
Different Quality Classes of Decomposing Plant Residues Influence Dissolved Organic Matter Stoichiometry Which Results in Different Soil Microbial Processing


Abstract
:1. Introduction
2. Materials and Methods
2.1. Framework of Data Used
2.2. Study Site and Soil
2.3. Treatments and Experimental Design
2.4. Soil Collection, and Extraction and Analysis for DOC and DN
2.5. Statistical Analysis
3. Results
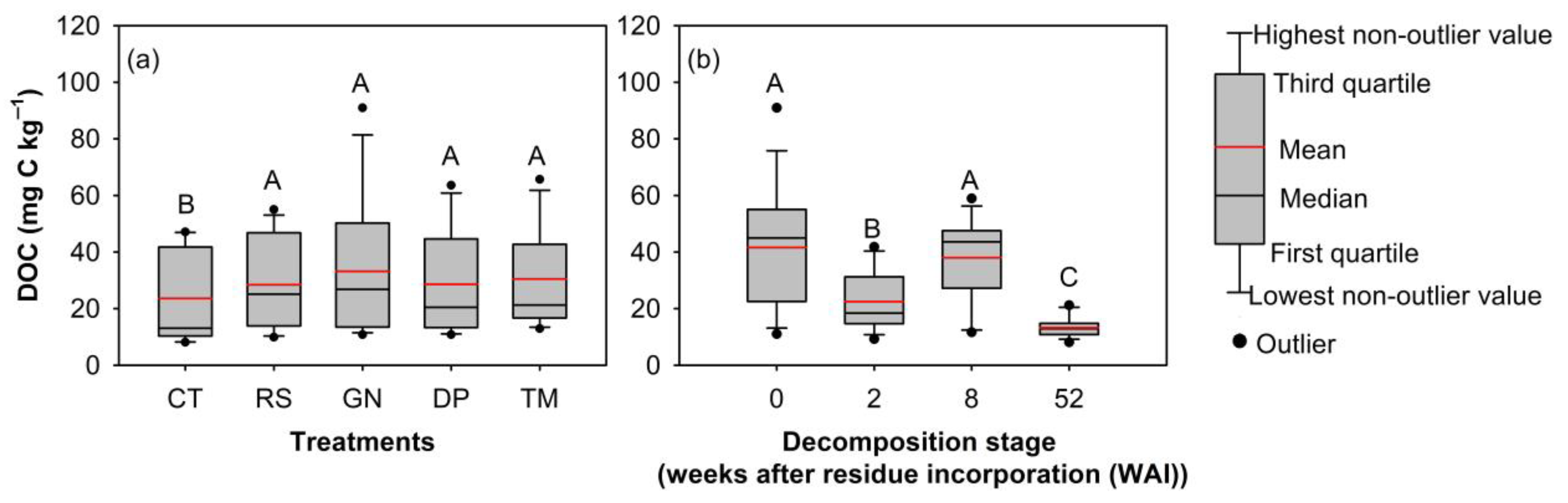
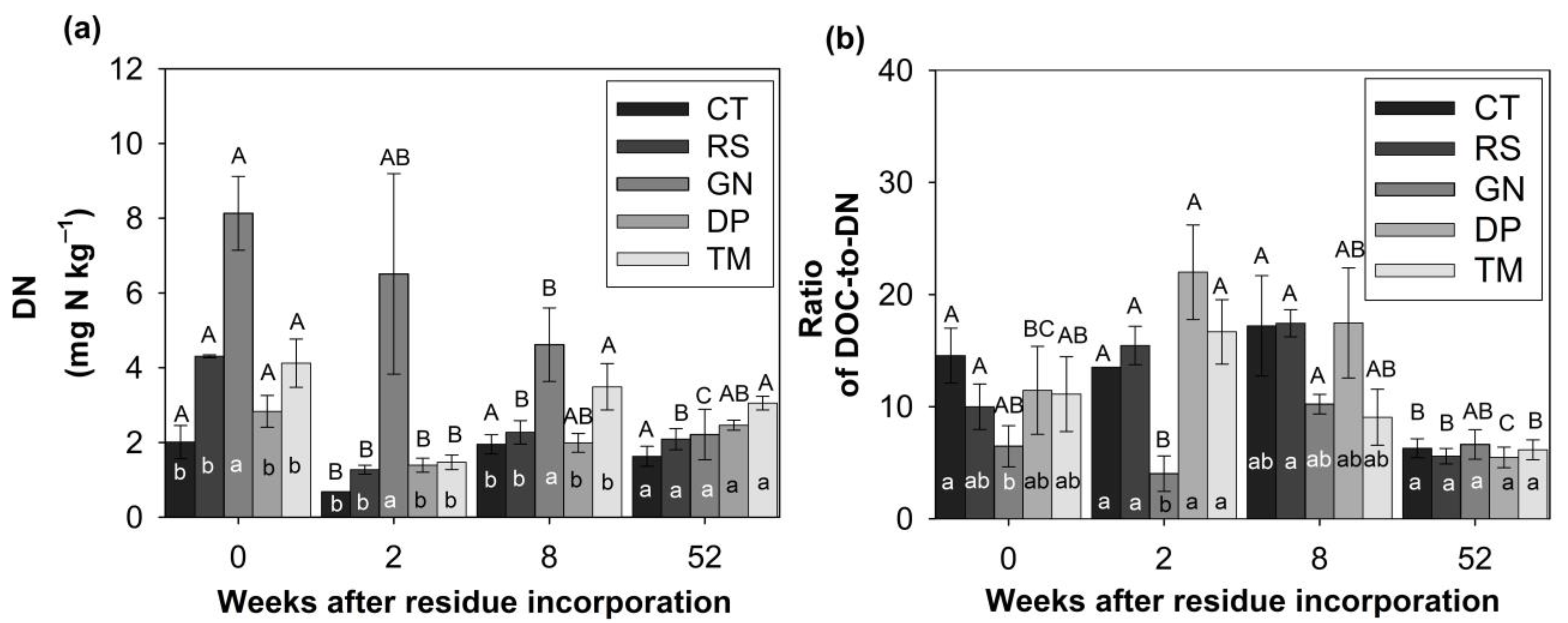
3.1. Organic Residue Quality Influences Production of DOM and the DOC-to-DN Ratio during Different Stages of Decomposition
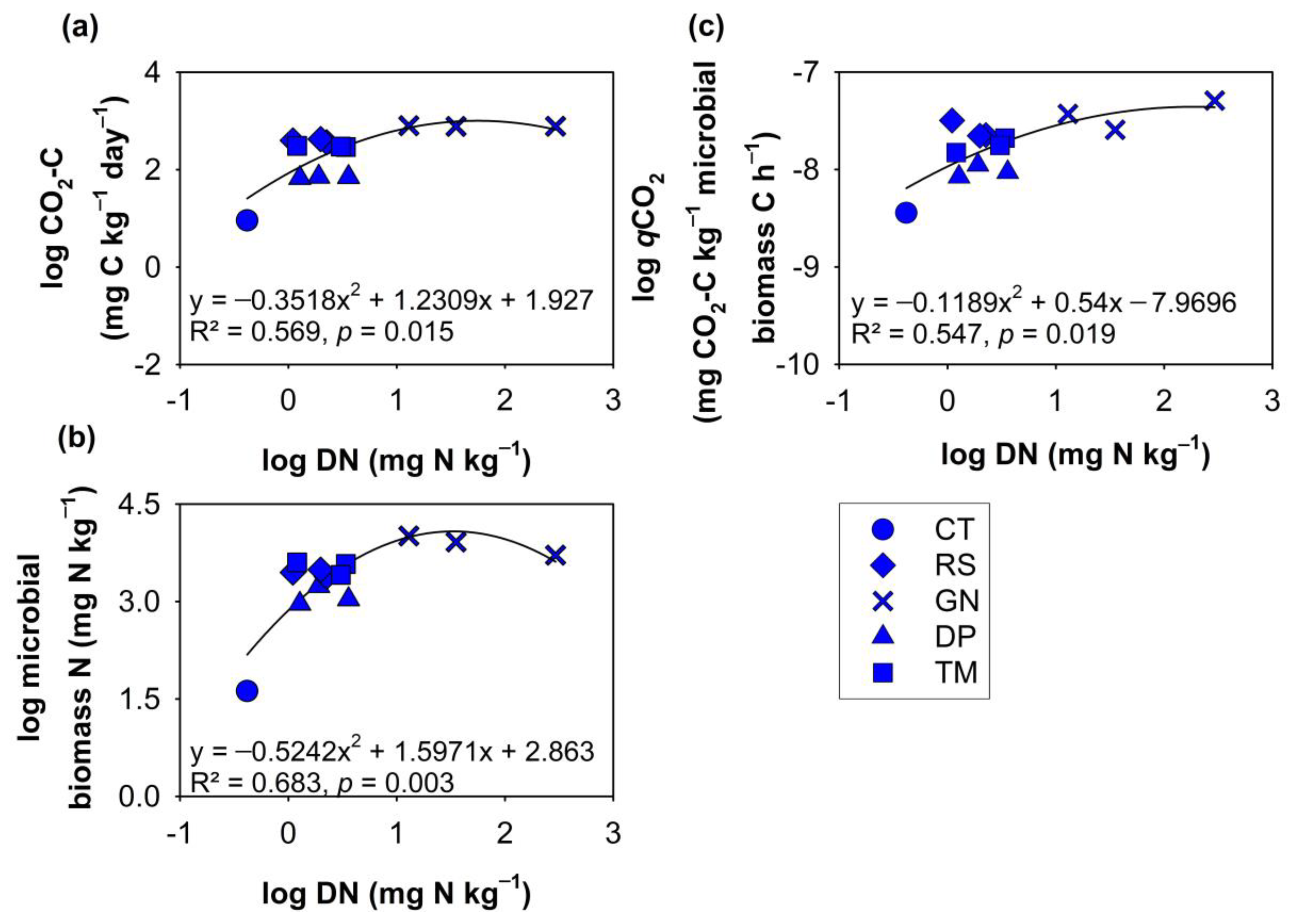
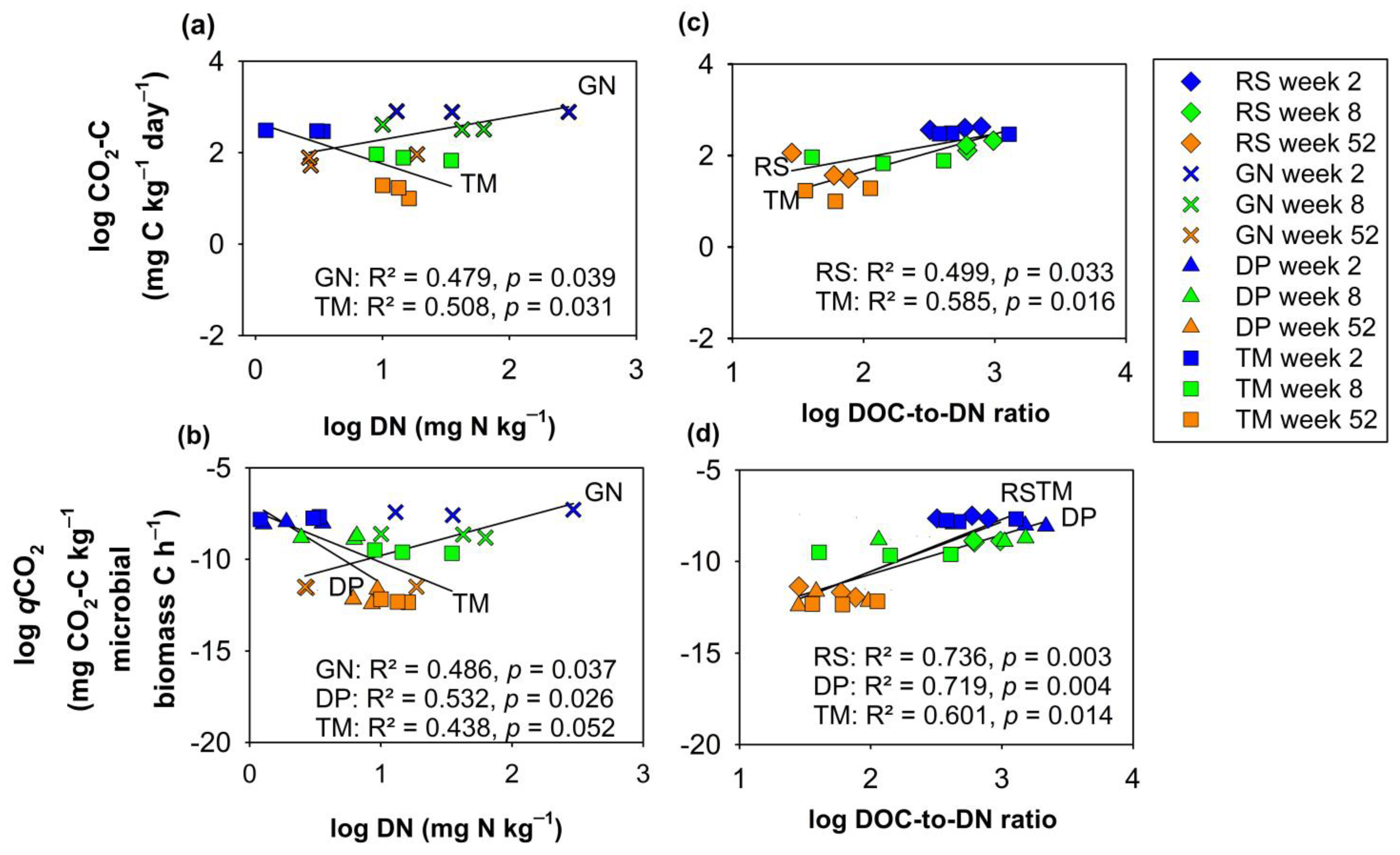
3.2. Relationships of DOM with Chemical Composition of Organic Residues and Soil Microbial Parameters at Various Decomposition Stages
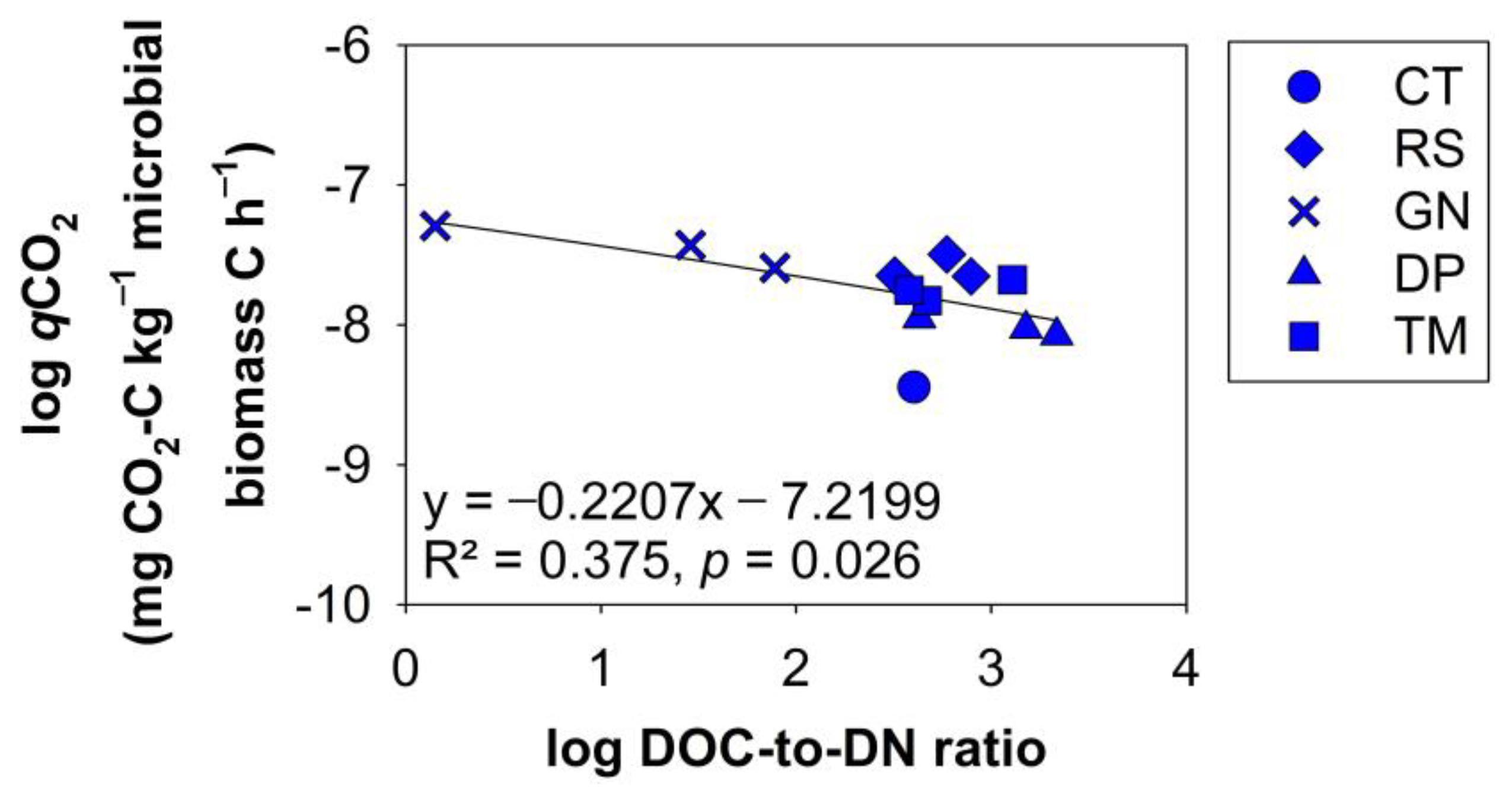
3.3. The Influence of DOM Produced by Each Chemical Quality Class (Residue) on Soil Microbial Processing
4. Discussion
4.1. Organic Residues Are the Main Source of DOC
4.2. Changes in DOC and DN Concentrations in Soils during Decomposition of Applied Contrasting Quality Class of Organic Residues Indicate the Biological Function and the Interaction of DOC and DN
4.3. Contrasting Chemical Quality Classes of Organic Residues Have Different Influences on Soil Micro-Bial Processing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krull, E.S.; Skjemstad, J.O.; Baldock, J.A. Functions of Soil Organic Matter and the Effect on Soil Properties; Cooperative Research Centre for Greenhouse Accounting, Glen Osmond SA: Canberra, Australia, 2004; p. 129. [Google Scholar]
- De Troyer, I.; Amery, F.; Van Moorleghem, C.; Smolders, E.; Merckx, R. Tracing the Source and Fate of Dissolved Organic Matter in Soil after Incorporation of a 13C Labelled Residue: A Batch Incubation Study. Soil Biol. Biochem. 2011, 43, 513–519. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of Soil Organic Matter via Biochemical and Physical Pathways of Litter Mass Loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Q.; Zhang, W.; Wang, X.; Mao, R.; Tigabu, M.; Ma, X. Linkage of Aggregate Formation, Aggregate-Associated C Distribution, and Microorganisms in Two Different-Textured Ultisols: A Short-Term Incubation Experiment. Geoderma 2021, 394, 114979. [Google Scholar] [CrossRef]
