


Insects Associated with Ancient Human Remains: How Archaeoentomology Can Provide Additional Information in Archaeological Studies




Abstract
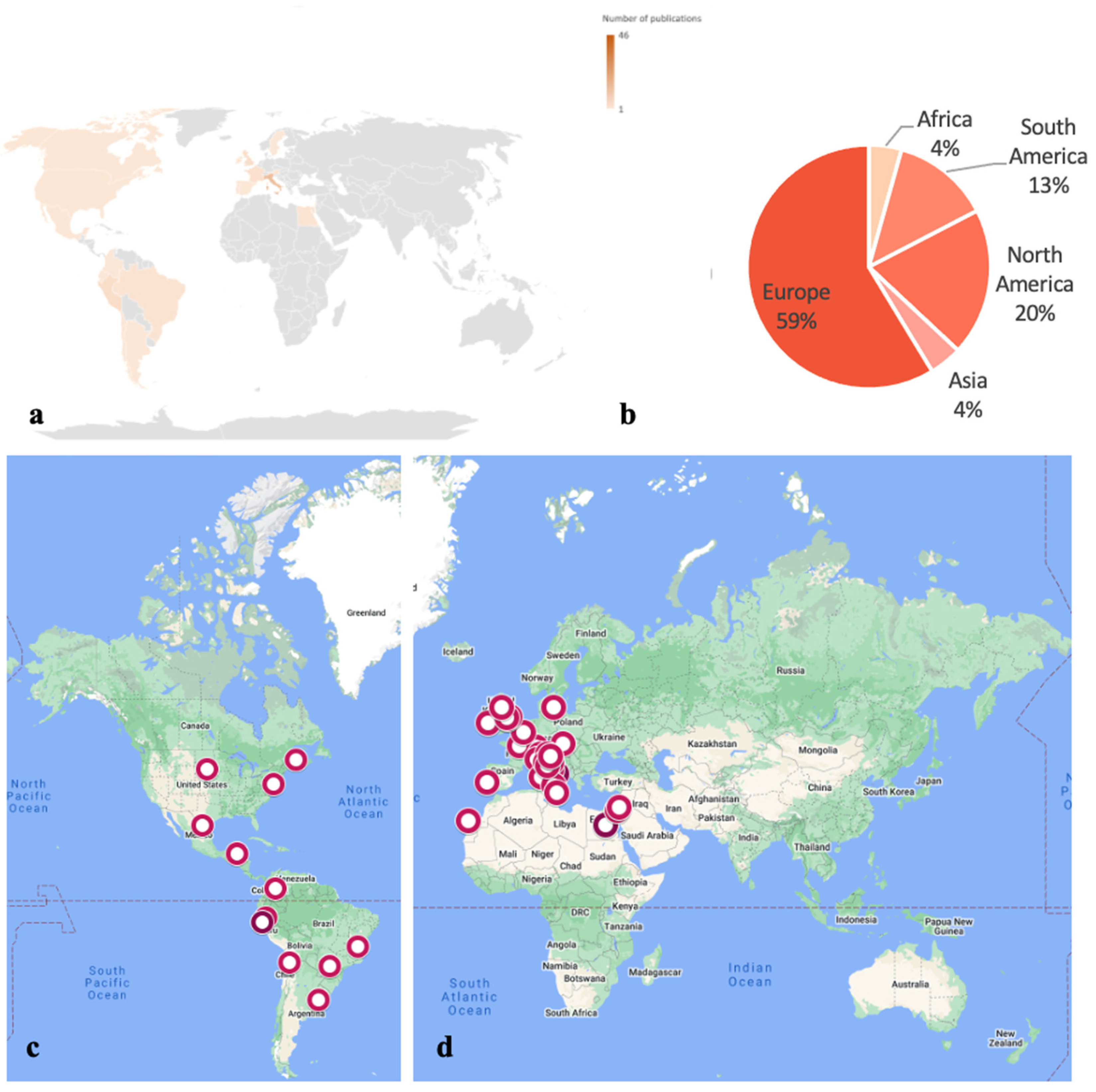
:1. Introduction
2. Funerary Archaeoentomology
3. Insects of Archaeoentomological Interest
4. Insect Species Identification with a Special Focus on Archaeological Settings
4.1. Morphological Identification of Insects
4.2. Cleaning Procedures for Insect Samples Collected from Archaeological Settings
4.3. Molecular Identification of Insects
4.4. Molecular Identification of Insects in Archaeological Settings
4.5. Hydrocarbon Analyses for the Identification of Insects
5. Information Provided by the Entomofauna Collected from an Archaeological Setting
5.1. Information regarding the Timeline of Death and Decomposition
5.2. Information regarding Funerary Practices
5.3. Information regarding Causes of Death
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Continent | Country | Region/ State/ Province/ County | City/Town | Type of Study | Number of Bodies | Insect and Other Arthropod Species Identified | Citation | ||
|---|---|---|---|---|---|---|---|---|---|
| Diptera | Coleoptera | Other | |||||||
| Africa | Egypt | n.a. | n.a. | Museal mummified remains | 2 | NR | Attagenus sp. Latreille (Dermestidae)—FA | Genus? sp.? (Lepidoptera: Tineidae)—C Tinea pellionella Linnaeus (Lepidoptera: Tineidae)—C | [57] |
| n.a. | n.a. | Museal mummified remains | 1 | Genus? sp.? (Calliphoridae)—P Piophila casei (Linnaeus) (Piophilidae)—P | Genus? sp.? (Dermestes)—A Genus? sp.? (Staphylinidae)—L | NR | [56] | ||
| America (South) | Brazil | Minas Gerais | Itacambria | Mummified remains | 1 | Megaselia scalaris (Loew) (Phoridae)—Pu | NR | NR | [55] |
| Paranáb | Altonia | Funerary urn | 1 | Euxesta sp. (Ulidiidae)—A | Bembidion sp. (Carabidae)—A Corticaria sp. (Latridiidae)—A Cossonus sp. (Curculionidae)—A Genus? sp.? (Curculionidae)—FA Genus? sp.? (Latridiidae)—FA Genus? sp.? (Scarabidae)—FA Genus? sp.? (Scolytinae)—FA Lagria villosa (Fabricius) (Tenebrionidae)—A Nilio sp. (Tenebrionidae)—FA Osoriellus sp. (Staphylinidae)—A Xyleborus affinis (Eichhoff) (Curculionidae)—A | Campontus sp. (Hymenoptera: Formicidae)—A Genus? sp.? (Blattaria)—A Genus? sp.? (Hemiptera: Cicadellidae)—A Genus? sp.? (Lepidoptera: Tineidae)—L Hypoponera sp. (Hymenoptera: Formicidae)—A | [20] | ||
| Chile | Atacama | Calama | Funerary bundle | 7 | Genus? sp.? (Sarcophagidae)—Pu | Anthrenus sp. (Dermestidae)—L Dermestes peruvianus Laporte (Dermestidae)—USL | Genus? sp.? (Lepidoptera: Tineidae)—C | [80] | |
| Colombia | Andean region | Bogotá | La Candelaria cave | 2 | Genus? sp.? (Calliphoridae)—A Genus? sp.? (Muscidae)—Pu Genus? sp.? (Sarcophagidae)—Pu | Dermestes maculatus De Geer (Dermestidae)—A Genus? sp.? (Dermestidae)—L Genus? sp.? (Histeridae)—A, FA Genus? sp.? (Trogidae)—FA Lasioderma serricorne (Fabricius) (Ptinidae) | NR | [117] | |
| Ecuador | Napo | n.a. | Funerary urn | 1 | NR | NR | Genus? sp.? (Blattodea)—BD possibly caused by termites | [22] | |
| Argentina | Beunos Aires | La Plata | Buried remains | 1 | Fannia canicularis (Lineaus) (Fanniidae)—Pu Genus? sp.? (Sarcophagidae)—P Hydrotaea aenescens (Wiedemann) (Muscidae)—Pu, FA Megaselia scalaris (Loew) (Phoridae)—Pu Muscina stabulans Fallén (Muscidae)—Pu | Atheta sp. (Staphylinidae)—A Carpophilus sp. (Nitidulidae)—FA Genus? sp.? (Dermestidae)—L Genus? sp.? (Tenebrionidae)—L | Genus? sp.? (Acari: Gamasidae)—A Genus? sp.? (Blattodea)—O Genus? sp.? (Dermaptera: Anisolabididae)—FA Genus? sp.? (Juliformia)—A Genus? sp.? (Polydesmida)—A Tineola bisselliella (Hummel) (Lepidoptera: Tineidae)—Pu | [125] | |
| America (North) | Belize | n.a. | n.a. | Maya mortuary cave | 25 | NR | Genus? sp.? (Dermestidae)—PC | Genus? sp.? (Blattodea)—BD possibly caused by termites | [51] |
| Canada | New Brunswick | n.a. | Buried remains | NR | Cynomyopsis cadaverina (Robineau-Desvoidy) (Calliphoridae)—Pu Genus? sp.? (Heleomyzidae)—Pu Hydrotaea sp. (Muscidae)—Pu Muscina assimilis (Fallén) (Muscidae)—Pu Phormia regina (Meigen) (Calliphoridae)—Pu Protophormia terraenovae (Robineau-Desvoidy) (Calliphoridae)—Pu | NR | NR | [140] | |
| Mexico | Coahuila | n.a. | Funerary cave | 1 | Fannia sp. (Fanniidae)—Pu Genus? sp.? (Sarcophagidae)—Pu Synthesiomyia nudiseta Van der Wulp (Muscidae)—Pu | Dermestes carnivorous Fabricius (Dermestidae)—A Genus? sp.? (Staphylinidae)—USL Saprinus (s. str.) alienus J.L. Le Conte (Histeridae)—FA Niptus sp. (Anobiidae)—A Omorgus sp. (Trogidae)—FA Xerosaprinus (s. str.) coerulescens (J.L. Le Conte) (Histeridae)—FA Xerosaprinus (s. str.) vitiosus (J.L. Le Conte) (Histeridae) –A | Acromyrmex veriscolor (Pergande) (Hymenoptera: Formicidae)—A Genus? sp.? (Lepidoptera: Family?)—USL | [133] | |
| Peru | Amazonas | Chachapoyas | Mummy bundle | 1 | Genus? sp.? (Calliphoridae)—Pu | NR | Genus? sp.? (Hymenoptera)—A Genus? sp.? (Lepidoptera: Tineidae)—Pu, FA | [111] | |
| La Libertad | Trujillo | Entombed remains | 1 | Cochliomyia macellaria (Fabricius) (Calliphoridae)—Pu Compsomyops verena (Walker) (Calliphoridae)—Pu Hydrotaea aenescens (Wiedemann) (Muscidae)—Pu Sarcophaga sp. (Sarcophagidae)—Pu Synthesiomyia nudiseta Van der Wulp (Muscidae)—Pu | Omorgus suberosus (Fabricius) (Trogidae)—FA | Genus? sp.? (Hymenoptera: Pteromalidae?)—PP | [99] | ||
| Trujillo | Moche pyramids | 1 | Cochliomyia macellaria (Fabricius) (Calliphoridae)—Pu Compsomyiops verena (Walker) (Calliphoridae)—Pu Genus? sp.? (Sarcophagidae)—Pu Hydrotaea aenescens (Wiedemann) (Muscidae)—Pu Synthesiomyia nudiseta Van der Wulp (Muscidae)—Pu | Omorgus suberosus (Fabricius) (Trogidae)—FA | Muscidifurax or Sphalangia sp.? (Hymenoptera: Pteromalidae?)—PP | [68] | |||
| 1 | NR | NR | Genus? sp.? (Blattodea)—BD possibly caused by termites | [48] | |||||
| Lima | Lurin | Pachacamac site | n.a. | NR | NR | Genus? sp.? (Pseudoscorpiones: Cheiridiidae)—A | [31] | ||
| Untied States of America | South Dakota | n.a. | Buried remains | n.a. | Genus? sp.? (Calliphoridae)—P Genus? sp.? (Sarcophagidae)—P | NR | NR | [106] | |
