Working around the Clock: Is a Person’s Endogenous Circadian Timing for Optimal Neurobehavioral Functioning Inherently Task-Dependent?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
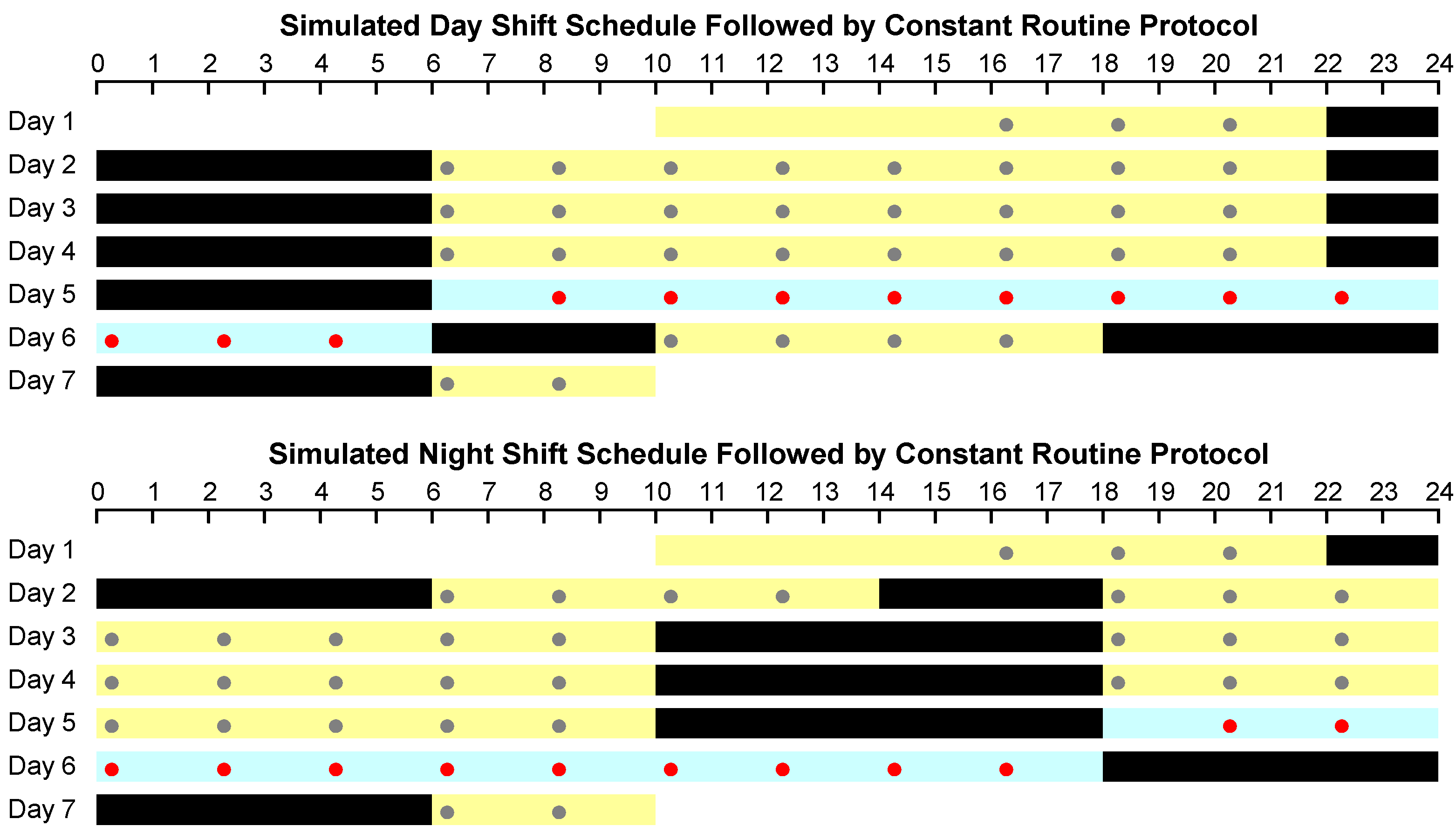
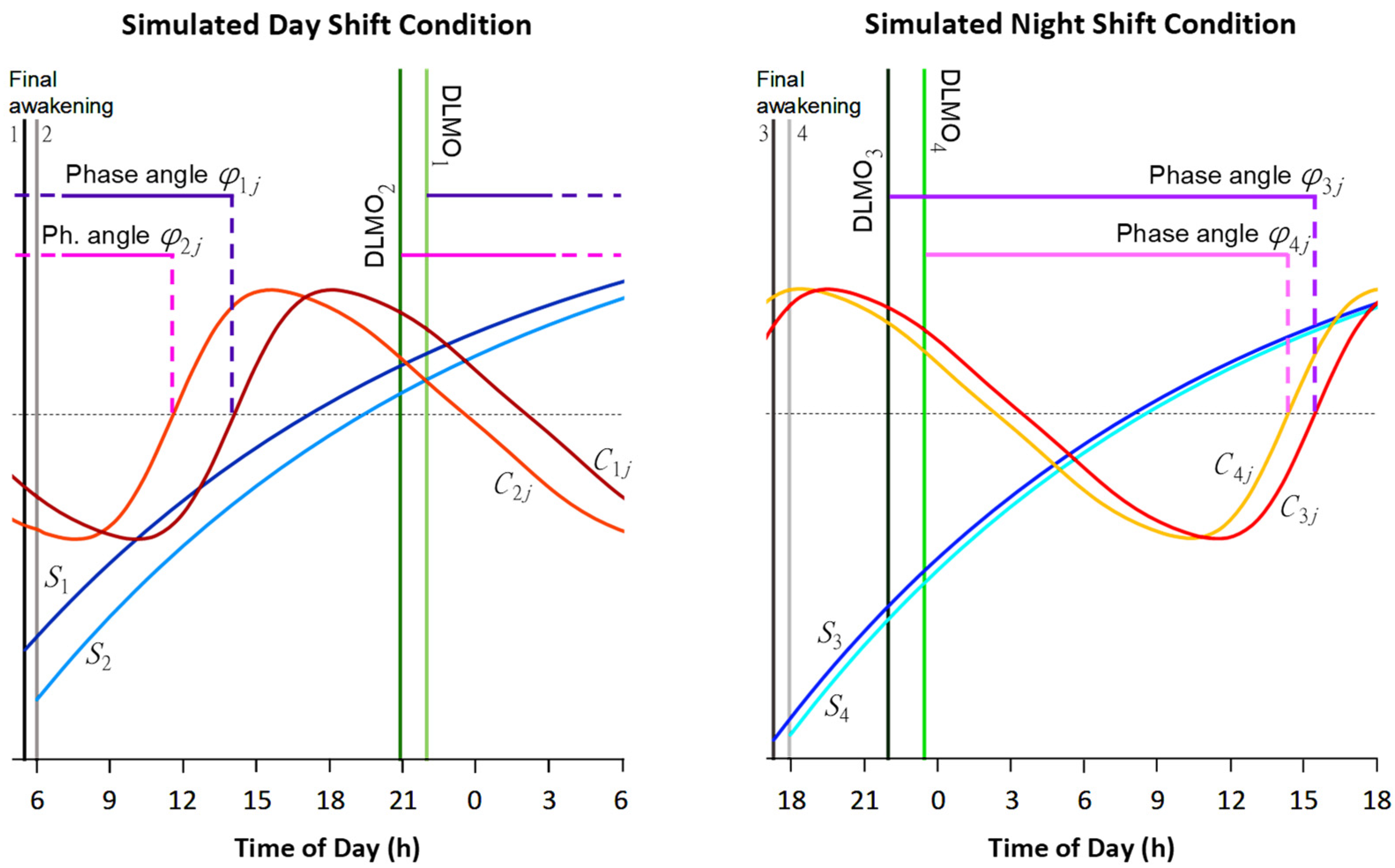
2.2. Experimental Design
2.3. Neurobehavioral Assays
2.4. Statistical Methods
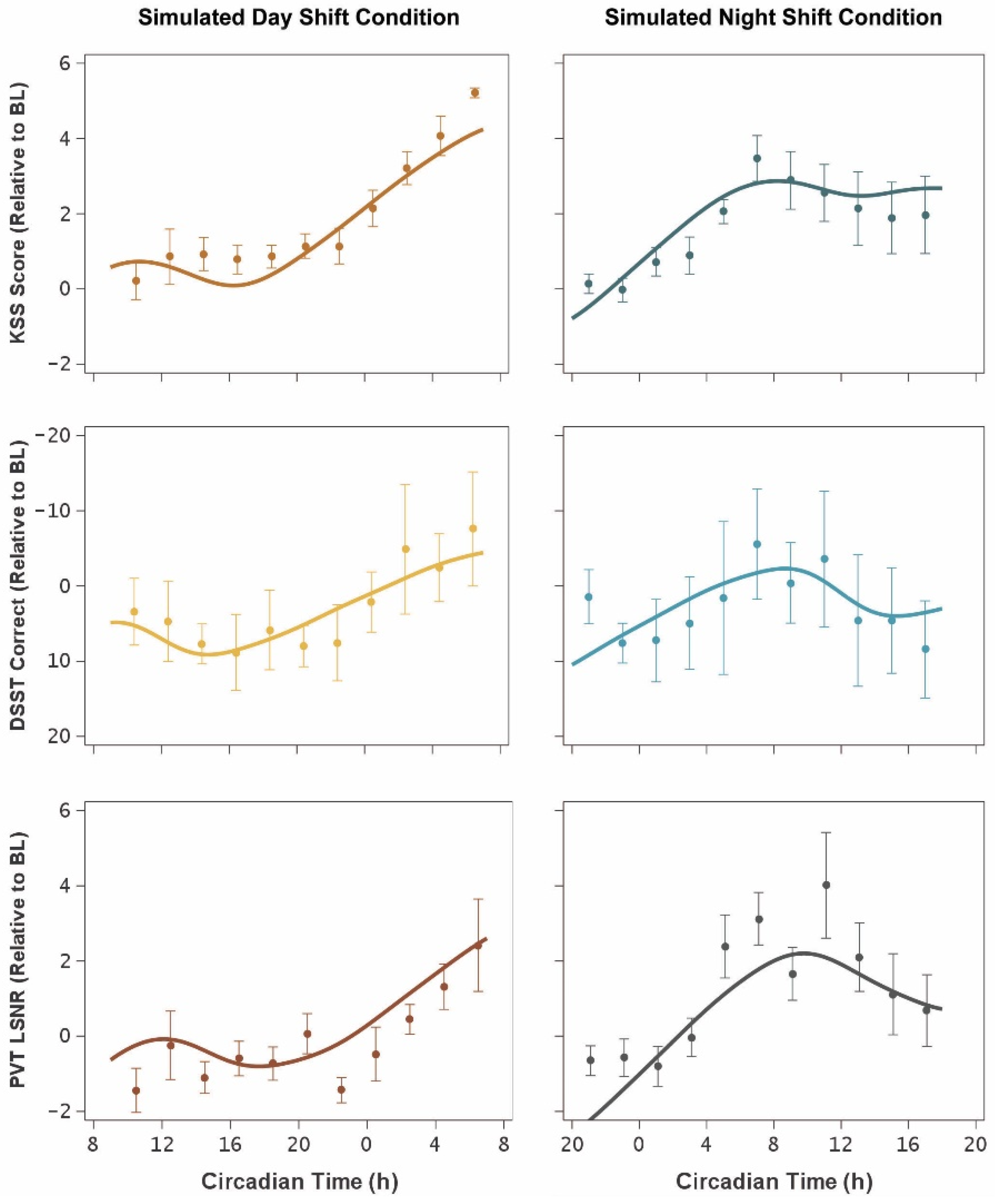
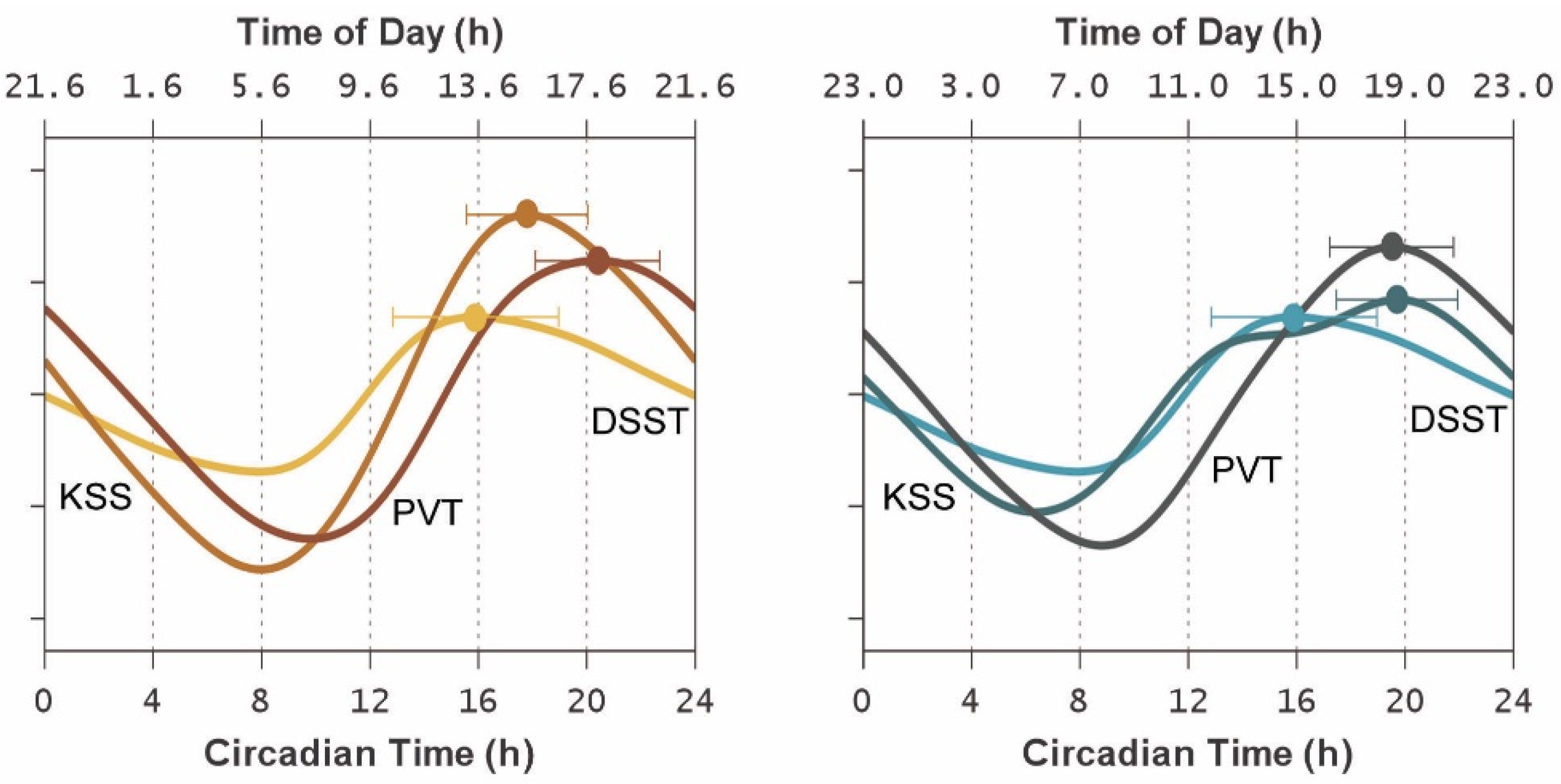
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daan, S.; Beersma, D.G.; Borbély, A.A. Timing of human sleep: Recovery process gated by a circadian pacemaker. Am. J. Physiol. 1984, 246, R161–R183. [Google Scholar] [CrossRef] [PubMed]
- Dijk, D.J.; Duffy, J.F.; Czeisler, C.A. Circadian and sleep/wake dependent aspects of subjective alertness and cognitive performance. J. Sleep Res. 1992, 1, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Dijk, D.J.; Czeisler, C.A. Paradoxical timing of the circadian rhythm of sleep propensity serves to consolidate sleep and wakefulness in humans. Neurosci. Lett. 1994, 166, 62–68. [Google Scholar] [CrossRef]
- Van Dongen, H.P.A.; Dinges, D.F. Sleep, circadian rhythms, and psychomotor vigilance. Clin. Sports Med. 2005, 24, 237–249. [Google Scholar] [CrossRef]
- Wilson, M.; Permito, R.; English, A.; Albritton, S.; Coogle, C.; Van Dongen, H.P.A. Performance and sleepiness in nurses working 12-h day shifts or night shifts in a community hospital. Accid. Anal. Prev. 2019, 126, 43–46. [Google Scholar] [CrossRef]
- Gurubhagavatula, I.; Barger, L.K.; Barnes, C.M.; Basner, M.; Boivin, D.B.; Dawson, D.; Drake, C.L.; Flynn-Evans, E.E.; Mysliwiec, V.; Patterson, P.D.; et al. Guiding principles for determining work shift duration and addressing the effects of work shift duration on performance, safety, and health. Sleep 2021, 15, zsab161. [Google Scholar] [CrossRef]
- Van Dongen, H.P.A.; Bender, A.M.; Dinges, D.F. Systematic individual differences in sleep homeostatic and circadian rhythm contributions to neurobehavioral impairment during sleep deprivation. Accid. Anal. Prev. 2012, 45, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Rusterholz, T.; Tarokh, L.; Van Dongen, H.P.A.; Achermann, P. Interindividual differences in the dynamics of the homeostatic process are trait-like and distinct for sleep versus wakefulness. J. Sleep Res. 2017, 26, 171–178. [Google Scholar] [CrossRef]
- Achermann, P.; Borbély, A.A. Simulation of daytime vigilance by the additive interaction of a homeostatic and a circadian process. Biol. Cybern. 1994, 71, 115–121. [Google Scholar] [CrossRef]
