supply of nutrients for the growth of the aerial plant parts [
5].
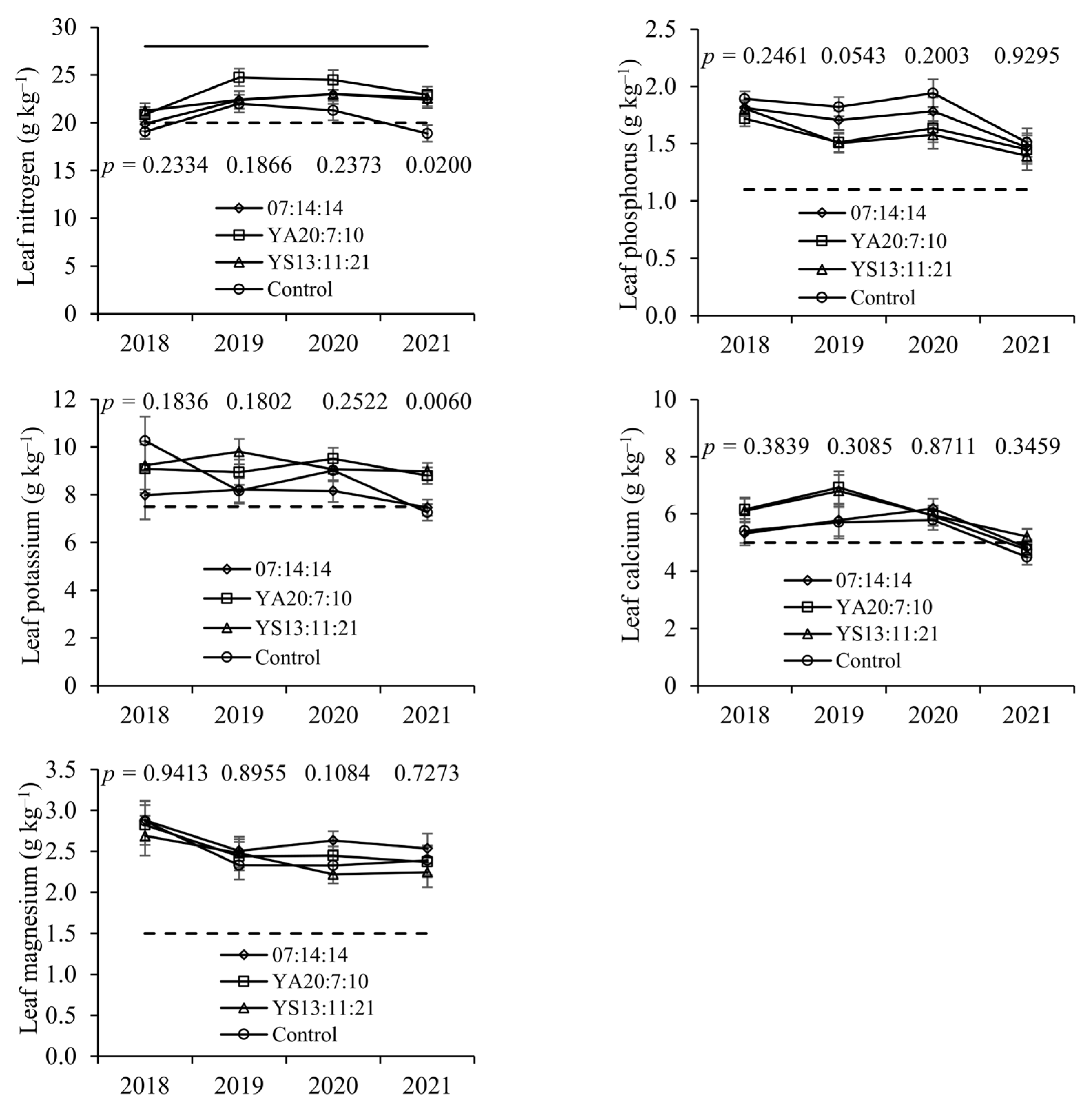
Leaf N concentrations tended to be higher in the fertilised treatments, which were more concentrated in N compared to the control, although significant differences only occurred on the last sampling date. In the control treatment, the values were close to the lower limit of the sufficiency range, having even fallen into the deficiency range on some sampling dates. The relevant structural role of N in plant tissues is undeniable [
33], and it is still recognized as the main nutrient that limits plant productivity in both natural ecosystems and cultivated fields [
19]. This result points to N as the most likely cause for the apparent drop in productivity in the control treatment. The SPAD values, which have been used mainly as an index of the N nutritional status of crops [
5,
29,
34], agreed with tissue N concentrations, and in 2021, significant differences were found between the YA20:7:10 treatment and the control. The F
V/F
M ratio, a widely used indicator of photoinhibition or other injuries at the PSII complexes [
35], followed the same trend as the N nutritional status indices, with values in the control being lower than in the treatment YA20:7:10 in the 2019 reading. However, the values never dropped below 0.78, which is the threshold limit below which most plants are considered to be under clear environmental stress [
26,
36,
37,
38]. Thus, the values of the maximum quantum efficiency of PSII also highlight the poor response of the photochemical reactions of photosynthesis of these huge trees to nutrient supply.