| Asia | Israel | n.a. | n.a. | Munhata site | n.a. | NR | Dermestes sp. (Dermestidae)—PC | NR | [49] |
| Palestine | Jericho | Jericho site | n.a. | NR | Dermestes sp. (Dermestidae)—PC | NR | [49] | ||
| Europe | Austria | Niederösterreich | Lanzenkirhcen | Buried remains | 7 | Calliphora sp. (Calliphoridae)—USL Lucillia sp. (Calliphoridae)—USL Ophyra capensis (Wiedemann) (Muscidae)—P Ophyra sp. (Muscidae)—Pu Protophormia terranovae Robinaeu-Desvoidy (Calliphoridae)—USL | NR | NR | [126,127] |
| Vienna | Vienna | Skeletal remains | 3 | Calliphora sp. (Calliphoridae)—Pu Conicera tibialis Schmitz (Phoridae)—Pu Lucilia sp. (Calliphoridae)—Pu Sarcophaga sp. (Sarcophagidae)—Pu | NR | NR | [104] | ||
| France | Alsace | Carspach | Skeletal remains | n.a. | Ophyra capensis (Wiedeman) (Muscidae)—Pu | NR | NR | [107] | |
| Centre-Val de Loire | Clery-Saint-André | Sarcophagus | 1 | NR | Attagenus sp. (Dermestidae)—USL Bruchus sp. (Bruchidae)—USL Leistus spinibarbis (Dejean) (Carabidae: Nebriinae)—A Necrobia salina Fairmaire and Laboulbene (Carabidae: Nebriinae)—A Othius laeviusculus Stephens (Staphylinidae: Staphylininae)—USL Oxyomus sylvestris Scopoli (Aphodiidae)—USL Platystethus arenarius (Geoffroy) (Staphylinidae: Oxytelinae)—USL Ptinus sp. (Anobiidae: Ptininae)—USL Tasgius ater (Gravenhorst) (Staphylinidae: Staphylininae)—USL | NR | [21] | ||
| Hauts-de-France | Lille | Coffins | 22 | Calliphora vicina Robineau-Desvoidy (Calliphoridae)—Pu, A Conicera tibialis Schmitz (Phoridae)—L, P Fannia scalaris (Fabricius) (Fanniidae)—L Fannia manicata (Meigen) (Fanniidae)—L, A Hydrotaea capensis (Weidemann) (Muscidae)—LII, LIII P, A Hydrotaea sp. (Muscidae)—P Leptocera caenosa (Robdani) (Sphaeroceridae)—L, P, A Megaselia sp. (Phoridae)—L, P Megaselia rufipes (Meigen) (Phoridae)—P Phoridae sp. (Phoridae)—P Triphelba hyalinata (Meigen) (Phoridae)—A | Omalium rivulare (Paykull) (Staphylinidae)—A Philonthus sp. (Staphylinidae)—A Staphylinidae sp. (Staphylinidae)—L | NR | [103] | ||
| Ireland | Kildimo | Limerick | Cemeterial buried remains | 6 | Calliphora vicina Robineau-Desvoidy (Calliphoridae)—P, FA Calliphora vomitoria (Linneaus) (Calliphoridae)—P, FA Genus? sp.? (Phoridae)—P | NR | NR | [121] | |
| Italy | Campania | Naples | Church | 4 | Coincera cfr tibialis Schmitz (Phoridae)—Pu Fannia sp. (Fanniidae)—Pu Hydrotaea capensis (Wiedemann) (Muscidae)—Pu, FA | Genus? sp.? (Dermestidae)—L Ptinus sp. (Ptinidae)—FA | Genus? sp.? (Tineidae)—L, C | [137] | |
| Monastery | 1 | Hermetia illucens (Linnaeus) (Stratiomyidae)—L | NR | NR | [109] | ||||
| Lombardia | Azzio | Crypt | n.a.. | Genus? sp.? (Phoridae)—Pu Hydrotaea capensis (Wiedemann) (Muscidae)—Pu | Cryptophagus montanus C.Bristout de Barneville (Cryptophagidae)—L Genus? sp.?—A Quedius sp. (Staphylinidae) –A | Genus? sp.? (Acarina)—A | [131] | ||
| Lazio regione | Rome | Catacomb | 2 | Calliphora sp. (cf. vicina) (Calliphoridae)—Pu Hydrotaea capensis (Wiedemann) (Muscidae)—Pu | Ablattaria laevigata (Fabricius) (Silphidae)—FA Alphitiobius diaperinus Panzer (Tenebrionidae)—USL Carabus (Archicarabus) alysidotus Illiger (Carabidae)—FA Dermestes sp. (Dermestidae)—L, FA Geotrupes spiniger (Marsham) (Geotrupidae)—FA Jekelius intermedius (Costa) (Geotrupidae)—FA Necrobia rufipes (De Geer) (Cleridae)—FA | NR | [23] | ||
| Marche | Urbino | Church | 1 | Hydrotaea leucostoma (Wiedemann) (Muscidae)—P, FA | Genus? sp.? (Dermestidae)—L, FA | Cydia splendana (Hübner) (Lepidoptera: Torticidae)—Ch | [116] | ||
| Pieddmont | Fossano | Monastery | 1 | Hydrotaea capensis (Wiedemann) (Musdicae)—Pu, A | Anthrenus verbascii Linneaus (Dermestidae)—A Genus? sp.? (Carabidae)—A Langelandia anophtalma Aubé (Zopheridae)—A Mycetaea subterranean (Fabricius)—A | Genus? sp.? (Aranea)—FA Messor sp. (Hymenoptera: Formicidae)—A | [124] | ||
| Sardinia | n.a. | Cathedral | 2 | Calliphora vicina Robineau-Desvoidy (Calliphoridae)—Pu Genus? sp.?—FA Hydrotaea capensis (Wiedemann) (Muscidae)—Pu Phormia regina Meigen (Calliphoridae)—Pu Sarcophaga sp. (Sarcophagidae)—Pu | Genus? sp.? (Tenebrionidae)—FA Saprinus semistriatus (Scriba) (Histeridae)—FA | Gen? sp.? (Tineidae)—C | [110] | ||
| Sicily | Palermo | Catacomb | 667 | Coincera tibialis Schmitz (Phoridae)—FA Fannia scalaris (Fabricius) (Fanniidae) -USL Hydrotaea ignava (Harris) (Muscidae)—USL Leptocera sp. (Sphaeroceridae)—USL | Gibbium psylloides Czenpiński (Ptinidae)—A Necrobia rufipes (De Geer) (Cleridae)—USL Oryzaephilus surinamensis (Linneaus) (Silvanidae)—A | Genus? sp.? (Arachnida: Pseudoscorpions)—FA Genus? sp.? (Hymenoptera: Braconidae)—USL Tinella Pellionella Linneaus (Tineidae)—USL | [140] | ||
| Trentino-Alto Adige | Laghi | Buried remains | 1 | Fannia canicularis (Linneaus) (Fanniidae)—Pu Phormia regina Meigen (Calliphoridae)—Pu Protophormia terraenovae (Robineau-Desvoidy) (Calliphoridae)—Pu | NR | NR | [15] | ||
| Tuscany | Florence | Embalming jars | 10 jars containing soft tissue | Conicera tibialis Schmitz (Phoridae)—Pu Hydrotaea capensis (Wiedemann) (Muscidae)—Pu | Ptinus dibius Sturm (Ptinidae)—FA Ptinus subpilosus Sturm (Ptinidae)—FA | NR | [119] | ||
| Lucca | Entombed remains | 1 | Conicera sp. (Phoridae)—Pu Hydrotaea capensis (Wiedemann) (Muscidae)—Pu Muscina sp. (Muscidae)—Pu | Anobium punctatum (Ptinidae)—FA Gnathoncus sp. (Histeridae)—FA Necrobia sp. (Cleridae)—FA Sitophilus granarius (Curculionidae)—FA Trox scaber (Trogidae)—FA | Genus? sp.? (Hymenoptera)—FA Genus? sp.? (Ichneumonidae: Julida)—FA Genus? sp.? (Ichneumonidae: Scorpiones)—FA Genus? sp.? (Lepidoptera: Tineidae)—C Genus? sp.? (Lepidoptera: Pyralidae)—C | [129] | |||
| Monticiano | Entombed remains | 1 | Hydrotaea capensis (Wiedemann) (Muscidae)—Pu | NR | Genus? sp.? (Lepidoptera: Tineidae)—FA | [81] | |||
| Veneto | Venice | Monastery | 1 | Chrysomya albiceps (Wiedemann) (Calliphoridae)—P, Pu | Anthrenus sp. (Dermestidae)—L Anthrenus (Nathrenus) verbasci (Linneaus) (Dermestidae)—L, A Attagenus (s. str.) unicolor (Brahm) (Dermestidae)—L Necrobia rufipes (De Geer) (Cleridae)—A | Genus? sp.? (Arachnida: Pseudoscorpions)—USL | [120] | ||
| n.a. | Mass grave | 7 | Protophormia terraenovae Robinaeu-Desvoidy (Calliphoridae)—Pu | NR | NR | [107] | |||