- Ghani, A.; Sarathchandra, U.; Ledgard, S.; Dexter, M.; Lindsey, S. Microbial Decomposition of Leached or Extracted Dissolved Organic Carbon and Nitrogen from Pasture Soils. Biol. Fertil. Soils. 2013, 49, 747–755. [Google Scholar] [CrossRef]
- Mariano, E.; Jones, D.L.; Hill, P.W.; Trivelin, P.C.O. Mineralisation and Sorption of Dissolved Organic Nitrogen Compounds in Litter and Soil from Sugarcane Fields. Soil Biol. Biochem. 2016, 103, 522–532. [Google Scholar] [CrossRef]
- Fang, Y.; Singh, B.P.; Collins, D.; Armstrong, R.; Van Zwieten, L.; Tavakkoli, E. Nutrient Stoichiometry and Labile Carbon Content of Organic Amendments Control Microbial Biomass and Carbon-Use Efficiency in a Poorly Structured Sodic-Subsoil. Biol. Fertil. Soils 2020, 56, 219–233. [Google Scholar] [CrossRef]
- Soong, J.L.; Parton, W.J.; Calderon, F.; Campbell, E.E.; Cotrufo, M.F. A New Conceptual Model on the Fate and Controls of Fresh and Pyrolized Plant Litter Decomposition. Biogeochemistry 2015, 124, 27–44. [Google Scholar] [CrossRef]
- Abdalla, K.; Sun, Y.; Zarebanadkouki, M.; Gaiser, T.; Seidel, S.; Pausch, J. Long-Term Continuous Farmyard Manure Application Increases Soil Carbon When Combined with Mineral Fertilizers Due to Lower Priming Effects. Geoderma 2022, 428, 116216. [Google Scholar] [CrossRef]
- Bucka, F.B.; Kölbl, A.; Uteau, D.; Peth, S.; Kögel-Knabner, I. Organic Matter Input Determines Structure Development and Aggregate Formation in Artificial Soils. Geoderma 2019, 354, 113881. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A. Stoichiometric Imbalances between Terrestrial Decomposer Communities and Their Resources: Mechanisms and Implications of Microbial Adaptations to Their Resources. Front. Microbiol. 2014, 5, 22. [Google Scholar] [CrossRef]
- Widdig, M.; Schleuss, P.M.; Biederman, L.A.; Borer, E.T.; Crawley, M.J.; Kirkman, K.P.; Seabloom, E.W.; Wragg, P.D.; Spohn, M. Microbial Carbon Use Efficiency in Grassland Soils Subjected to Nitrogen and Phosphorus Additions. Soil Biol. Biochem. 2020, 146, 107815. [Google Scholar] [CrossRef]