- Van Dongen, H.P.A.; Dinges, D.F. Investigating the interaction between the homeostatic and circadian processes of sleep-wake regulation for the prediction of waking neurobehavioral performance. J. Sleep Res. 2003, 12, 181–187. [Google Scholar] [CrossRef]
- Mallis, M.M.; Mejdal, S.; Nguyen, T.T.; Dinges, D.F. Summary of the key features of seven biomathematical models of human fatigue and performance. Aviat. Space Environ. Med. 2004, 75, A4–A14. [Google Scholar] [PubMed]
- Riedy, S.M.; Hursh, S.R.; Dawson, D.; Balkin, T.J.; Van Dongen, H.P.A. Sleep, sleepiness, and performance prediction modeling. In Principles and Practice of Sleep Medicine, 7th ed.; Kryger, M., Roth, T., Goldstein, C.A., Dement, W.C., Eds.; Elsevier: Philadelphia, PA, USA, 2022; pp. 780–787. [Google Scholar]
- Knock, S.A.; Magee, M.; Stone, J.E.; Ganesan, S.; Mulhall, M.D.; Lockley, S.W.; Howard, M.E.; Rajaratnam, S.M.W.; Sletten, T.L.; Postnova, S. Prediction of shiftworker alertness, sleep, and circadian phase using a model of arousal dynamics constrained by shift schedules and light exposure. Sleep 2021, 44, zsab146. [Google Scholar] [CrossRef] [PubMed]
- McCauley, M.E.; McCauley, P.; Riedy, S.M.; Banks, S.; Ecker, A.J.; Kalachev, L.V.; Rangan, S.; Dinges, D.F.; Van Dongen, H.P.A. Fatigue risk management based on self-reported fatigue: Expanding a biomathematical model of fatigue-related performance deficits to also predict subjective sleepiness. Transp. Res. Part F Traffic Psychol. Behav. 2021, 79, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Rangan, S.; Riedy, S.M.; Bassett, R.; Klinck, Z.A.; Hagerty, P.; Schek, E.; Zhang, Y.; Hursh, S.R.; Van Dongen, H.P.A. Predictive and proactive fatigue risk management approaches in commercial aviation. Chronobiol. Int. 2020, 37, 1479–1482. [Google Scholar] [CrossRef]
- Vital-Lopez, F.G.; Doty, T.J.; Reifman, J. Optimal sleep and work schedules to maximize alertness. Sleep 2021, 44, zsab144. [Google Scholar] [CrossRef]
- Hockey, G.R.J.; Colquhoun, W.P. Diurnal variation in human performance: A review. In Aspects of Human Efficiency: Diurnal Rhythm and Loss of Sleep; Colquhoun, W.P., Ed.; English Universities Press: London, UK, 1972; pp. 1–23. [Google Scholar]
- Kirkcaldy, B.D. Performance and circadian rhythms. Eur. J. Appl. Physiol. Occup. Physiol. 1984, 52, 375–379. [Google Scholar] [CrossRef]
- Folkard, S.; Hume, K.; Minors, D.; Waterhouse, J.M.; Watson, F.L. Independence of the circadian rhythm in alertness from the sleep/wake cycle. Nature 1985, 313, 678–679. [Google Scholar] [CrossRef]