Leaf P concentrations did not vary significantly with treatments and always remained within the sufficiency range established for this species (1.1 to 3.0%) [
25,
39]. Initial soil P levels were at a level classified as medium (
Table 1), and in the region, it has been difficult to obtain a response of different crop species to P applications [
5,
40,
41]. In chestnut, the lack of response may be due to those reasons but also to the buffering effect of the perennial parts, already mentioned for N, and to a possible role of mycorrhizal fungi. Chestnut is recognized as a plant that establishes symbiotic relationships with several mycorrhizal fungi [
42]. One of the main benefits for mycorrhizal plants is the access to sparingly soluble P sources that non-mycorrhizal plants do not have [
43,
44,
45,
46,
47]. Thus, whatever the reason, the results seem to indicate a reduced importance of P in chestnut tree fertilisation programmes.
Although soil K levels increased with the application of fertilisers, no significant differences were observed in the K concentrations of the chestnut leaves. Leaf K concentrations varied greatly over the years and between treatments, although they generally remained within the sufficiency range. This pattern of K is common in shrub and tree species [
13,
21,
38,
48] and may be due to source/sink relationships and/or environmental constraints. Growing fruits are a primary sink for available K, the nutrient being remobilized from leaves [
33]. In chestnut, fruit growth coincides with the end of summer, a period in which there is often little soil moisture, which limits the movement of nutrients in the soil by mass flow and diffusion, making nutrient uptake difficult [
19]. In addition, the original levels of K in the soil were relatively high, which would have reduced the impact of applying K as a fertiliser. Finally, the buffering effect of the perennial tree structure may have moderated the effects of the fertiliser applications, as mentioned for N and P.
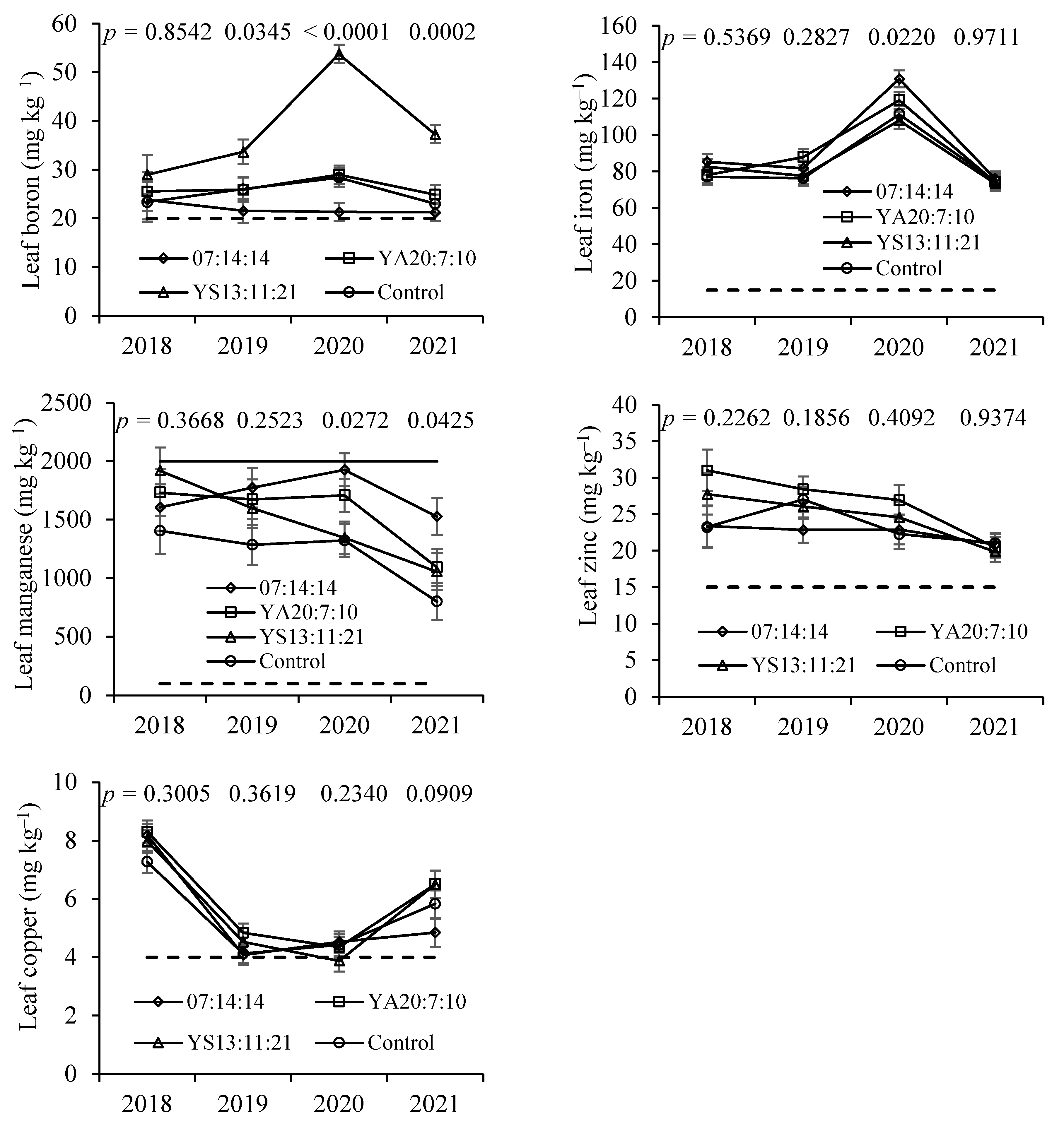