- Kallenbach, C.M.; Frey, S.D.; Grandy, A.S. Direct Evidence for Microbial-Derived Soil Organic Matter Formation and Its Ecophysiological Controls. Nat. Commun. 2016, 7, 13630. [Google Scholar] [CrossRef] [PubMed]
- Cotrufo, M.F.; Lavallee, J.M. Soil Organic Matter Formation, Persistence, and Functioning: A Synthesis of Current Understanding to Inform Its Conservation and Regeneration. In Advances in Agronomy, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2022; Volume 172. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; Blackwell Scientific Publications: Oxford, UK, 1979; p. 372. [Google Scholar]
- Palm, C.A.; Gachengo, C.N.; Delve, R.J.; Cadisch, G.; Giller, K.E. Organic Inputs for Soil Fertility Management in Tropical Agroecosystems: Application of an Organic Resource Database. Agric. Ecosyst. Environ. 2001, 83, 27–42. [Google Scholar] [CrossRef]
- Puttaso, A.; Vityakon, P.; Rasche, F.; Saenjan, P.; Treloges, V.; Cadisch, G. Does Organic Residue Quality Influence Carbon Retention in a Tropical Sandy Soil? Soil Sci. Soc. Am. J. 2013, 77, 1001–1011. [Google Scholar] [CrossRef]
- Kunlanit, B.; Vityakon, P.; Puttaso, A.; Cadisch, G.; Rasche, F. Mechanisms Controlling Soil Organic Carbon Composition Pertaining to Microbial Decomposition of Biochemically Contrasting Organic Residues: Evidence from MidDRIFTS Peak Area Analysis. Soil Biol. Biochem. 2014, 76, 100–108. [Google Scholar] [CrossRef]
- Puttaso, A.; Vityakon, P.; Saenjan, P.; Cadisch, G. Relationship between Residue Quality, Decomposition Patterns, and Soil Organic Matter Accumulation in a Tropical Sandy Soil after 13 Years. Nutr. Cycl. Agroecosyst. 2011, 89, 159–174. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) Framework Integrates Plant Litter Decomposition with Soil Organic Matter Stabilization: Do Labile Plant Inputs Form Stable Soil Organic Matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef]
- Xu, X.; Schimel, J.P.; Janssens, I.A.; Song, X.; Song, C.; Yu, G.; Sinsabaugh, R.L.; Tang, D.; Zhang, X.; Thornton, P.E. Global Pattern and Controls of Soil Microbial Metabolic Quotient. Ecol. Monogr. 2017, 87, 429–441. [Google Scholar] [CrossRef]