- Monk, T.H.; Fookson, J.E.; Moline, M.L.; Pollak, C.P. Diurnal variation in mood and performance in a time-isolated environment. Chronobiol. Int. 1985, 2, 185–193. [Google Scholar] [CrossRef]
- Monk, T.H.; Leng, V.C. Interactions between inter-individual and inter-task differences in the diurnal variation of human performance. Chronobiol. Int. 1986, 3, 171–177. [Google Scholar] [CrossRef]
- Folkard, S. Diurnal variation in logical reasoning. Br. J. Psychol. 1975, 66, 1–8. [Google Scholar] [CrossRef]
- Folkard, S.; Monk, T.H. Circadian rhythms in human memory. Br. J. Psychol. 1980, 71, 295–307. [Google Scholar] [CrossRef]
- Colquhoun, W.P. Rhythms in performance. In Handbook of Behavioral Neurobiology; Aschoff, J., Ed.; Plenum Press: New York, NY, USA, 1981; Volume 4, pp. 333–348. [Google Scholar]
- Folkard, S.; Wever, R.A.; Wildgruber, C.M. Multi-oscillatory control of circadian rhythms in human performance. Nature 1983, 305, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Monk, T.H.; Weitzman, E.D.; Fookson, J.E.; Moline, M.L.; Kronauer, R.E.; Gander, P.H. Task variables determine which biological clock controls circadian rhythms in human performance. Nature 1983, 304, 543–545. [Google Scholar] [CrossRef] [PubMed]
- Muto, V.; Jasper, M.; Meyer, C.; Kussé, C.; Chellappa, S.L.; Degueldre, C.; Balteau, E.; Shaffii-Le Bourdiec, A.; Luxen, A.; Middleton, B.; et al. Local modulation of human brain responses by circadian rhythmicity and sleep debt. Science 2016, 353, 687–690. [Google Scholar] [CrossRef]
- Johnson, M.P.; Duffy, J.F.; Dijk, D.J.; Ronda, J.M.; Dyal, C.M.; Czeisler, C.A. Short-term memory, alertness and performance: A reappraisal of their relationship to body temperature. J. Sleep Res. 1992, 1, 24–29. [Google Scholar] [CrossRef]
- Monk, T.H.; Buysse, D.J.; Reynolds, C.F., III; Berga, S.L.; Jarrett, D.B.; Begley, A.E.; Kupfer, D.J. Circadian rhythms in human performance and mood under constant conditions. J. Sleep Res. 1997, 6, 9–18. [Google Scholar] [CrossRef]
- Owens, D.S.; Macdonald, I.; Tucker, P.; Sytnik, N.; Minors, D.; Waterhouse, J.; Totterdell, P.; Folkard, S. Diurnal trends in mood and performance do not all parallel alertness. Scand. J. Work Environ. Health 1998, 24 (Suppl. 3), 109–114. [Google Scholar]
- Chellappa, S.L.; Morris, C.J.; Scheer, F.A.J.L. Daily circadian misalignment impairs human cognitive performance task-dependently. Sci. Rep. 2018, 8, 3041. [Google Scholar] [CrossRef] [Green Version]
