Critical Neural Networks Minimize Metabolic Cost

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Problem Statement
1.2. Contributions
2. Methods
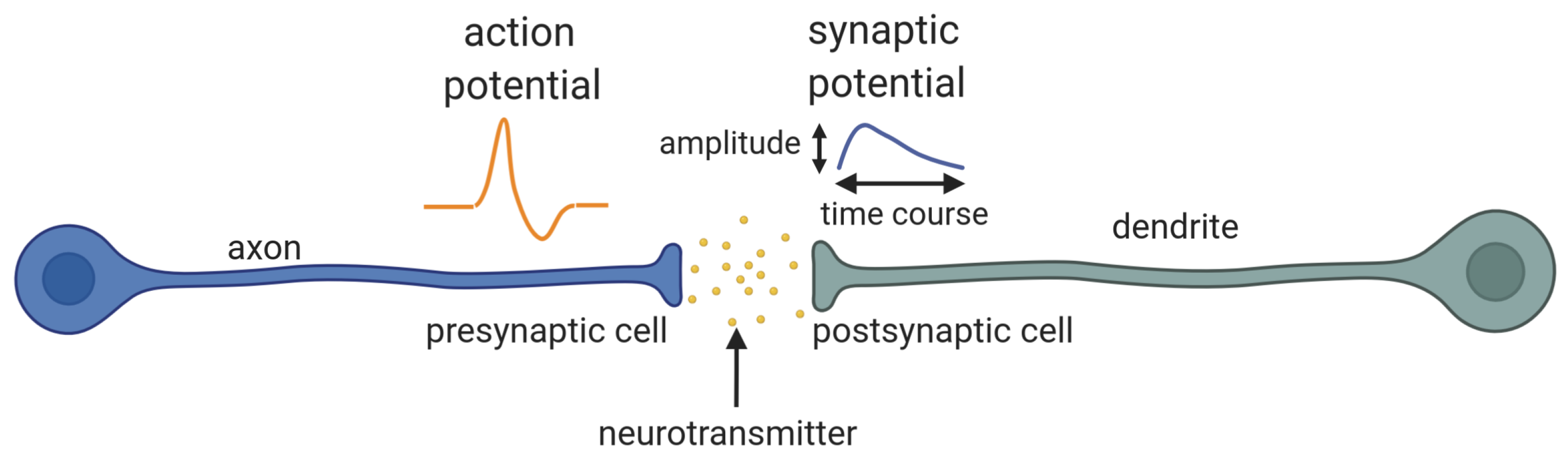
2.1. Izhikevich Neuron Model
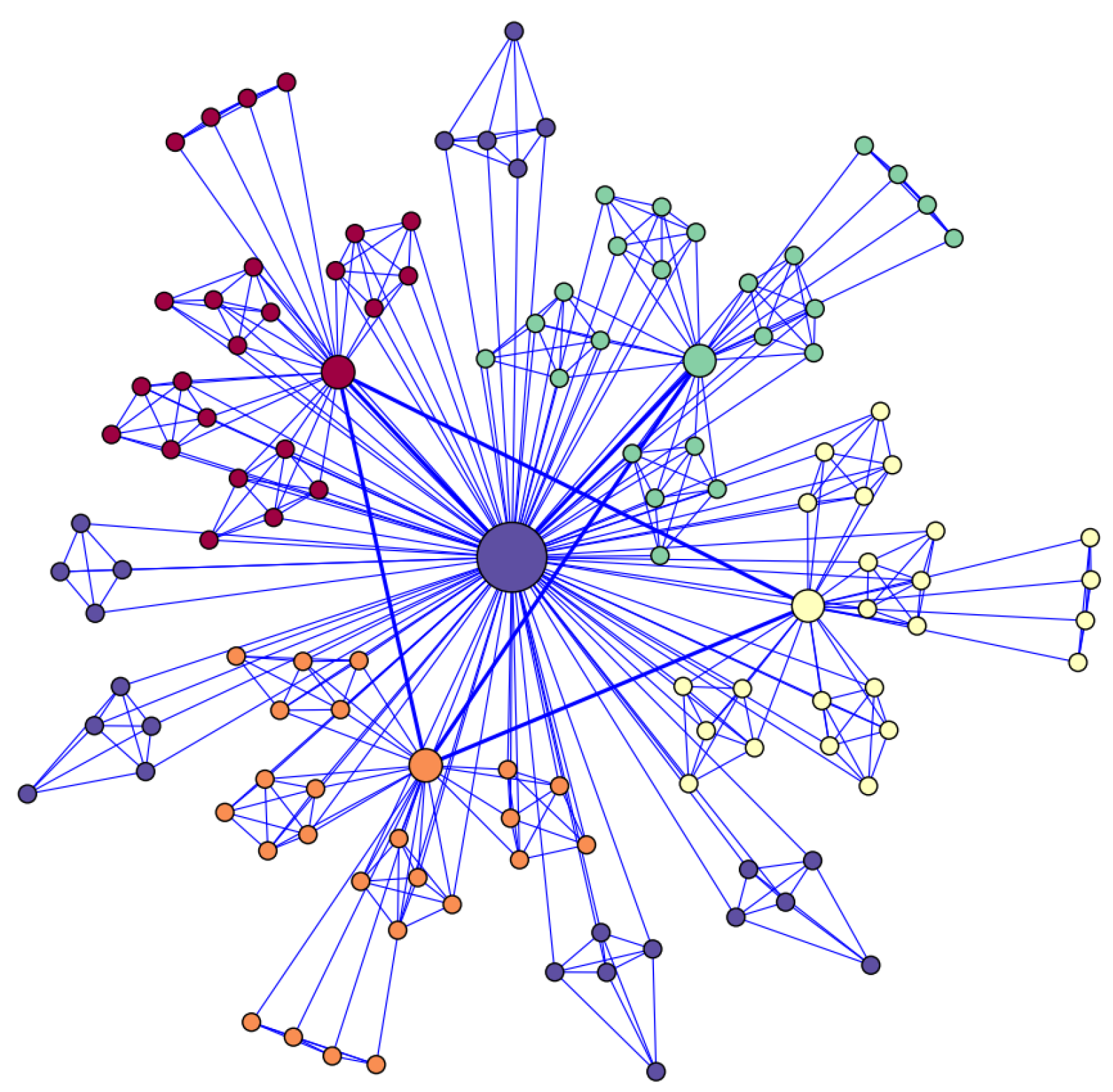