| Malta | Northern region of Malta | St Pauls Bay | Xemxija tombs | 15,000 fragmented bones | NR | Genus? sp.? (Dermestidae)—PC | NR | [50] | |
| Portugal | Lisboa region | Lisbon | Crypt | 1 | Ophyra capensis (Wiedemann) (Muscidae)—Pu, FA | NR | Genus? sp. (Diplopoda) | [39,40] | |
| Spain | Macaronesia | Las Palmas de Gran Canaria | Mummified remains | 3 | Chrysomya albiceps (Wiedemann) (Calliphoridae)—Pu Genus? sp.? (Fanniidae)—USL | Dermestes maculatus De Geer (Dermestidae)—A, FA Mezium americanum Laporte de Castelnau (Anobiidae)—A Necrobia rufipes (De Geer) (Cleridae)—A Stegobium paniceum (Linneaus) (Anobiidae)—A | Genus? sp.? (Tineidae)—USL Pheidole sp. (Hymenoptera: Formicidae)—FA Nosopsyllus fasciatus (Siphonaptera: Ceratophyllidae)—FA | [136] | |
| Sweden | Scania | Lund | Coffin | 1 | Chloromyia formosa (Scopoli) (Stratiomyidae)—USL Genus? sp.?—USL Genus? sp.? (Anisopodidae)—USL Genus? sp.? (Calliphoridae)—USL Genus? sp.? (Chaoboridae)—USL Genus? sp.? (Heleomyzidae)—USL Genus? sp.? (Rhagionidae)—USL Mochlonyx sp. (Chaoboridae)—USL | Amara ovata (Fabricius) (Carabidae)—USL Anobium punctatum De Geer (Anobiidae)—USL Atomaria munda Erichson (Cryptophagidae)—USL Atomaria nigripennis Kugelann (Cryptophagidae)—A Attagenus pellio (Linneaus) (Dermestidae)—USL Corticaria fulva (Comolli) (Latridiidae)—USL Cryptophagus dentatus (Herbst) (Cryptophagidae)—USL Cryptophagus cf. distinguendus Strum (Cryptophagidae)—USL Cryptophagus saginatus Strum (Cryptophagidae)—USL Cryptophagus cellaris (Scopoli) (Cryptophagidae)—USL Dermestes lardarius Linneaus (Dermestidae)—USL Epauloecus unicolor (Piller & Mitterpacher) (Ptinidae)—A Ernobius mollis (Linneaus) (Anobiidae)—USL Latridius minutus (Linneaus) (Latridiidae)—USL Meligethes cf. aeneus (Fabricius) (Nitidulidae)—USL Mycetaea subterranea (Fabricius) (Endomychidae)—USL Ocys quinquestriatus (Gyllenhal) (Carabidae)—USL Orthoperus sp. (Corylophidae)—USL Phyllodrepa puberula (Bernhauer) (Staphylinidae)—USL Polydrusus cf. flavipes (De Geer) (Curculionidae)—USL Protapion sp. (Apionidae) –USL Ptinus fur (Linneaus) (Ptinidae)—USL Stegobium paniceum (Linneaus) (Anobiidae)—USL Typhaea sp. (Mycetophagidae)—USL | Aphidoidea sp. (Hemiptera: Aphididae)—USL Cheyletus sp. (Trombidformes: Cheyletidae)—USL Cimex lectularius Linneaus (Hemiptera: Cimicidae)—USL Coleophora sp. (Lepidoptera: Coleophoridae)—USL Dahlicini sp. (Lepidoptera: Psychidae)—USL Eulaelaps stabularis (C.L. Koch) (Mesostigmata: Haemogamasidae)—A Eulohmannia ribagai (Berlese) (Sarcoptiformes: Eulohmannidae)—USL Eupteryx aurata (Linneaus) (Hemiptera: Cicadellidae)—USL Forficula auricularia Linneaus (Dermaptera: Forficulidae)—USL Genus? sp.? (Araneae)—USL Genus? sp.? (Hymenoptera)—USL Genus? sp.? (Hymenoptera: Pteromalidae)—USL Hemannia sp. (Oribatida: Hermanniidae)—USL Hypoaspis sp. (Mesostigmata: Laelapidae)—USL Lasius niger (Linneaus) (Hymenoptera: Formicidae)—USL Lygaeidae sp. (Hemiptera: Lygaeidae)—USL Macrosiphoniella cf. abrotani (Walker) (Hemiptera: Aphididae)—FA Parasitus sp. (Mesostigmata: Parasitidae)—USL Phorodon humuli (Schrank) (Hemiptera: Aphididae)—USL Picrostigeus/Batakomacrus sp. (Hymenoptera: Ichneumonidae)—USL Scotophaeus cf. scutulatus (L. Koch) (Araneae: Gnaphosidae)—USL Steatoda cf. bipunctata (Linneaus) (Araneae: Theridiidae)—USL Tegenaria domestica (Clerck) (Araneae: Agelenidae)—USL Tinea pellionella (Linneaus) (Lepidoptera: Tineidae)—USL | [135] | |
| UK | England | Manchester | Museal mummified remains | 4 | Genus? sp.? (Chrysomya)—L, Pu Genus? sp.? (Piophilidae)—Pu, A Musca domestica (Linnaeus) (Muscidae)—Pu | Genus? sp.? (Carabidae)—FA Gibbium psylloides Czenpiński (Ptinidae)—FA Mesotsenopa sp. (Tenebrionidae)—FA Necrobia rufipes (Fabricius) (Cleridae)—A | Blatta orientalis Linnaeus (Dictyoptera: Blattodea)—O | [118] | |
| North Yorkshire | n.a. | Cathedral | 1 | Genus? sp.? (Phoridae)—Pu Genus? sp.? (Sphaeroceridae)—Pu | Aleocharinae sp. (Staphylinidae)—USL Cryptophagus sp. (Cryptophagidae)—FA Mycetaea subterranea (Marsham) (Endomycidae)—FA Phyllodrpa floralis (Paykull) (Staphylinidae)—USL Quedius mesomelinus (Marsham) (Staphylinidae)—FA Rhizophagus parallelocollis Gyllenhal (Monotomidae)—FA | NR | [16] | ||
| Scotland | Black Burn | Monastery | 3 | Fannia sp. (Fannidae)—Pu Genus? sp.? (Heleomyzidae)—Pu Genus? sp.? (Leptoceridae)—FA Genus? sp.? (Muscidae)—Pu Genus? sp.? (Phoridae)—Pu Genus? sp.? (Pscyhodidae)—Pu Terrilmosina racovitzai (Bezzi) (Sphareoceridae)—P Trichocera sp. (Trichoceridae)—L | Acrolocha sulcula (Stephens) (Staphylinidae)—USL Aleochara sp. (Staphylinidae)—USL Genus? sp.? (Staphylinidae: Aleocharinae)—USL Aphodius rufipes (Linneaus) (Scarabaeidae)—USL Atomaria sp. (Cryptophagidae)—USL Catops sp. (Leiodidae)—USL Corticaria sp. (Cryptophagidae)—USL Cryptophagus scutellatus (Newman) (Cryptophagidae)—USL Cryptophagus sp. (Cryptophagidae)—USL Lathridius minutus (Linnaeus) (Lathridiidae)—USL Ootypus globosus (Waltl) (Cryptophagidae)—USL Philonthus sp. (Staphylinidae)—USL Ptenidium sp. (Ptiliidae) –USL Quedius mesomelinus (Marsham) (Staphylinidae)—USL Rhizophagus parallelocollis (Gyllenhal) (Rhizophagidae)—USL Tipnus unicolor (Piller and Mitterpacher) (Ptinidae)—USL Trichocellus sp. (Carabidae)—FA Xylodromus concinnus (Marsham) (Staphylinidae)—USL | NR | [19] | ||
| Glasgow | Cathedral | 1 | NR | Rhizophagus parallelocollis Gyllenhal (Monotomidae)—USL | NR | [18] | |||
References
- Whitworth, T. Fossilized human remains from northern Kenya. Nature 1960, 185, 947–948. [Google Scholar] [CrossRef]
- Dent, B.B.; Forbes, S.L.; Stuart, B.H. Review of human decomposition processes in soil. Environmental 2004, 45, 576–585. [Google Scholar] [CrossRef]
- Zumpt, F. Myiasis in Man and Animals in the Old World: A Textbook for Physicians, Veterinarians and Zoologists; Butterworth: London, UK, 1965. [Google Scholar]
- Voss, S.C.; Spafford, H.; Dadour, I.R. Annual and seasonal patterns of insect succession on decomposing remains at two locations in Western Australia. Forensic Sci. Int. 2009, 193, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Vass, A.A. Beyond the grave-understanding human decomposition. Microbiol. Today 2001, 28, 190–193. [Google Scholar]