- McMahon, W.R.; Ftouni, S.; Drummond, S.P.A.; Maruff, P.; Lockley, S.W.; Rajaratnam, S.M.W.; Anderson, C. The wake maintenance zone shows task dependent changes in cognitive function following one night without sleep. Sleep 2018, 41, zsy148. [Google Scholar] [CrossRef]
- Collet, J.; Ftouni, S.; Clough, M.; Cain, S.W.; Fielding, J.; Anderson, C. Differential impact of sleep deprivation and circadian timing on reflexive versus inhibitory control of attention. Sci. Rep. 2020, 10, 7270. [Google Scholar] [CrossRef]
- Skeiky, L.; Hudson, A.N.; Van Dongen, H.P.A. Circadian rhythms in sleepiness, alertness, and performance. In Principles and Practice of Sleep Medicine, 7th ed.; Kryger, M., Roth, T., Goldstein, C.A., Dement, W.C., Eds.; Elsevier: Philadelphia, PA, USA, 2022; pp. 407–414. [Google Scholar]
- Bendat, J.S.; Piersol, A.G. Random Data: Analysis and Measurement Procedures, 4th ed.; John Wiley & Sons: New York, NY, USA, 2010. [Google Scholar]
- Van Dongen, H.P.A.; Baynard, M.D.; Maislin, G.; Dinges, D.F. Systematic interindividual differences in neurobehavioral impairment from sleep loss: Evidence of trait-like differential vulnerability. Sleep 2004, 27, 423–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dongen, H.P.A.; Caldwell, J.A.; Caldwell, J.L. Investigating systematic individual differences in sleep-deprived performance on a high-fidelity flight simulator. Behav. Res. Meth. 2006, 38, 333–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dongen, H.P.A.; Belenky, G.; Krueger, J.M. A local, bottom-up perspective on sleep deprivation and neurobehavioral performance. Curr. Top. Med. Chem. 2011, 11, 2414–2422. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Ferguson, S.A.; Matthews, R.W.; Sargent, C.; Darwent, D.; Kennaway, D.J.; Roach, G.D. Interindividual differences in neurobehavioral performance in response to increasing homeostatic sleep pressure. Chronobiol. Int. 2010, 27, 922–933. [Google Scholar] [CrossRef] [PubMed]
- Honn, K.A.; Halverson, T.; Jackson, M.L.; Krusmark, M.; Chavali, V.P.; Gunzelmann, G.; Van Dongen, H.P.A. New insights into the cognitive effects of sleep deprivation by decomposition of a cognitive throughput task. Sleep 2020, 43, zsz319. [Google Scholar] [CrossRef] [PubMed]
- Folkard, S.; Monk, T.H. Time of day and processing strategy in free recall. Q. J. Exp. Psychol. 1979, 31, 461–475. [Google Scholar] [CrossRef]
- Hudson, A.N.; Hansen, D.A.; Hinson, J.M.; Whitney, P.; Layton, M.E.; DePriest, D.M.; Van Dongen, H.P.A.; Honn, K.A. Speed/accuracy trade-off in the effects of acute total sleep deprivation on a sustained attention and response inhibition task. Chronobiol. Int. 2020, 37, 1441–1444. [Google Scholar] [CrossRef]