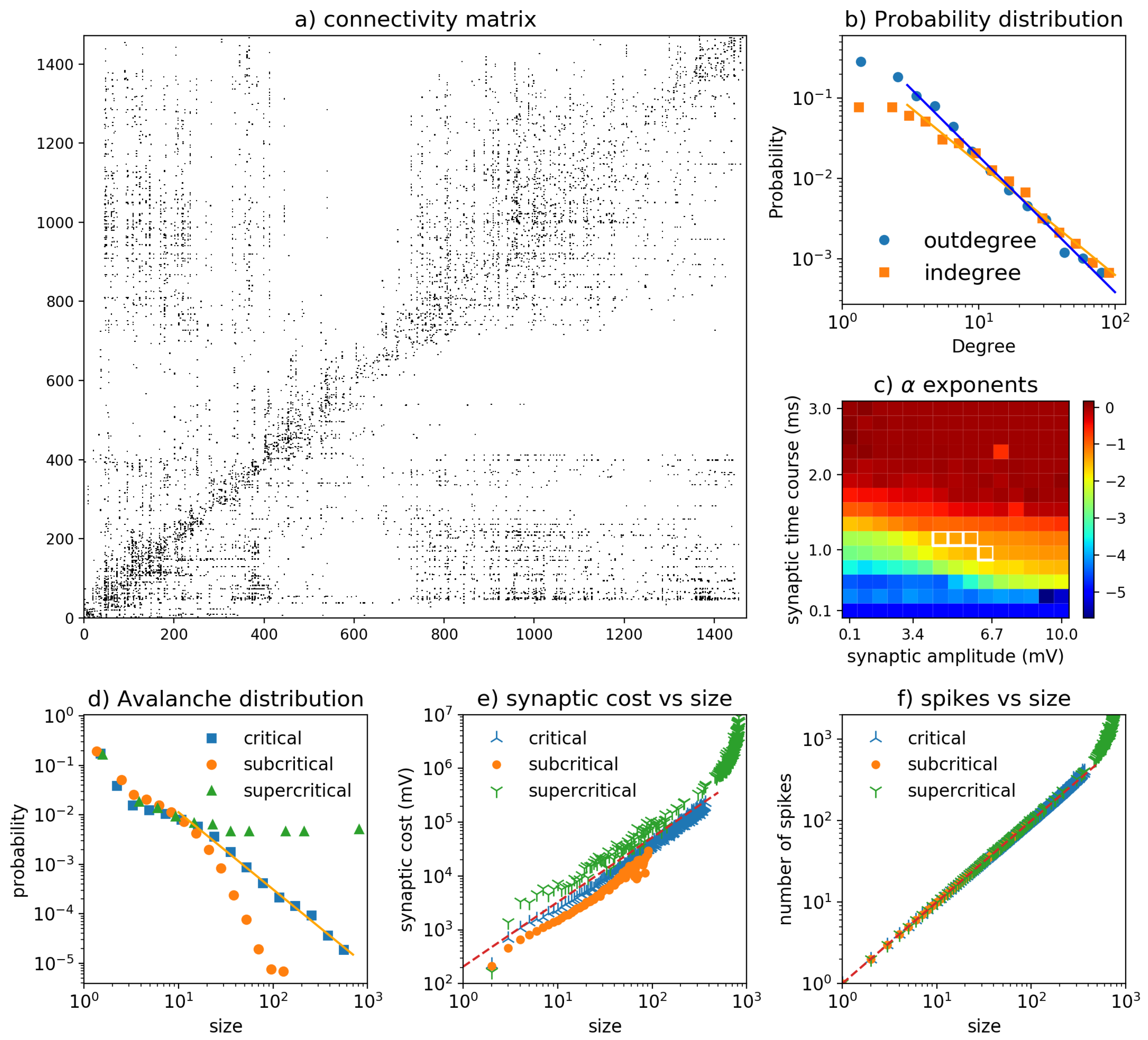
2.2. Hierarchical Network and Rich-Club Organization
2.3. Goodness-of-Fit Test
3. Results
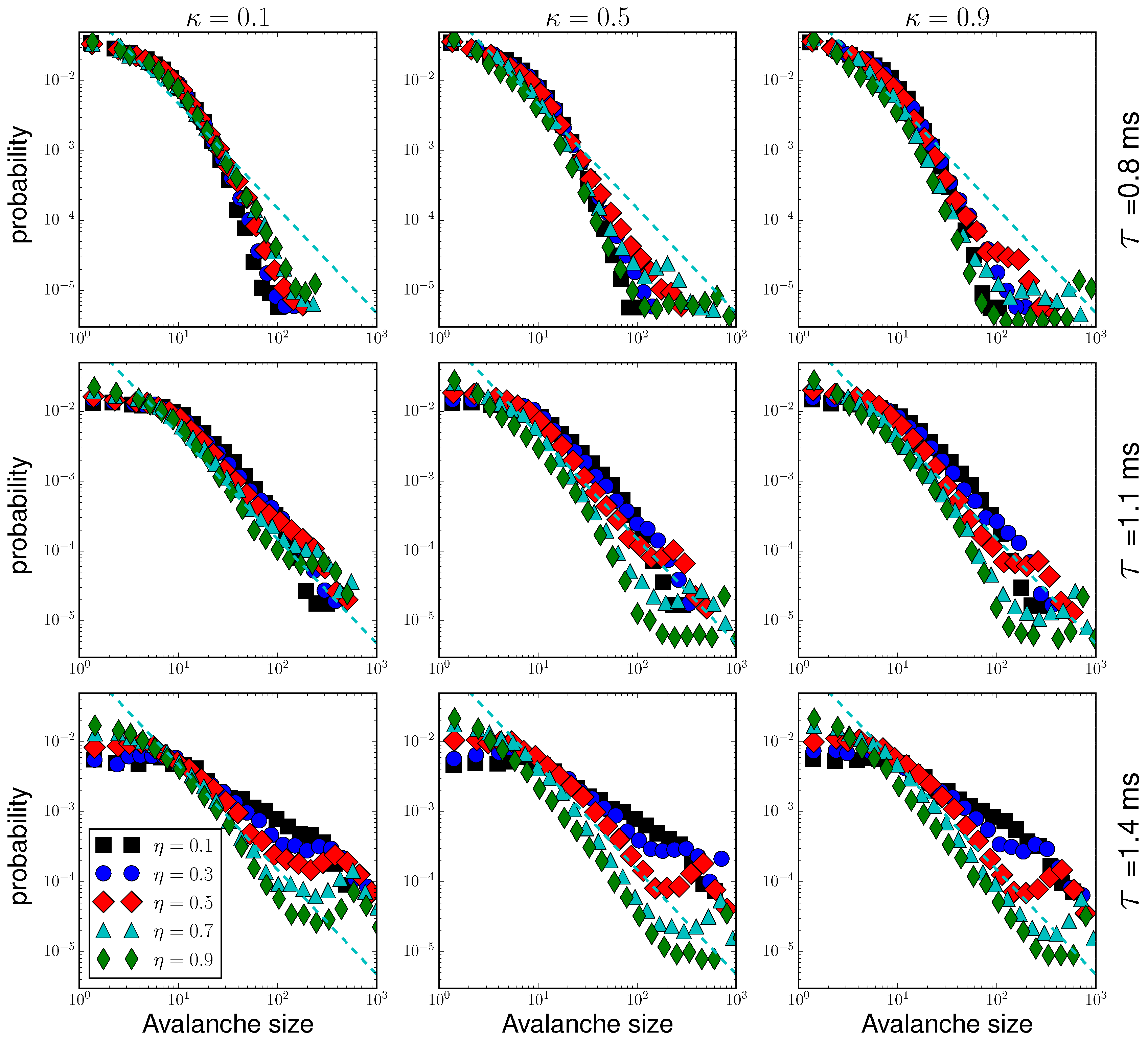
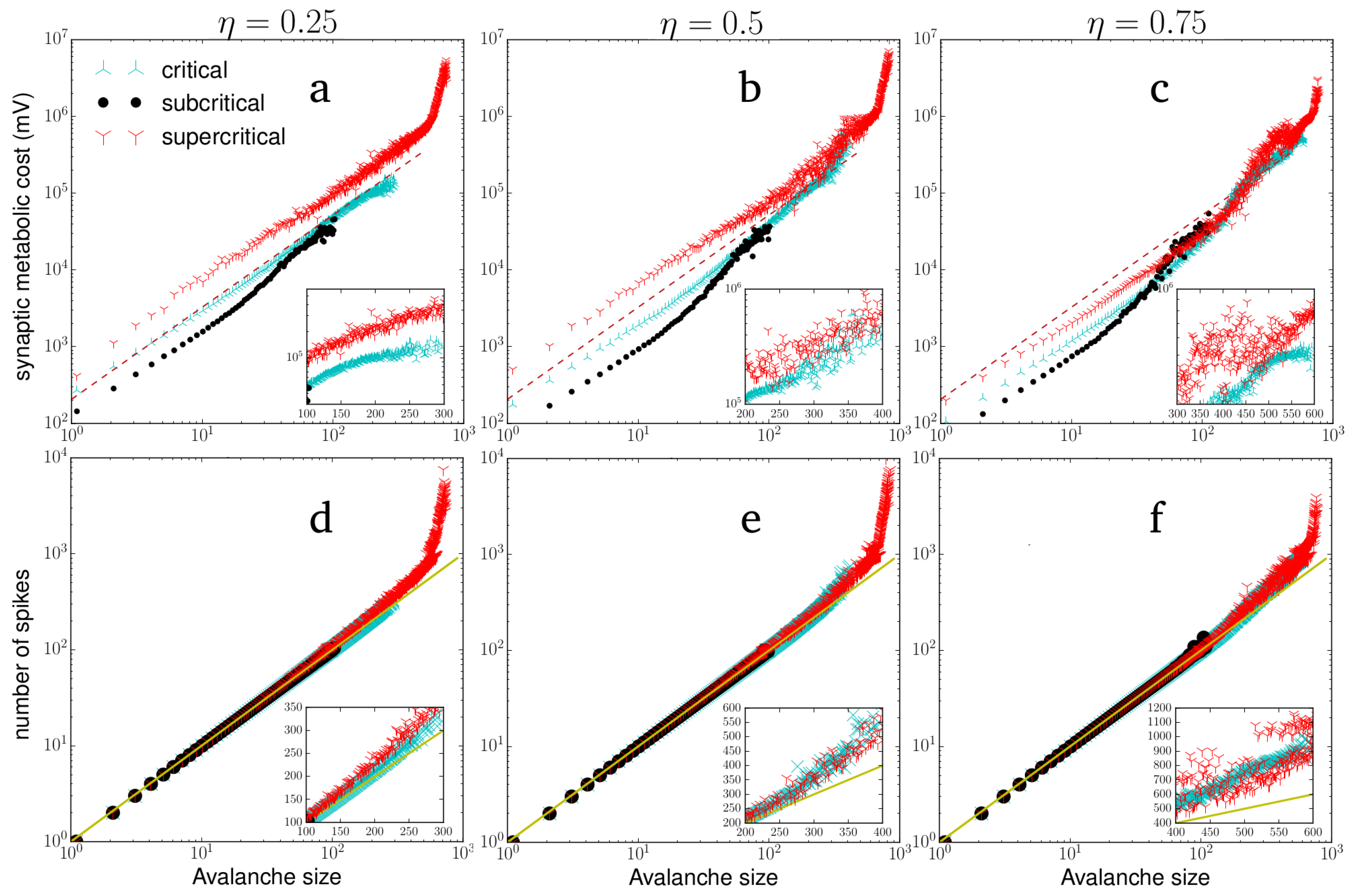
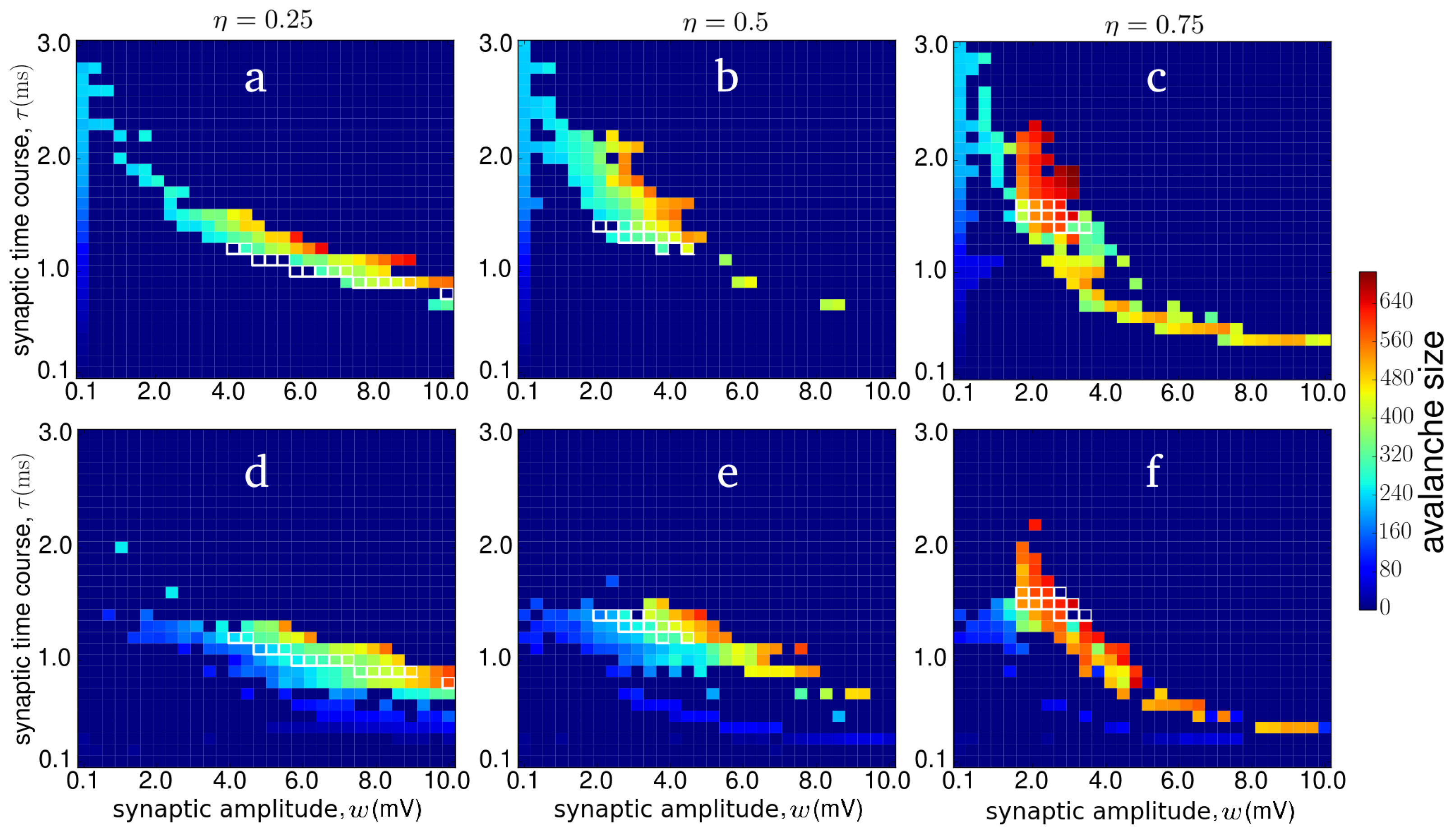
3.1. Criticality in Rich-Club Neural Networks