- Goff, M.L. Early postmortem changes and stages of decomposition. In Current Concepts in Forensic Entomology, 1st ed.; Amendt, J., Campobasso, C., Goff, M., Grassberger, M., Eds.; Springer Dordrecht: Dordrecht, The Netherlands, 2009; pp. 1–24. [Google Scholar]
- Huchet, J.B. L’Archéoentomologie funéraire: Une approche originale dans l’interprétation des sépultures. Bull. Mem. Soc. Anthropol. Paris 1996, 8, 299–311. [Google Scholar] [CrossRef]
- Mégnin, P. La Faune des Cadavres: Application de l’entomologie à la Médecine Légale; G. Masson: Paris, France, 1894. [Google Scholar]
- Wells, J.D. A forensic entomological analysis can yield an estimate of postmortem interval, and not just a minimum postmortem interval: An explanation and illustration using a case. J. Forensic Sci. 2019, 64, 634–637. [Google Scholar] [CrossRef]
- Matuszewski, S. Post-mortem interval estimation based on insect evidence: Current challenges. Insects 2021, 12, 314. [Google Scholar] [CrossRef]
- Amendt, J.; Campobasso, C.P.; Gaudry, E.; Reiter, C.; LeBlanc, H.N.; Hall, M. Best practice in forensic entomology—Standards and guidelines. Int. J. Leg. Med 2007, 121, 90–104. [Google Scholar] [CrossRef]
- Giordani, G. Non-Invasive Approaches to Morphological and Molecular Identification of Insects from Museum, Archaeo-funerary and Forensic Contexts. Ph.D. Thesis, University of Huddersfield, Huddersfield, UK, 2019. [Google Scholar]
- Huchet, J.B. Insect remains and their traces: Relevant fossil witnesses in the reconstruction of past funerary practices. Anthropologie 2014, 52, 329–346. [Google Scholar]
- Buckland, P.I.; Buckland, P.C.; Olsson, F. Paleoentomology: Insects and other arthropods in environmental archaeology. In Encyclopedia of Global Archaeology, 2nd ed.; Smith, C., Ed.; Springer: New York, NY, USA, 2020; pp. 8291–8312. [Google Scholar]
- Vanin, S.; Turchetto, M.; Galassi, A.; Cattaneo, C. Forensic entomology and the archaeology of war. J. Confl. Archaeol. 2009, 5, 127–139. [Google Scholar] [CrossRef]
- Panagiotakopulu, E.; Buckland, P.C. Forensic archaeoentomology—An insect fauna from a burial in York Minster. Forensic Sci. Int. 2012, 221, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Buckland, P.C.; Cooper, G.R.; Sadler, J.P. A Bibliography of Quaternary Entomology. Trans. Ent. Soc. 2004, 116, 329–346. [Google Scholar]
- Buckland, P.C. Insect fauna from within the cranium of skeleton 570. In Excavations at Glasgow Cathedral; Routledge: Abingdon, UK, 1988; Volume 1997, p. 157. [Google Scholar]
- McCormick, F.; Buckland, P.; Carter, S.; Cerón-Carrasco, R.; Fawcett, R.; Ford, B.; Flavell, D.A.; Hamilton-Dyer, S.; Lorimer, D.H.; Mills, C. Excavations at Pluscarden Priory, Moray; Society of Antiquaries of Scotland: Edinburgh, Scotland, 1995. [Google Scholar]
- Macari, B.P. Arqueoentomologia: Um estudo de Caso Tupiguarani, Altônia, Paraná, Brasil; Federal University of Paraná: Paraná, Brasil, 2013. [Google Scholar]
- Huchet, J.B. Les sépultures prestigieuses de l’église Notre-Dame de Cléry-saint-André (Loiret): Etude pluridisciplinaire du caveau de Louis XI. In Les Sépultures Prestigieuses de l’église Notre-Dame de Cléry-Saint-André (Loiret); Georges-Zimmermann, P., Ed.; L’Harmattan: Paris, France, 2015; pp. 172–183. [Google Scholar]
- Jastremski, N.A.; Sánchez-Polo, A. Human Skeletal Remains Recovered from a Napo Funerary Urn in the Ecuadorian Amazon: A Taphonomic and Mortuary Assessment. Bioarchaeol. Int 2021, 5, 143. [Google Scholar] [CrossRef]
- Huchet, J.B.; Castex, D. The walking dead-life after death—Archaeoentomological evidence in a Roman catacomb: (Saints Marcellinus and Peter, central area, 1st–3rd century AD). In The Routledge Handbook of Archaeothanatology, 1st ed.; Knüsel, C.J., Schotsman, E.M.J., Eds.; Routledge: Oxforshire, UK, 2022; pp. 481–498. [Google Scholar]
- Hope, F.W. Observations on some mummified beetles taken from the inside of a mummified ibis. Trans. Ent. Soc. 1842, 3, 191–193. [Google Scholar]
- Huchet, J.B.; Callou, C.; Lichtenberg, R.; Dunand, F. The dog mummy, the ticks and the louse fly: Archaeological report of severe ectoparasitosis in Ancient Egypt. Int. J. Paleopathol. 2013, 3, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Giordani, G.; Erauw, C.; Eeckhout, P.A.; Owens, L.S.; Vanin, S. Patterns of camelid sacrifice at the site of Pachacamac, Peruvian Central Coast, during the Late Intermediate Period (AD1000–1470): Perspectives from funerary archaeoentomology. J. Archaeol. Sci. 2020, 114, 105065. [Google Scholar] [CrossRef]
- Han, H.; Liu, Z.; Meng, F.; Jiang, Y.; Cai, J. Identification of olfactory genes of a forensically important blow fly, Aldrichina grahami (Diptera: Calliphoridae). PeerJ 2020, 8, e9581. [Google Scholar] [CrossRef]
- Barton, P.S.; Archer, M.S.; Quaggiotto, M.M.; Wallman, J.F. Invertebrate succession in natural terrestrial environments. In Forensic Entomology: The Utility of Arthropods in Legal Investiations, 3rd ed.; Byrd, J.H., Tomberlin, J.K., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 141–153. [Google Scholar]
- Smith, K.G. A Manual of Forensic Entomology, 1st ed.; Cornell University Press: New York, NY, USA, 1986. [Google Scholar]
- Magni, P.A.; Voss, S.C.; Testi, R.; Borrini, M.; Dadour, I.R. A biological and procedural review of forensically significant Dermestes species (Coleoptera: Dermestidae). J. Med. Entomol. 2015, 52, 755–769. [Google Scholar] [CrossRef]
- Morrow, J.J.; Taylor, L.; Peck, L.; Elowsky, C.; Owens, L.S.; Eeckhout, P.; Reinhard, K.J. Pseudoscorpions of the family Cheiridiidae (Arachnida: Pseudoscorpiones) recovered from burial sediments at Pachacamac (500–1500 CE), Perú. J. Arachnol. 2017, 45, 370–375. [Google Scholar] [CrossRef] [Green Version]
- Kadej, M.; Szleszkowski, Ł.; Thannhäuser, A.; Jurek, T. A mummified human corpse and associated insects of forensic importance in indoor conditions. Int. J. Leg. Med. 2020, 134, 1963–1971. [Google Scholar] [CrossRef]