- Duffy, J.F.; Dijk, D.J. Getting through to circadian oscillators: Why use constant routines? J. Biol. Rhythms 2002, 17, 4–13. [Google Scholar] [CrossRef]
- Lim, J.; Dinges, D.F. Sleep deprivation and vigilant attention. Ann. N. Y. Acad. Sci. 2008, 1129, 305–322. [Google Scholar] [CrossRef]
- Van Dongen, H.P.A.; Maislin, G.; Mullington, J.M.; Dinges, D.F. The cumulative cost of additional wakefulness: Dose-response effects on neurobehavioral functions and sleep physiology from chronic sleep restriction and total sleep deprivation. Sleep 2003, 26, 117–126. [Google Scholar] [CrossRef]
- Åkerstedt, T.; Anund, A.; Axelsson, J.; Kecklund, G. Subjective sleepiness is a sensitive indicator of insufficient sleep and impaired waking function. J. Sleep Res. 2014, 23, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Olofsen, E.; Dinges, D.F.; Van Dongen, H.P.A. Nonlinear mixed-effects modeling: Individualization and prediction. Aviat. Space Environ. Med. 2004, 75, A134–A140. [Google Scholar] [PubMed]
- Smith, C.S.; Reilly, C.; Midkiff, K. Evaluation of three circadian rhythm questionnaires with suggestions for an improved measure of morningness. J. Appl. Psychol. 1989, 74, 728–738. [Google Scholar] [CrossRef] [PubMed]
- Iber, C.; Ancoli-Israel, S.; Chesson, A.L., Jr.; Quan, S.F. The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology, and Technical Specifications, 1st ed.; American Academy of Sleep Medicine: Westchester, IL, USA, 2007. [Google Scholar]
- Skornyakov, E.; Gaddameedhi, S.; Paech, G.M.; Sparrow, A.R.; Satterfield, B.C.; Shattuck, N.L.; Layton, M.E.; Karatsoreos, I.; Van Dongen, H.P.A. Cardiac autonomic activity during simulated shift work. Ind. Health 2019, 57, 118–132. [Google Scholar] [CrossRef] [Green Version]
- Skene, D.; Skornyakov, E.; Chowdhury, N.; Gajula, R.P.; Middleton, B.; Satterfield, B.C.; Porter, K.I.; Van Dongen, H.P.A.; Gaddameedhi, S. Separation of circadian- and behavior-driven metabolite rhythms in humans provides a window on peripheral oscillators and metabolism. Proc. Natl. Acad. Sci. USA 2018, 115, 7825–7830. [Google Scholar] [CrossRef] [Green Version]
- Åkerstedt, T.; Gillberg, M. Subjective and objective sleepiness in the active individual. Int. J. Neurosci. 1990, 52, 29–37. [Google Scholar] [CrossRef]
- Wechsler, D. Wechsler Adult Intelligence Scale–Revised; Psychological Corp.: New York, NY, USA, 1981. [Google Scholar]
- Dinges, D.F.; Powell, J.W. Microcomputer analyses of performance on a portable, simple visual RT task during sustained operations. Behav. Res. Methods Instrum. Comput. 1985, 17, 652–655. [Google Scholar] [CrossRef]