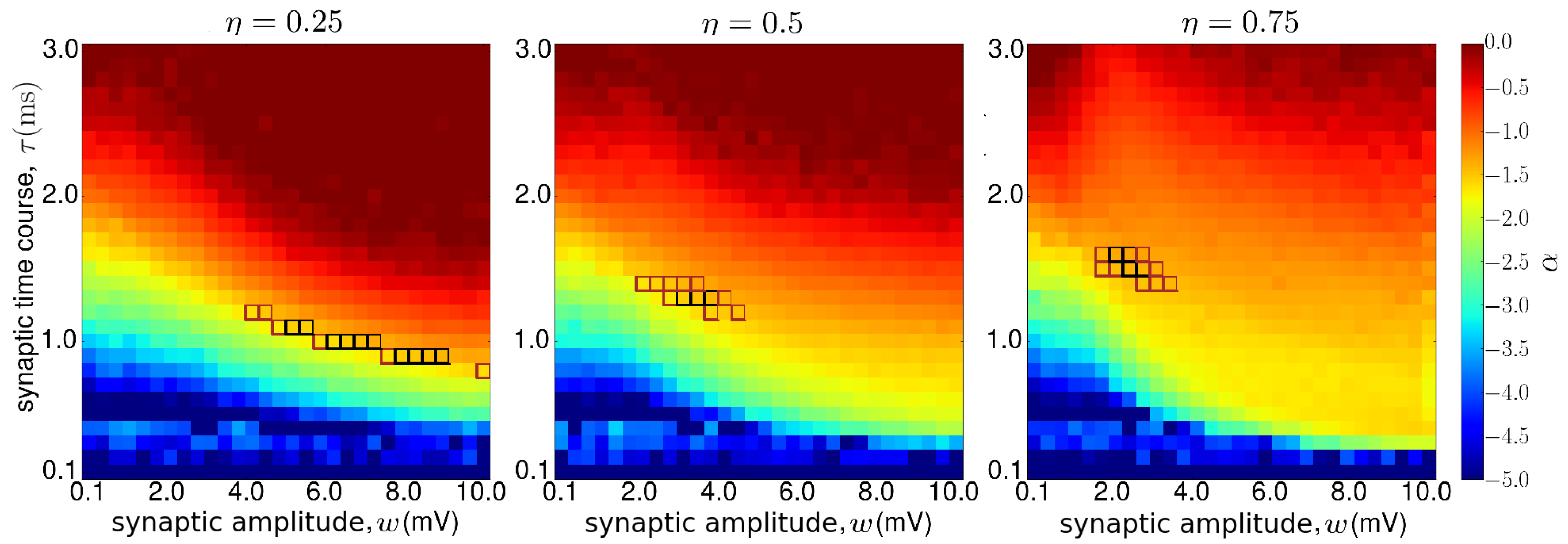
3.2. Metabolic Cost of Dynamic Regimes
3.3. Drosophila Network
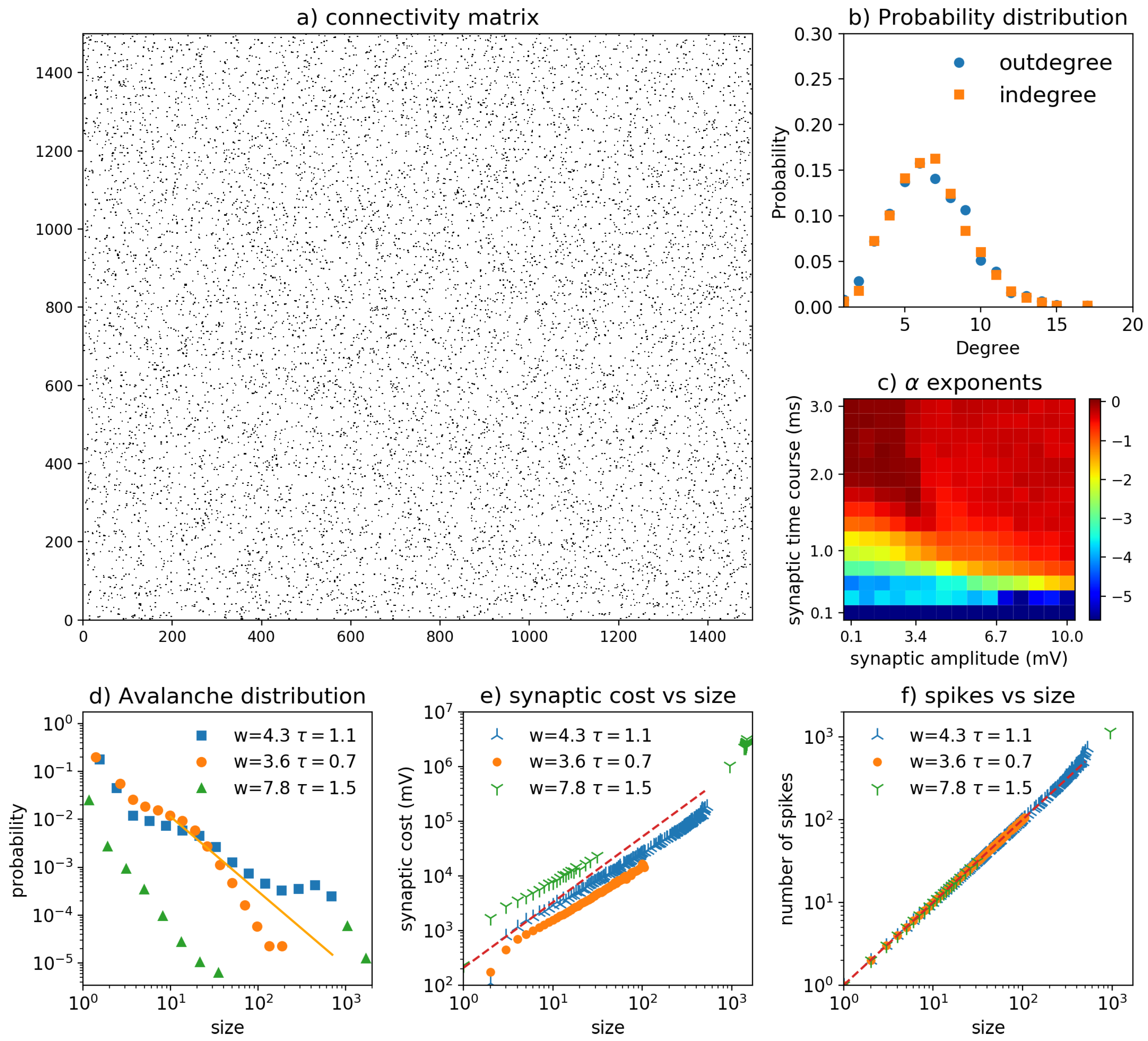
3.4. Erdos–Renyi Network
3.5. Disconnected Network
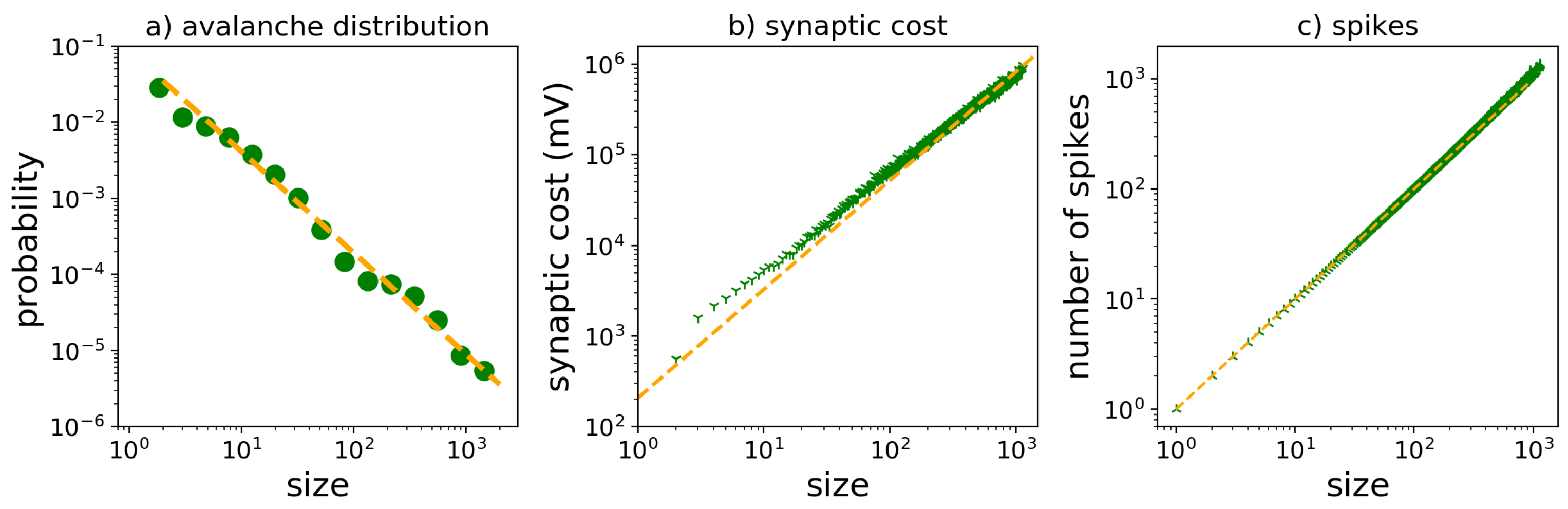