- Gaudry, E. The insects colonisation of buried remains. In Current Concepts in Forensic Entomology, 1st ed.; Springer Dordrecht: Dordrecht, The Netherlands, 2009; pp. 273–311. [Google Scholar]
- Pastula, E.C.; Merritt, R.W. Insect arrival pattern and succession on buried carrion in Michigan. J. Med. Entomol. 2013, 50, 432–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, L.; Moreau, G.; Czuprynski, S.; Bernhardt, V.; Amendt, J. An empirical comparison of decomposition and fly colonisation of concealed carcasses in the Old and New World. Int. J. Leg. Med. 2019, 133, 1593–1602. [Google Scholar] [CrossRef] [PubMed]
- Magni, P.A.; Petersen, C.; Georgy, J.; Dadour, I.R. The effect of suitcase concealment on insect colonization: A pilot study in Western Australia. GJFSM 2019, 1, 513. [Google Scholar] [CrossRef]
- Schoenly, K. A statistical analysis of successional patterns in carrion-arthropod assemblages: Implications for forensic entomology and determination of the postmortem interval. J. Forensic Sci. 1992, 37, 1489–1513. [Google Scholar] [CrossRef]
- Ibrahim, A.A.; Galal, F.H.; Seufi, A.M.; Elhefnawy, A.A. Insect succession associated with corpse’s decomposition of the guinea pig Cavia porcellus in Benha city, Egypt. Egypt. Acad. J. Biol. 2013, 5, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Couri, M.S.; de Souza, S.M.F.M.; Cunha, A.M.; Pinheiro, J.; Cunha, E. Diptera Brachycera found inside the esophagus of a mummified adult male from the early XIX century, Lisbon, Portugal. Mem. Inst. Oswaldo Cruz 2008, 103, 211–213. [Google Scholar] [CrossRef] [Green Version]
- Couri, M.S.; Cunha, A.M.; de Souza, S.M.F.M.; Laeta, M. Ophyra capensis (Wiedemann)(Diptera, Muscidae) found inside the esophagus of a mummy in Lisbon (Portugal). Pap. Avulsos Zool. 2009, 49, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Magni, P.A.; Guareschi, E.E. After the flood: A multidisciplinary investigation of human remains found in a floodplain and first record of raft spiders colonizing a corpse. JCHS 2021, 6, 68–75. [Google Scholar]
- Guareschi, E.; Dadour, I.R.; Magni, P.A. A taphonomic examination of inhumed and entombed remains in Parma cemeteries, Italy. GJFSM 2019, 1, 1–8. [Google Scholar]
- Magni, P.A.; Borrini, M.; Dadour, I.R. Human remains found in two wells: A forensic entomology perspective. Forensic Sci. Med. Pathol. 2013, 9, 413–417. [Google Scholar] [CrossRef]
- Byrd, J.H.; Castner, J.L. Forensic Entomology: The Utility of Arthropods in Legal Investigations, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Gemmellaro, M.D.; Bucolo, C.; Musumeci, E.; Hamilton, G.C.; Weidner, L.M. First observations of initial blow fly (Diptera: Calliphoridae) activity on lava fields and in subterranean environments in Sicily in cool temperatures. J. Med. Entomol. 2018, 55, 1622–1626. [Google Scholar] [CrossRef] [PubMed]
- Goff, M.L. Estimation of postmortem interval using arthropod development and successional patterns. Forensic Sci. Rev. 1993, 5, 81. [Google Scholar] [PubMed]
- Kenward, H.; Large, F. Recording the preservational condition of archaeological insect fossils. Environ. Archaeol 1998, 2, 49–60. [Google Scholar] [CrossRef]
- Huchet, J.B.; Deverly, D.; Gutierrez, B.; Chauchat, C. Taphonomic evidence of a human skeleton gnawed by termites in a Moche-civilisation grave at Huaca de la Luna, Peru. Int. J. Osteoarchaeol. 2011, 21, 92–102. [Google Scholar] [CrossRef]
- Huchet, J.B.; Le Mort, F.; Rabinovich, R.; Blau, S.; Coqueugniot, H.; Arensburg, B. Identification of dermestid pupal chambers on Southern Levant human bones: Inference for reconstruction of Middle Bronze Age mortuary practices. J. Archaeol. Sci. 2013, 40, 3793–3803. [Google Scholar] [CrossRef]
- Thompson, J.E.; Martin-Vega, D.; Buck, L.T.; Power, R.K.; Stoddart, S.; Malone, C. Identification of dermestid beetle modification on Neolithic Maltese human bone: Implications for funerary practices at the Xemxija tombs. J. Archaeol. Sci. Rep. 2018, 22, 123–131. [Google Scholar] [CrossRef]
- Wrobel, G.D.; Biggs, J. Osteophageous insect damage on human bone from Je’reftheel, a Maya mortuary cave site in west-central Belize. Int. J. Osetoarchaeol. 2018, 28, 745–756. [Google Scholar] [CrossRef]
- Powers, N.; Sibun, L. Standards and Guidance for Forensic Archaeologists; Institute for Archaeology: Earley, Reading, UK, 2016. [Google Scholar]
- Magni, P.A.; Guercini, S.; Leighton, A.; Dadour, I. Forensic entomologists: An evaluation of their status. J. Insect Sci. 2013, 13, 78. [Google Scholar] [CrossRef] [Green Version]
- Sanford, M.R.; Byrd, J.H.; Tomberlin, J.K.; Wallace, J.R. Entomological evidence collection methods: American board of forensic entomology approved protocols. In Forensic Entomology: The Utility of Arthropods in Legal Investigations, 3rd ed.; Byrd, J.H., Tomberlin, J.K., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 63–85. [Google Scholar]
- Braga, M.V.; Mendonça, P.M.; Barbosa, R.R.; Blomquist, G.J.; Novo, S.P.C.; Dutra, J.M.F.; de Souza, S.M.; Queiroz, M.M.C. Identification of Megaselia scalaris (Loew, 1866)(Diptera: Phoridae) in mummified human body from Itacambira (MG), Brazil, using scanning electron microscopy and cuticular hydrocarbons. J. Nat. Hist. 2016, 50, 1381–1388. [Google Scholar] [CrossRef]
- Cockburn, A.; Barraco, R.A.; Reyman, T.A.; Peck, W.H. Autopsy of an Egyptian mummy: A mummy can be a scientific treasure chest; to unlock its secrets, a multidisciplinary approach is needed. Science 1975, 187, 1155–1160. [Google Scholar] [CrossRef]
- Oras, E.; Anderson, J.; Tõrv, M.; Vahur, S.; Rammo, R.; Remmer, S.; Mölder, M.; Malve, M.; Saag, L.; Saage, R. Multidisciplinary investigation of two Egyptian child mummies curated at the University of Tartu Art Museum, Estonia (Late/Graeco-Roman Periods). PLoS ONE 2020, 15, e0227446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raguso, R.A. Don’t forget the flies: Dipteran diversity and its consequences for floral ecology and evolution. Appl. Entomol. Zool. 2020, 55, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Lucidcentral: Identification and Diagnostic Tools. Available online: https://www.lucidcentral.org (accessed on 26 August 2022).