- Dorrian, J.; Rogers, N.L.; Dinges, D.F. Psychomotor vigilance performance: Neurocognitive assay sensitive to sleep loss. In Sleep Deprivation: Clinical Issues, Pharmacology, and Sleep Loss Effects; Kushida, C.A., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 2005; pp. 39–70. [Google Scholar]
- Chavali, V.P.; Riedy, S.M.; Van Dongen, H.P.A. Signal-to-noise ratio in PVT performance as a cognitive measure of the effect of sleep deprivation on the fidelity of information processing. Sleep 2017, 40, zsx016. [Google Scholar] [CrossRef]
- Lundholm, K.R.; Honn, K.A.; Skeiky, L.; Muck, R.A.; Van Dongen, H.P.A. Trait interindividual differences in the magnitude of subjective sleepiness from sleep inertia. Clocks Sleep 2021, 3, 298–311. [Google Scholar] [CrossRef]
- Borbély, A.A.; Achermann, P. Sleep homeostasis and models of sleep regulation. J. Biol. Rhythm. 1999, 14, 557–568. [Google Scholar] [CrossRef]
- Kerkhof, G.A.; Van Dongen, H.P.A. Morning-type and evening-type individuals differ in the phase position of their endogenous circadian oscillator. Neurosci. Lett. 1996, 218, 153–156. [Google Scholar] [CrossRef]
- Kantermann, T.; Eastman, C.I. Circadian phase, circadian period and chronotype are reproducible over months. Chronobiol. Int. 2018, 35, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Eastman, C.I.; Liu, L.; Fogg, L.F. Circadian rhythm adaptation to simulated night shift work: Effect of nocturnal bright-light duration. Sleep 1995, 18, 399–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strogatz, S.H.; Kronauer, R.E.; Czeisler, C.A. Circadian pacemaker interferes with sleep onset at specific times each day: Role in insomnia. Am. J. Physiol. 1987, 253, R172–R178. [Google Scholar] [CrossRef] [Green Version]
- Lavie, P. Ultrashort sleep-waking schedule. III. ‘Gates’ and ‘forbidden zones’ for sleep. Electroencephalogr. Clin. Neurophysiol. 1986, 63, 414–425. [Google Scholar] [CrossRef]
- Shekleton, J.A.; Rajaratnam, S.M.; Gooley, J.J.; Van Reen, E.; Czeisler, C.A.; Lockley, S.W. Improved neurobehavioral performance during the wake maintenance zone. J. Clin. Sleep Med. 2013, 9, 353–362. [Google Scholar] [CrossRef] [Green Version]
- De Zeeuw, J.; Wisniewski, S.; Papakonstantinou, A.; Bes, F.; Wahnschaffe, A.; Zaleska, M.; Kunz, D.; Münch, M. The alerting effect of the wake maintenance zone during 40 hours of sleep deprivation. Sci. Rep. 2018, 8, 11012. [Google Scholar] [CrossRef] [Green Version]
- Mackie, J.L. Causes and conditions. Am. Philos. Q. 1965, 2, 245–264. [Google Scholar]