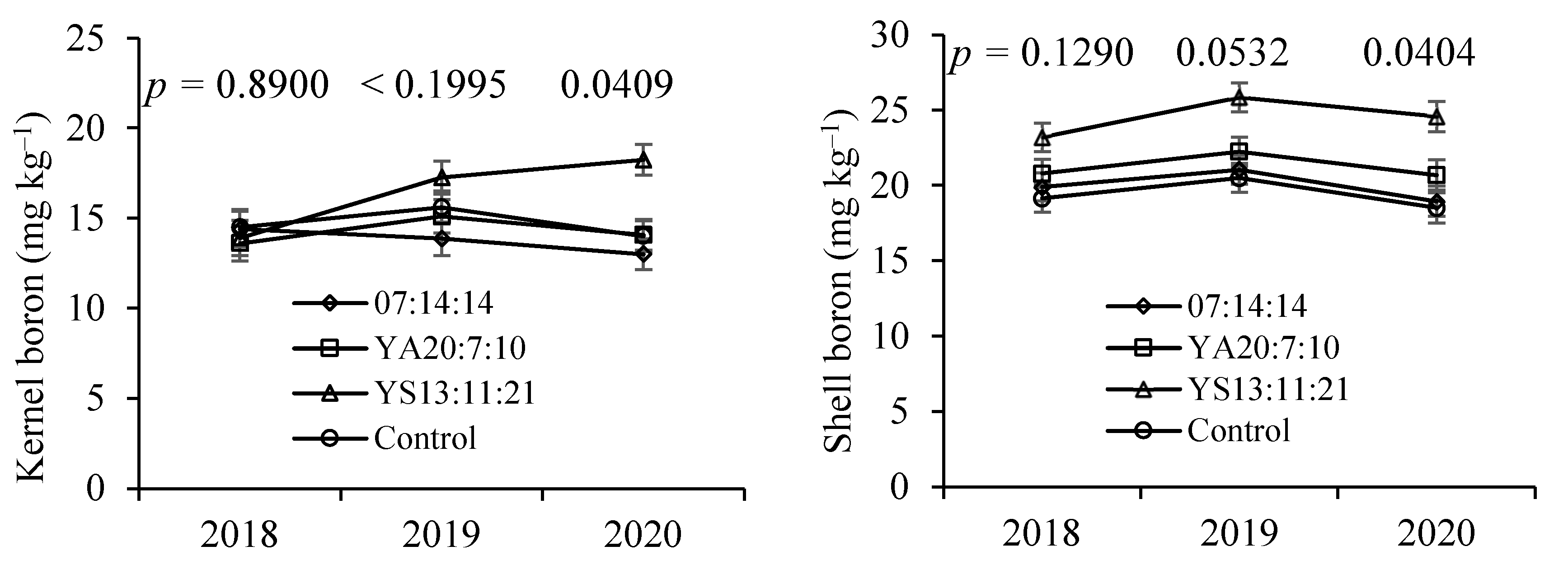
Tissue B levels differed between treatments on three dates, with YS13:11:21 fertiliser (B-rich) values being significantly higher than in the other treatments. Additionally in the fruits, B concentrations were the highest in the YA13:11:21 treatment, in particular, in the shell. Boron is very important in dicots, where it plays an important role in cell wall and membrane integrity, with these plants requiring greater amounts of B than monocots [
48,
49,
50]. In the region, dicots often respond to the application of B [
12,
41,
51]. In this field trial, however, tissue B levels were never below the sufficiency range even though they were close to the limit. Perhaps for this reason, B was not determinant in crop productivity, in contrast to what has been shown in other studies with chestnut [
11,
52,
53,
54].
In the control treatment, mean leaf Mn levels were lower than in the fertilised treatments, and the differences were statistically significant on the last sampling date. A tendency for lower Mn levels in soils was also observed in the control treatment. Soil pH in the control was higher than in the fertilised treatments, particularly in those that had a greater amount of applied N. Nitrification can decrease soil pH [
19], and the increase of available Mn levels in the fertilised treatments may have been a reflection of pH reduction [
19,
49]. Even so, leaf Mn levels never exceeded the upper limit of the sufficiency range, which is set at 2000 mg kg
−1 [
25,
39], so its effect on crop productivity must not have been relevant.
In the autumn, the availability of inorganic N in the soil as measured by hydrolysable NH
4+ and cold-extracted NO
3- was higher in the treatments with more N-concentrated fertilisers. This may indicate a greater risk of N loss through leaching and/or denitrification, since the rainy season follows, a precondition for the occurrence of these phenomena [