- Li, J.; Sang, C.; Yang, J.; Qu, L.; Xia, Z.; Sun, H.; Jiang, P.; Wang, X.; He, H.; Wang, C. Stoichiometric Imbalance and Microbial Community Regulate Microbial Elements Use Efficiencies under Nitrogen Addition. Soil Biol. Biochem. 2021, 156, 108207. [Google Scholar] [CrossRef]
- Kiikkilä, O.; Smolander, A.; Kitunen, V. Degradability, Molecular Weight and Adsorption Properties of Dissolved Organic Carbon and Nitrogen Leached from Different Types of Decomposing Litter. Plant Soil 2013, 373, 787–798. [Google Scholar] [CrossRef]
- Vityakon, P.; Meepech, S.; Cadisch, G.; Toomsan, B. Soil Organic Matter and Nitrogen Transformation Mediated by Plant Residues of Different Qualities in Sandy Acid Upland and Paddy Soils. NJAS-Wagening. J. Life Sci. 2000, 48, 75–90. [Google Scholar] [CrossRef]
- Anderson, J.P.E. Agronomy monograph number 9, part II. In Chemical and Biological Properties; American Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 1982; pp. 831–871. [Google Scholar]
- Amato, M.; Ladd, J.N. Assay for microbial biomass based on ninhydrin-reactive nitrogen in extracts of fumigated soils. Soil Biol. Biochem. 1988, 20, 107–114. [Google Scholar] [CrossRef]
- Anderson, T.H.; Domsch, K.H. Carbon link between microbial biomass and soil organic matter. In Perspectives in Microbial Ecology, Proceedings of the 4th International Symposium on Microbial Ecology, Ljubljana, Slovenia, 24–29 August 1986; Slovene Society for Microbiology: Ljubljana, Slovenia, 1986; pp. 467–471. [Google Scholar]
- Kunlanit, B.; Rasche, F.; Puttaso, A.; Cadisch, G.; Vityakon, P. Vertical Dynamics of Dissolved Organic Carbon in Relation to Organic Input Quality and Microaggregate Formation in a Coarse–Textured Ultisol. Eur. J. Soil Sci. 2020, 71, 459–471. [Google Scholar] [CrossRef]
- Cook, S.; Peacock, M.; Evans, C.D.; Page, S.E.; Whelan, M.; Gauci, V.; Khoon, K.L. Cold Storage as a Method for the Long-Term Preservation of Tropical Dissolved Organic Carbon (DOC). Mires Peat 2016, 18, 1–8. [Google Scholar] [CrossRef]
- Bowering, K.L.; Edwards, K.A.; Wiersma, Y.F.; Billings, S.A.; Warren, J.; Skinner, A.; Ziegler, S.E. Dissolved Organic Carbon Mobilization Across a Climate Transect of Mesic Boreal Forests Is Explained by Air Temperature and Snowpack Duration. Ecosystems 2023, 26, 55–71. [Google Scholar] [CrossRef]