- Rubin, D.B. Estimating causal effects of treatments in randomized and nonrandomized studies. J. Educ. Psychol. 1974, 66, 688–701. [Google Scholar] [CrossRef] [Green Version]
- Santhi, N.; Lazar, A.S.; McCabe, P.J.; Lo, J.C.; Groeger, J.A.; Dijk, D.J. Sex differences in the circadian regulation of sleep and waking cognition in humans. Proc. Natl. Acad. Sci. USA 2016, 113, E2730–E2739. [Google Scholar] [CrossRef] [Green Version]
- Dijk, D.J.; Landolt, H.P. Sleep physiology, circadian rhythms, waking performance and the development of sleep-wake therapeutics. Handb. Exp. Pharmacol. 2019, 253, 441–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitney, P.; Hinson, J.M. Measurement of cognition in studies of sleep deprivation. Prog. Brain Res. 2010, 185, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Whitney, P.; Hinson, J.M.; Satterfield, B.C.; Grant, D.A.; Honn, K.A.; Van Dongen, H.P.A. Sleep deprivation diminishes attentional control effectiveness and impairs flexible adaptation to changing conditions. Sci. Rep. 2017, 7, 16020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurinec, C.A.; Whitney, P.; Hinson, J.M.; Hansen, D.A.; Van Dongen, H.P.A. Sleep deprivation impairs binding of information with its context. Sleep 2021, 44, zsab113. [Google Scholar] [CrossRef]
- Stepan, M.E.; Fenn, K.M.; Altmann, E.M. Effects of sleep deprivation on procedural errors. J. Exp. Psychol. Gen. 2019, 148, 1828–1833. [Google Scholar] [CrossRef]
- Stenson, A.R.; Kurinec, C.A.; Hinson, J.M.; Whitney, P.; Van Dongen, H.P.A. Total sleep deprivation reduces top-down regulation of emotion without altering bottom-up affective processing. PLoS ONE 2021, 16, e0256983. [Google Scholar] [CrossRef]
- Krueger, J.M.; Huang, Y.H.; Rector, D.M.; Buysse, D.J. Sleep: A synchrony of cell activity-driven small network states. Eur. J. Neurosci. 2013, 38, 2199–2209. [Google Scholar] [CrossRef] [Green Version]
- Nir, Y.; Andrillon, T.; Marmelshtein, A.; Suthana, N.; Cirelli, C.; Tononi, G.; Fried, I. Selective neuronal lapses precede human cognitive lapses following sleep deprivation. Nat. Med. 2017, 23, 1474–1480. [Google Scholar] [CrossRef]
- Thomas, C.W.; Guillaumin, M.C.; McKillop, L.E.; Achermann, P.; Vyazovskiy, V.V. Global sleep homeostasis reflects temporally and spatially integrated local cortical neuronal activity. eLife 2020, 9, e54148. [Google Scholar] [CrossRef]