- Szpila, K.; Richet, R.; Pape, T. Third instar larvae of flesh flies (Diptera: Sarcophagidae) of forensic importance—Critical review of characters and key for European species. Parasitol. Res. 2015, 114, 2279–2289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukontason, K.; Sukontason, K.L.; Piangjai, S.; Boonchu, N.; Kurahashi, H.; Hope, M.; Olson, J.K. Identification of forensically important fly eggs using a potassium permanganate staining technique. Micron 2004, 35, 391–395. [Google Scholar] [CrossRef]
- Byrd, J.H. Laboratory rearing of forensic insects. In Forensic Entomology: The Utility of Arthropods in Legal Investigations, 2nd ed.; Byrd, J.H., Castner, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2009; pp. 177–200. [Google Scholar]
- Weeks, J.D.; O’Neill, M.A.; Gaston, K.J.; Gauld, I.D. Automating insect identification: Exploring the limitations of a prototype system. J. Appl. Entomol. 1999, 123, 1–8. [Google Scholar] [CrossRef]
- Yang, H.P.; Ma, C.S.; Wen, H.; Zhan, Q.B.; Wang, X.L. A tool for developing an automatic insect identification system based on wing outlines. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef]
- Nelson, H.G. A method of cleaning insects for study. Coleoptera. Bull. 1949, 3, 89–92. [Google Scholar]
- Giordani, G.; Grzywacz, A.; Vanin, S. Characterization and identification of puparia of Hydrotaea Robineau-Desvoidy, 1830 (Diptera: Muscidae) from forensic and archaeological contexts. J. Med. Entomol. 2019, 56, 45–54. [Google Scholar] [CrossRef]
- Pradelli, J.; Tuccia, F.; Giordani, G.; Vanin, S. Puparia cleaning techniques for forensic and archaeo-funerary studies. Insects 2021, 12, 104. [Google Scholar] [CrossRef]
- Huchet, J.B.; Greenberg, B. Flies, Mochicas and burial practices: A case study from Huaca de la Luna, Peru. J. Archaeol. Sci. 2010, 37, 2846–2856. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information: Taxonomy. Available online: https://www.ncbi.nlm.nih.gov/taxonomy (accessed on 26 August 2022).
- Merheb, M.; Matar, R.; Hodeify, R.; Siddiqui, S.S.; Vazhappilly, C.G.; Marton, J.; Azharuddin, S.; Al Zouabi, H. Mitochondrial DNA, a powerful tool to decipher ancient human civilization from domestication to music, and to uncover historical murder cases. Cells 2019, 8, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandrini, F.; Mazzanti, M.; Onofri, V.; Turchi, C.; Tagliabracci, A. MtDNA analysis for genetic identification of forensically important insects. Forensic Sci. Int. Genet. 2008, 1, 584–585. [Google Scholar] [CrossRef]
- Thomsen, P.F.; Elias, S.; Gilbert, M.T.; Haile, J.; Munch, K.; Kuzmina, S.; Froese, D.G.; Sher, A.; Holdaway, R.N.; Willerslev, E. Non-destructive sampling of ancient insect DNA. PLoS ONE 2009, 4, e5048. [Google Scholar] [CrossRef] [PubMed]
- Mazzanti, M.; Alessandrini, F.; Tagliabracci, A.; Wells, J.D.; Campobasso, C.P. DNA degradation and genetic analysis of empty puparia: Genetic identification limits in forensic entomology. Forensic Sci. Int. 2010, 195, 99–102. [Google Scholar] [CrossRef]
- Tuccia, F.; Giordani, G.; Vanin, S. A combined protocol for identification of maggots of forensic interest. Sci. Justice 2016, 56, 264–268. [Google Scholar] [CrossRef]
- Iancu, L.; Junkins, E.N.; Necula-Petrareanu, G.; Purcarea, C. Characterizing forensically important insect and microbial community colonization patterns in buried remains. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Knapp, M.; Hofreiter, M. Next generation sequencing of ancient DNA: Requirements, strategies and perspectives. Genes 2010, 1, 227–243. [Google Scholar] [CrossRef]
- Parrott, J.J. A molecular Study of Contemporary and Museum Calliphoridae of Forensic Importance. Doctoral Dissertation, University of Portsmouth, Portsmouth, UK, 2013. [Google Scholar]
- Snounou, G.; Singh, B. Nested PCR analysis of Plasmodium parasites. In Malaria Methods and Protocols, 1st ed.; Doolan, D.L., Ed.; Humana Press: Totowa, NJ, USA, 2002; pp. 189–203. [Google Scholar]
- Behjati, S.; Tarpey, P.S. What is next generation sequencing? Arch. Dis. Child. 2013, 98, 236–238. [Google Scholar] [CrossRef] [Green Version]
- Squella, D.J. Insectos hallados en fardos funerarios provenientes del cementerio arqueológico de Topater (Región de Atacama, Chile). Acta Entomol. Chil. 2007, 31, 31–34. [Google Scholar]
- Morrow, J.J.; Baldwin, D.A.; Higley, L.; Piombino-Mascali, D.; Reinhard, K.J. Curatorial implications of Ophyra capensis (order Diptera, family Muscidae) puparia recovered from the body of the Blessed Antonio Patrizi, Monticiano, Italy (Middle Ages). J. Forensic Leg. Med. 2015, 36, 81–83. [Google Scholar] [CrossRef] [Green Version]
- Vanin, S.; Azzoni, M.; Giordani, G.; Belcastro, M.G. Bias and potential misinterpretations in the analysis of insects collected from human remains of archaeological interest. Archaeol. Anthropol. Sci. 2021, 13, 1–6. [Google Scholar] [CrossRef]
- Lockey, K.H. Insect hydrocarbon classes: Implications for chemotaxonomy. Insect Biochem. 1991, 21, 91–97. [Google Scholar] [CrossRef]
- Singer, T.L. Roles of hydrocarbons in the recognition systems of insects. Am. Zool. 1998, 38, 394–405. [Google Scholar] [CrossRef]
- Moore, H. Cuticular hydrocarbon analysis in forensic entomology: A Review. AEFS 2017, 1, 127–138. [Google Scholar] [CrossRef]
- Byrne, A.L.; Camann, M.A.; Cyr, T.L.; Catts, E.P.; Espelie, K.E. Forensic implications of biochemical differences among geographic populations of the black blow fly, Phormia regina (Meigen). J. Forensic Sci. 1995, 40, 372–377. [Google Scholar] [CrossRef]
- Bernier, U.R.; Carlson, D.A.; Geden, C.J. Gas chromatography/mass spectrometry analysis of the cuticular hydrocarbons from parasitic wasps of the genus Muscidifurax. J. Am. Soc. Mass Spectrom 1998, 9, 320–332. [Google Scholar] [CrossRef] [Green Version]
- Anyanwu, G.I.; Molyneux, D.H.; Phillips, A. Variation in cuticular hydrocarbons among strains of the Anopheles gambiae sensu stricto by analysis of cuticular hydrocarbons using gas liquid chromatography of larvae. Mem. Inst. Oswaldo Cruz 2000, 95, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Kather, R.; Martin, S.J. Cuticular hydrocarbon profiles as a taxonomic tool: Advantages, limitations and technical aspects. Physiol. Entomol. 2012, 37, 25–32. [Google Scholar] [CrossRef]
- Braga, M.V.; Pinto, Z.T.; de Carvalho Queiroz, M.M.; Matsumoto, N.; Blomquist, G.J. Cuticular hydrocarbons as a tool for the identification of insect species: Puparial cases from Sarcophagidae. Acta Trop. 2013, 128, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Kula, C.; Moore, H.E. The Temperature Effect on the Cuticular Chemical Profile of Lucilia Sericata Blowfly Larvae. Ph.D. Thesis, Turkish Ministry of National Education, Ankara, Turkey, 2020. [Google Scholar]