- Fellman, J.B.; D’Amore, D.V.; Hood, E.; Boone, R.D. Fluorescence Characteristics and Biodegradability of Dissolved Organic Matter in Forest and Wetland Soils from Coastal Temperate Watersheds in Southeast Alaska. Biogeochemistry 2008, 88, 169–184. [Google Scholar] [CrossRef]
- Wallace, B.; Purcell, M.; Furlong, J. Total organic carbon analysis as a precursor to disinfection byproducts in potable water: Oxidation technique considerations. J. Environ. Monit. 2002, 4, 35–42. [Google Scholar] [CrossRef]
- Laurén, A.; Lappalainen, M.; Saari, P.; Kukkonen, J.V.K.; Koivusalo, H.; Piirainen, S.; Setälä, H.; Sarjala, T.; Bylund, D.; Heinonen, J.; et al. Nitrogen and Carbon Dynamics and the Role of Enchytraeid Worms in Decomposition of L, F and H Layers of Boreal Mor. Water. Air. Soil Pollut. 2012, 223, 3701–3719. [Google Scholar] [CrossRef]
- Marschner, B.; Kalbitz, K. Controls of Bioavailability and Biodegradability of Dissolved Organic Matter in Soils. Geoderma 2003, 113, 211–235. [Google Scholar] [CrossRef]
- Schleuss, P.M.; Widdig, M.; Biederman, L.A.; Borer, E.T.; Crawley, M.J.; Kirkman, K.P.; Seabloom, E.W.; Wragg, P.D.; Spohn, M. Microbial Substrate Stoichiometry Governs Nutrient Effects on Nitrogen Cycling in Grassland Soils. Soil Biol. Biochem. 2021, 155, 108168. [Google Scholar] [CrossRef]
- Kalbitz, K.; Schwesig, D.; Schmerwitz, J.; Kaiser, K.; Haumaier, L.; Glaser, B.; Ellerbrock, R.; Leinweber, P. Changes in Properties of Soil-Derived Dissolved Organic Matter Induced by Biodegradation. Soil Biol. Biochem. 2003, 35, 1129–1142. [Google Scholar] [CrossRef]
- Poosathit, R.; Vityakon, P.; Kunlanit, B.; Rasche, F. Molecular Structure of Dissolved Organic Carbon in a Sandy Soil Receiving Contrasting Quality Organic Residues. Geoderma 2023, 440, 116720. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Hämmerle, I.; Fuchslueger, L.; Hofhansl, F.; Knoltsch, A.; Schnecker, J.; Takriti, M.; Watzka, M.; Wild, B.; et al. Adjustment of Microbial Nitrogen Use Efficiency to Carbon:Nitrogen Imbalances Regulates Soil Nitrogen Cycling. Nat. Commun. 2014, 5, 3694. [Google Scholar] [CrossRef]