- Smith, M.R.; Eastman, C.I. Shift work: Health, performance and safety problems, traditional countermeasures, and innovative management strategies to reduce circadian misalignment. Nat. Sci. Sleep 2012, 4, 111–132. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, S.; Magee, M.; Stone, J.E.; Mulhall, M.D.; Collins, A.; Howard, M.E.; Lockley, S.W.; Rajaratnam, S.M.W.; Sletten, T.L. The impact of shift work on sleep, alertness and performance in healthcare workers. Sci. Rep. 2019, 9, 4635. [Google Scholar] [CrossRef] [PubMed]
- Rosa, D.E.; Marot, L.P.; de Mello, M.T.; Marqueze, E.C.; Narciso, F.V.; de Araújo, L.B.; Crispim, C.A. Association between chronotype and psychomotor performance of rotating shift workers. Sci. Rep. 2021, 11, 6919. [Google Scholar] [CrossRef] [PubMed]
- Raslear, T.G.; Hursh, S.R.; Van Dongen, H.P.A. Predicting cognitive impairment and accident risk. Prog. Brain Res. 2011, 190, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Wong, I.S.; Popkin, S.; Folkard, S. Working Time Society consensus statements: A multi-level approach to managing occupational sleep-related fatigue. Ind. Health 2019, 57, 228–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dongen, H.P.A.; Belenky, G. Individual differences in vulnerability to sleep loss in the work environment. Ind. Health 2009, 47, 518–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, R.Y. The suprachiasmatic nucleus and the circadian timing system. Prog. Mol. Biol. Transl. Sci. 2013, 119, 1–28. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muck, R.A.; Hudson, A.N.; Honn, K.A.; Gaddameedhi, S.; Van Dongen, H.P.A. Working around the Clock: Is a Person’s Endogenous Circadian Timing for Optimal Neurobehavioral Functioning Inherently Task-Dependent? Clocks & Sleep 2022, 4, 23-36. https://doi.org/10.3390/clockssleep4010005
Muck RA, Hudson AN, Honn KA, Gaddameedhi S, Van Dongen HPA. Working around the Clock: Is a Person’s Endogenous Circadian Timing for Optimal Neurobehavioral Functioning Inherently Task-Dependent? Clocks & Sleep. 2022; 4(1):23-36. https://doi.org/10.3390/clockssleep4010005
Chicago/Turabian StyleMuck, Rachael A., Amanda N. Hudson, Kimberly A. Honn, Shobhan Gaddameedhi, and Hans P. A. Van Dongen. 2022. "Working around the Clock: Is a Person’s Endogenous Circadian Timing for Optimal Neurobehavioral Functioning Inherently Task-Dependent?" Clocks & Sleep 4, no. 1: 23-36. https://doi.org/10.3390/clockssleep4010005
APA StyleMuck, R. A., Hudson, A. N., Honn, K. A., Gaddameedhi, S., & Van Dongen, H. P. A. (2022). Working around the Clock: Is a Person’s Endogenous Circadian Timing for Optimal Neurobehavioral Functioning Inherently Task-Dependent? Clocks & Sleep, 4(1), 23-36. https://doi.org/10.3390/clockssleep4010005