- Moore, H.; Lutz, L.; Bernhardt, V.; Drijfhout, F.P.; Cody, R.B.; Amendt, J. Cuticular hydrocarbons for the identification and geographic assignment of empty puparia of forensically important flies. Int. J. Leg. Med. 2022, 136, 1791–1800. [Google Scholar] [CrossRef]
- Moore, H.E.; Adam, C.D.; Drijfhout, F.P. Identifying 1st instar larvae for three forensically important blowfly species using “fingerprint” cuticular hydrocarbon analysis. Forensic Sci. Int. 2014, 240, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Moore, H.E.; Pechal, J.L.; Benbow, M.E.; Drijfhout, F.P. The potential use of cuticular hydrocarbons and multivariate analysis to age empty puparial cases of Calliphora vicina and Lucilia sericata. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Moore, H.E.; Hall, M.; Drijfhout, F.P.; Cody, R.B.; Whitmore, D. Cuticular hydrocarbons for identifying Sarcophagidae (Diptera). Sci. Rep. 2021, 11, 1–11. [Google Scholar]
- Martin, S.; Drijfhout, F. A review of ant cuticular hydrocarbons. J. Chem. Ecol. 2009, 35, 1151–1161. [Google Scholar] [CrossRef]
- Wagner, D.; Tissot, M.; Gordon, D. Task-related environment alters the cuticular hydrocarbon composition of harvester ants. J. Chem. Ecol. 2001, 27, 1805–1819. [Google Scholar] [CrossRef]
- Martin, S.J.; Helanterae, H.; Drijfhout, F.P. Evolution of species-specific cuticular hydrocarbon patterns in Formica ants. Biol. J. Linn. Soc. 2008, 95, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Huchet, J.B. Flies, the Dead and Offerings: Archaeoentomology of Mochica graves from the Pyramid of the Moon, Peru. Rech. Amerindien. Que. 2017, 47, 23–36. [Google Scholar]
- Vanin, S.; Huchet, J.B. Forensic entomology and funerary archaeoentomology. In Taphonomy of Human Remains: Forensic Analysis of the Dead and the Depositional Environment, 1st ed.; Schotsmans, E.M.J., Márquez-Grant, N., Forbes, S.L., Eds.; Wiley: Oxford, UK, 2017; pp. 167–186. [Google Scholar]
- Price, P.W.; Denno, R.F.; Eubanks, M.D.; Finke, D.L.; Kaplan, I. Insect Ecology: Behavior, Populations and Communities; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Anderson, G.S. Insect succession on carrion and its relationship to determining time of death. In Forensic Entomology: The Utility of Arthropods in Legal Investigations, 1st ed.; Byrd, J.H., Castner, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 143–175. [Google Scholar]
- Bourel, B.; Tournel, G.; Hédouin, V.; Gosset, D. Entomofauna of buried bodies in northern France. Int. J. Leg. Med. 2004, 118, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Waitzbauer, W. Dipterenreste aus Awarengräbern im Wiener Ortsgebiet. Verh. Zool.-Bot. Ges. Osterreich 1984, 122, 35–42. [Google Scholar]
- Le Cabec, A.; Colard, T.; Charabidze, D.; Chaussain, C.; Di Carlo, G.; Gaudzinski-Windheuser, S.; Hublin, J.J.; Melis, R.T.; Pioli, L.; Ramirez-Rozzi, F. Insights into the palaeobiology of an early Homo infant: Multidisciplinary investigation of the GAR IVE hemi-mandible, Melka Kunture, Ethiopia. Sci. Rep. 2021, 11, 1–14. [Google Scholar]
- Gilbert, B.M.; Bass, W.M. Seasonal dating of burials from the presence of fly pupae. Am. Antiq. 1967, 32, 534–535. [Google Scholar] [CrossRef]
- Gaudio, D.; Betto, A.; Vanin, S.; De Guio, A.; Galassi, A.; Cattaneo, C. Excavation and study of skeletal remains from a World War I mass grave. Int. J. Osteoarchaeol. 2015, 25, 585–592. [Google Scholar] [CrossRef]
- Huchet, J.B. L’archéo-entomologie: Les insectes nécrophages associés aux soldats de Carspach. A l’Est du Nouveau 2013, 109–110. [Google Scholar]
- Benelli, G.; Canale, A.; Raspi, A.; Fornaciari, G. The death scenario of an Italian Renaissance princess can shed light on a zoological dilemma: Did the black soldier fly reach Europe with Columbus? J. Archaeol. Sci. 2014, 49, 203–205. [Google Scholar] [CrossRef]
- Giordani, G.; Tuccia, F.; Floris, I.; Vanin, S. First record of Phormia regina (Meigen, 1826) (Diptera: Calliphoridae) from mummies at the Sant’Antonio Abate Cathedral of Castelsardo, Sardinia, Italy. PeerJ 2018, 6, e4176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nystrom, K.C.; Goff, A.; Goff, M.L. Mortuary behaviour reconstruction through palaeoentomology: A case study from Chachapoya, Perú. Int. J. Osteoarchaeol. 2005, 15, 175–185. [Google Scholar] [CrossRef]
- Pinto, S.L.; Giordani, G.; Tuccia, F.; Ventura, F.; Vanin, S. First records of Synthesiomyia nudiseta (Diptera: Muscidae) from forensic cases in Italy. Forensic Sci. Int. 2017, 276, 1–7. [Google Scholar] [CrossRef]
- Velásquez, Y.; Ivorra, T.; Grzywacz, A.; Martínez-Sánchez, A.; Magaña, C.; García-Rojo, A.; Rojo, S. Larval morphology, development and forensic importance of Synthesiomyia nudiseta (Diptera: Muscidae) in Europe: A rare species or just overlooked? Bull. Entomol. Res 2013, 103, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Campobasso, C.P.; Di Vella, G.; Introna, F. Factors affecting decomposition and Diptera colonization. Forensic Sci. Int. 2001, 120, 18–27. [Google Scholar] [CrossRef]
- Magni, P.A.; Pazzi, M.; Vincenti, M.; Alladio, E.; Brandimarte, M.; Dadour, I.R. Development and validation of a GC–MS method for nicotine detection in Calliphora vomitoria (L.)(Diptera: Calliphoridae). Forensic Sci. Int. 2016, 261, 53–60. [Google Scholar] [CrossRef]
- Masetti, M.; Menconi, M.; Fornaciari, G. Insect remains associated with the mummy of Cardinal Giulio della Rovere, Archbishop of Ravenna (1533–1578). In Proceedings of the VI World Congress on Mummy Studies, Teguise, Lanzarote, 20–24 February 2008; pp. 379–385. [Google Scholar]
- Vergara-Pineda, S.; Rojas-Chávez, J.M.; Mansilla, J.; Campos-de-la-Rosa, T. Fragmentos de insectos asociados a momias prehispánicas. Entomol. Mex 2009, 8, 798–801. [Google Scholar]
- David, A.R. The insects associated with the Manchester mummies. In The Manchester Museum Mummy Project; David, A.R. Manchester Museum: Manchester, UK, 1979; pp. 113–118. [Google Scholar]
- Morrow, J.J.; Myhra, A.; Piombino-Mascali, D.; Lippi, D.; Roe, A.; Higley, L.; Reinhard, K.J. Archaeoentomological and archaeoacarological investigations of embalming jar contents from the San Lorenzo Basilica in Florence, Italy. J. Archaeol. Sci. Rep. 2016, 10, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Huchet, J.B. Archaeoentomological study of the insects remains found within the mummy of Namenkhet Amon, San Lazzaro Armenian Monastery,(Venice/Italy). Adv. Egyptol. 2010, 1, 59–80. [Google Scholar]
- Lynch, L.G.; Reilly, E. Early medieval human burials and insect remains from Kildimo, Co. Limerick. J. Ir. Archaeol. 2011, 20, 65–76. [Google Scholar]
- Miles, D. Socio-economic aspects of secondary burial. Oceania 1965, 35, 161–174. [Google Scholar] [CrossRef]
- Guareschi, E.E.; Magni, P.A. Preliminary Taphonomical Comparison of the Decomposition Process in Simple Burials, Traditional Tombs and Aerated Tombs in an Urban Cemetery in Northern Italy. Forensic Sci. 2022, 2, 505–515. [Google Scholar] [CrossRef]