- Manzoni, S.; Taylor, P.; Richter, A.; Porporato, A.; Ågren, G.I. Environmental and Stoichiometric Controls on Microbial Carbon-Use Efficiency in Soils. New Phytol. 2012, 196, 79–91. [Google Scholar] [CrossRef]
- Manzoni, S.; Čapek, P.; Mooshammer, M.; Lindahl, B.D.; Richter, A.; Šantrůčková, H. Optimal Metabolic Regulation along Resource Stoichiometry Gradients. Ecol. Lett. 2017, 20, 1182–1191. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Manzoni, S.; Moorhead, D.L.; Richter, A. Carbon Use Efficiency of Microbial Communities: Stoichiometry, Methodology and Modelling. Ecol. Lett. 2013, 16, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Córdova, S.C.; Olk, D.C.; Dietzel, R.N.; Mueller, K.E.; Archontouilis, S.V.; Castellano, M.J. Plant Litter Quality Affects the Accumulation Rate, Composition, and Stability of Mineral-Associated Soil Organic Matter. Soil Biol. Biochem. 2018, 125, 115–124. [Google Scholar] [CrossRef]
- Rappe-George, M.O.; Choma, M.; Čapek, P.; Börjesson, G.; Kaštovská, E.; Šantrůčková, H.; Gärdenäs, A.I. Indications That Long-Term Nitrogen Loading Limits Carbon Resources for Soil Microbes. Soil Biol. Biochem. 2017, 115, 310–321. [Google Scholar] [CrossRef]
- Kamolmanit, B.; Vityakon, P.; Kaewpradit, W.; Cadisch, G.; Rasche, F. Soil Fungal Communities and Enzyme Activities in a Sandy, Highly Weathered Tropical Soil Treated with Biochemically Contrasting Organic Inputs. Biol. Fertil. Soils. 2013, 49, 905–917. [Google Scholar] [CrossRef]
- Widdig, M.; Heintz-Buschart, A.; Schleuss, P.M.; Guhr, A.; Borer, E.T.; Seabloom, E.W.; Spohn, M. Effects of Nitrogen and Phosphorus Addition on Microbial Community Composition and Element Cycling in a Grassland Soil. Soil Biol. Biochem. 2020, 151, 108041. [Google Scholar] [CrossRef]
- Pingthaisong, W.; Blagodatsky, S.; Vityakon, P.; Cadisch, G. Mixing Plant Residues of Different Quality Reduces Priming Effect and Contributes to Soil Carbon Retention. Soil Biol. Biochem. 2024, 188, 109242. [Google Scholar] [CrossRef]