19]. However, in April, dry matter and tissue N concentrations in the spontaneous vegetation were also higher in the treatments that received more N as a fertiliser. These plants, which appear after the first autumn rains and develop during the winter, can play important roles by controlling soil erosion [
55,
56], increasing soil organic matter [
57,
58] and developing ecosystem biodiversity [
59,
60]. They also act as an N catch crop [
21,
61], justifying the promotion of their presence in orchard soils [
62]. This result also shows that the effect of N applications is easier to obtain in herbaceous vegetation than in a tree, probably due to the latter’s large perennial structure. It seems clear that in large trees, it is more difficult to get a response to fertilisation and therefore, more difficult to optimize a fertilisation programme. In trees, a dynamic optimization method should always be used [
63], by which, based on a given annual fertilisation plan, nutrient concentrations in leaves are monitored and fertiliser rates adjusted according to increasing (reduce fertilisation) or decreasing (increase fertilisation) concentrations of a particular nutrient being observed in leaves. This is a programme optimized for long-term monitoring and not just based on annual observations, which is the procedure currently used in fruit crops.
In Mediterranean climates, with rainfall concentrated in the winter, and the summer being particularly dry, in rainfed orchards, where there are no fertigation practices, there is only one window of opportunity to apply fertilizers, which is in early spring, just before the regrowth of vegetation. If applying earlier, there is a risk of loss of mobile nutrients, such as N, by leaching and denitrification, whereas if applying later, there is a risk of loss of effectiveness due to reduced soil moisture [
21]. In addition, slow and controlled release fertilizers tend to be less effective, as they delay nutrient availability for the summer, when the opportunity for root uptake is low [
64].



 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}