- Vanin, S.; Boano, R.; Giordani, G.; Carta, G.; Fulcheri, E. Description of the entomofauna associated with the remains of the Cistercian nun Angela Veronica Bava (1591–1637). Med. Hist. 2022, 6, e2022006. [Google Scholar]
- Mariani, R.; García-Mancuso, R.; Varela, G.L.; Inda, A.M. Entomofauna of a buried body: Study of the exhumation of a human cadaver in Buenos Aires, Argentina. Forensic Sci. Int. 2014, 237, 19–26. [Google Scholar] [CrossRef]
- Scharrer-Liška, G.; Grassberger, M. Archäoentomologische untersuchungen von grab 34 awarischen Gräberfeldes von Frohsdorf, Niederösterreich. Archaol. Korresp. 2005, 35, 531–544. [Google Scholar]
- Scharrer-Liška, G.; Grassberger, M. Aussagemöglichkeiten der Archäoentomologie anhand des awarischen Gräberfeldes von Frohsdorf, Niederösterreich. In Proceedings of the Erster Österreichischer Archäometriekongress, Salzburg, Austria, 15–17 May 2009. [Google Scholar]
- Fornaciari, A.; Giuffra, V.; Pezzini, F. Secondary burial and mummification practices in the Kingdom of the two Sicilies. Mortality 2010, 15, 223–249. [Google Scholar] [CrossRef]
- Loni, A.; Vanin, S.; Fornaciari, A.; Tomei, P.E.; Giuffra, V.; Benelli, G. Back to the Middle Ages: Entomological and Botanical Elements Reveal New Aspects of the Burial of Saint Davino of Armenia. Insects 2022, 13, 1113. [Google Scholar] [CrossRef]
- Pardo, I. L’elaborazione del lutto in un quartiere tradizionale di Napoli. (L’élaboration du deuil dans un quartier traditionnel de Naples). Ras. Ital. Soc. 1982, 23, 535–569. [Google Scholar]
- Pradelli, J.; Rossetti, C.; Tuccia, F.; Giordani, G.; Licata, M.; Birkhoff, J.M.; Verzeletti, A.; Vanin, S. Environmental necrophagous fauna selection in a funerary hypogeal context: The putridarium of the Franciscan monastery of Azzio (northern Italy). J. Archaeol. Sci. Rep. 2019, 24, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Biswadeep, P.; Patowary, A. Embalming: The Art of Preserving the Dead. IJHRMLP 2016, 2, 10. [Google Scholar]
- Barnes, K.M.; Whiffin, A.L.; Bulling, M.T. A preliminary study on the antibacterial activity and insect repellent properties of embalming fluids from the 18th Dynasty (1550–1292 BCE) in ancient Egypt. J. Archaeol. Sci. Rep. 2019, 25, 600–609. [Google Scholar] [CrossRef]
- Huchet, J.B.; Pereira, G.; Gomy, Y.; Philips, T.K.; Alatorre-Bracamontes, C.E.; Vásquez-Bolaños, M.; Mansilla, J. Archaeoentomological study of a pre-Columbian funerary bundle (mortuary cave of Candelaria, Coahuila, Mexico). Ann. Soc. Entomol. Fr. 2013, 49, 277–290. [Google Scholar] [CrossRef]
- Fägerström, C.; Buckland, P.I.; Lemdahl, G.; Karsten, P.; Lagerås, P.; Manhag, A. Insects and other invertebrate remains from the coffin of a 17th century bishop in Lund Minster, S Sweden. J. Archaeol. Sci. Rep. 2020, 31, 102299. [Google Scholar] [CrossRef]
- López-Dos Santos, N.; Martínez, C.P.; Darias, T.D.; Barroso, V.A.; Vázquez, J.V. Archaeoentomology applied to the Gran Canaria mummies: First results. In Proceedings of the Extraordinary World Congress on Mummy Studies, Tenerife, Spain, 5–9 September 2018. [Google Scholar]
- Loni, A.; Fornaciari, A.; Canale, A.; Giuffra, V.; Vanin, S.; Benelli, G. Insights on Funeral Practices and Insects Associated With the Tombs of King Ferrante II d’Aragona and Other Renaissance Nobles. J. Med. Entomol. 2019, 56, 1582–1589. [Google Scholar] [CrossRef]
- Giuffra, V.; Fornaciari, A.; Minozzi, S.; Vitiello, A.; Fornaciari, G. Autoptic practices in 16th–18th century Florence: Skeletal evidences from the Medici family. Int. J. Paleopathol. 2016, 15, 21–30. [Google Scholar] [CrossRef]
- Baumjohann, K.; Benecke, M. Insect Traces and the Mummies of Palermo—A Status Report. Entomol. Heute 2019, 31, 73–93. [Google Scholar]
- Teskey, H.J.; Turnbull, C. Diptera puparia from pre-historic graves. Can. Entomol. 1979, 111, 527–528. [Google Scholar] [CrossRef]
- Chophi, R.; Sharma, S.; Sharma, S.; Singh, R. Forensic entomotoxicology: Current concepts, trends and challenges. J. Forensic Leg. Med. 2019, 67, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.L.; Lord, W.D.; Goff, M.L.; Donnelly, B.; McDonough, E.T.; Alexis, J.C. Isolation of amitriptyline and nortriptyline from fly puparia (Phoridae) and beetle exuviae (Dermestidae) associated with mummified human remains. J. Forensic Sci. 1994, 39, 1305–1313. [Google Scholar] [CrossRef]
- Gagliano-Candela, R.; Aventaggiato, L. The detection of toxic substances in entomological specimens. Int. J. Leg. Med. 2001, 114, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Goff, M.L.; Brown, W.A.; Hewadikaram, K.A.; Omori, A.I. Effect of heroin in decomposing tissues on the development rate of Boettcherisca peregrina (Diptera, Sarcophagidae) and implications of this effect on estimation of postmortem intervals using arthropod development patterns. J. Forensic Sci. 1991, 36, 537–542. [Google Scholar] [CrossRef]
- Hédoui, V.; Bourel, B.; Martin-Bouyer, L.; Bécart, A.; Tournel, G.; Deveaux, M.; Gosset, D. Morphine perfused rabbits: A tool for experiments in forensic entomotoxicology. J. Forensic Sci. 1999, 44, 347–350. [Google Scholar] [CrossRef]
- Magni, P.A.; Pazzi, M.; Droghi, J.; Vincenti, M.; Dadour, I.R. Development and validation of an HPLC-MS/MS method for the detection of ketamine in Calliphora vomitoria (L.)(Diptera: Calliphoridae). J. Forensic Leg. Med. 2018, 58, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Nolte, K.B.; Pinder, R.D.; Lord, W.D. Insect larvae used to detect cocaine poisoning in a decomposed body. J. Forensic Sci. 1992, 37, 1179–1185. [Google Scholar] [CrossRef]
- Introna, F.; Di Vella, G.; Gagliano Candela, R. Identification d’opiacés sur les pupes vides: Contribution expérimentale. J. Med. Leg. Droit. Med. 2001, 44, 211–215. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magni, P.A.; Harvey, A.D.; Guareschi, E.E. Insects Associated with Ancient Human Remains: How Archaeoentomology Can Provide Additional Information in Archaeological Studies. Heritage 2023, 6, 435-465. https://doi.org/10.3390/heritage6010023
Magni PA, Harvey AD, Guareschi EE. Insects Associated with Ancient Human Remains: How Archaeoentomology Can Provide Additional Information in Archaeological Studies. Heritage. 2023; 6(1):435-465. https://doi.org/10.3390/heritage6010023
Chicago/Turabian StyleMagni, Paola Annarosa, Abigail Dianne Harvey, and Edda Emanuela Guareschi. 2023. "Insects Associated with Ancient Human Remains: How Archaeoentomology Can Provide Additional Information in Archaeological Studies" Heritage 6, no. 1: 435-465. https://doi.org/10.3390/heritage6010023
APA StyleMagni, P. A., Harvey, A. D., & Guareschi, E. E. (2023). Insects Associated with Ancient Human Remains: How Archaeoentomology Can Provide Additional Information in Archaeological Studies. Heritage, 6(1), 435-465. https://doi.org/10.3390/heritage6010023