- Blagodatskaya, E.; Kuzyakov, Y. Mechanisms of Real and Apparent Priming Effects and Their Dependence on Soil Microbial Biomass and Community Structure: Critical Review. Biol. Fertil. Soils 2008, 45, 115–131. [Google Scholar] [CrossRef]
- Zhang, Q.; Cheng, L.; Feng, J.; Mei, K.; Zeng, Q.; Zhu, B.; Chen, Y. Nitrogen Addition Stimulates Priming Effect in a Subtropical Forest Soil. Soil Biol. Biochem. 2021, 160, 108339. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treat- Ments 1 | Initial Organic Residue Quality 2 | Quality Class | ||||||
|---|---|---|---|---|---|---|---|---|
| Carbon (C) | Nitrogen (N) | Lignin (L) | Cellulose (CL) | C-to-N Ratio | L-to-N Ratio | LCI 3 | ||
| (g kg−1) | ||||||||
| RS | 367 | 5 | 29 | 507 | 78 | 6 | 0.05 | Low quality (low N but high CL) |
| GN | 388 | 23 | 68 | 178 | 17 | 3 | 0.28 | High quality (high N but low L) |
| DP | 453 | 6 | 175 | 306 | 80 | 31 | 0.37 | Low quality (low N but high L) |
| TM | 427 | 14 | 88 | 143 | 32 | 6 | 0.38 | Medium quality (medium N and L) |
| Treat- Ments 1 | Variables Pertaining to Soil Microbial Characteristics during Different Decomposition Stages (Weeks after Residue Application) 2 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Week 2 | Week 8 | Week 52 | ||||||||||||
| CO2-C | MBC | MBN | qCO2 | CO2-C | MBC | MBN | qCO2 | CO2-C | MBC | MBN | qCO2 | |||
| CT | 3 ± 0 | 52 ± 7 | 6 ± 1 | 2.82 × 10−4 | 4 ± 0 | 48 ± 9 | 6 ± 1 | 1.22 × 10−4 | 3 ± 0 | 88 ± 6 | 8 ± 1 | 0.10 × 10−4 | ||
| RS | 13 ± 0 | 168 ± 20 | 31 ± 1 | 5.02 × 10−4 | 9 ± 1 | 100 ± 11 | 26 ± 2 | 1.37 × 10−4 | 6 ± 1 | 142 ± 7 | 23 ± 1 | 0.09 × 10−4 | ||
| GN | 18 ± 0 | 182 ± 29 | 49 ± 4 | 5.92 × 10−4 | 13 ± 1 | 106 ± 9 | 32 ± 2 | 1.68 × 10−4 | 6 ± 0 | 140 ± 12 | 27 ± 2 | 0.10 × 10−4 | ||
| DP | 6 ± 0 | 123 ± 6 | 22 ± 2 | 3.29 × 10−4 | 9 ± 1 | 93 ± 3 | 22 ± 1 | 1.48 × 10−4 | 4 ± 1 | 156 ± 4 | 20 ± 2 | 0.06 × 10−4 | ||
| TM | 12 ± 0 | 178 ± 21 | 34 ± 2 | 4.30 × 10−4 | 8 ± 0 | 130 ± 3 | 20 ± 1 | 0.68 × 10−4 | 3 ± 0 | 142 ± 10 | 25 ± 0 | 0.05 × 10−4 | ||
| Parameters | DF | p-Values and Significance Levels 1 | ||
|---|---|---|---|---|
| DOC | DN | DOC-to-DN Ratio | ||
| Pr > F | Pr > F | Pr > F | ||
| Residue quality classes (Q) 2 | 4 | 0.0273 * | <0.0001 *** | <0.0001 *** |
| Decomposition time (T) 3 | 3 | <0.0001 *** | <0.0001 *** | <0.0001 *** |
| Q × T | 12 | 0.0948 ns | 0.0004 *** | 0.0007 *** |
| Soil Parameters | Time (Week) | Carbon (C) | Nitrogen (N) | Lignin (L) | Cellulose | C-to-N Ratio | L-to-N Ratio | Lignocellulose Index (LCI) |
|---|---|---|---|---|---|---|---|---|
| DOC | 2 | 0.452 | −0.214 | 0.397 | −0.002 | 0.267 | 0.462 | 0.252 |
| 8 | −0.271 | 0.097 | −0.198 | 0.050 | −0.140 | −0.168 | −0.081 | |
| 52 | 0.428 | 0.359 | 0.335 | −0.641 * | −0.301 | −0.018 | 0.530 | |
| DN | 2 | −0.252 | 0.728 ** | −0.021 | −0.416 | −0.751 ** | −0.563 | 0.236 |
| 8 | −0.202 | 0.762 ** | −0.096 | −0.610 * | −0.778 ** | −0.647 * | 0.266 | |
| 52 | 0.367 | 0.151 | 0.279 | −0.393 | −0.104 | 0.096 | 0.369 | |
| DOC-to-DN ratio | 2 | 0.523 | −0.655 * | 0.347 | 0.336 | 0.728 ** | 0.662 * | −0.079 |
| 8 | −0.129 | −0.586 * | −0.141 | 0.652 * | 0.563 | 0.356 | −0.427 | |
| 52 | −0.075 | 0.312 | −0.025 | −0.244 | −0.317 | −0.254 | 0.116 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poosathit, R.; Kunlanit, B.; Rasche, F.; Vityakon, P. Different Quality Classes of Decomposing Plant Residues Influence Dissolved Organic Matter Stoichiometry Which Results in Different Soil Microbial Processing. Soil Syst. 2024, 8, 28. https://doi.org/10.3390/soilsystems8010028
Poosathit R, Kunlanit B, Rasche F, Vityakon P. Different Quality Classes of Decomposing Plant Residues Influence Dissolved Organic Matter Stoichiometry Which Results in Different Soil Microbial Processing. Soil Systems. 2024; 8(1):28. https://doi.org/10.3390/soilsystems8010028
Chicago/Turabian StylePoosathit, Ratanaporn, Benjapon Kunlanit, Frank Rasche, and Patma Vityakon. 2024. "Different Quality Classes of Decomposing Plant Residues Influence Dissolved Organic Matter Stoichiometry Which Results in Different Soil Microbial Processing" Soil Systems 8, no. 1: 28. https://doi.org/10.3390/soilsystems8010028
APA StylePoosathit, R., Kunlanit, B., Rasche, F., & Vityakon, P. (2024). Different Quality Classes of Decomposing Plant Residues Influence Dissolved Organic Matter Stoichiometry Which Results in Different Soil Microbial Processing. Soil Systems, 8(1), 28. https://doi.org/10.3390/soilsystems8010028