3.6. Critical Neural Networks Minimize Metabolic Cost
4. Discussion
Code Availability
Funding
Acknowledgments
Conflicts of Interest
References
- Chialvo, D.R. Emergent complex neural dynamics. Nat. Phys. 2010, 6, 744. [Google Scholar] [CrossRef] [Green Version]
- Bak, P. How Nature Works: The Science of Self-Organized Criticality; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Kinouchi, O.; Copelli, M. Optimal dynamical range of excitable networks at criticality. Nat. Phys. 2006, 2, 348. [Google Scholar] [CrossRef] [Green Version]
- Shew, W.L.; Yang, H.; Petermann, T.; Roy, R.; Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 2009, 29, 15595–15600. [Google Scholar] [CrossRef]
- Mandelbrot, B. Multifractals and 1/f Noise: Wild Self-Affinity in Physics; Springer: New York, NY, USA, 1998. [Google Scholar]
- Bak, P.; Tang, C.; Wiesenfeld, K. Self-organized criticality. Phys. Rev. A 1988, 38, 364. [Google Scholar] [CrossRef]
- Linkenkaer-Hansen, K.; Nikouline, V.V.; Palva, J.M.; Ilmoniemi, R.J. Long-range temporal correlations and scaling behavior in human brain oscillations. J. Neurosci. 2001, 21, 1370–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, H.E. Phase Transitions and Critical Phenomena; Clarendon Press: Oxford, UK, 1971. [Google Scholar]
- Beggs, J.M.; Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 2003, 23, 11167–11177. [Google Scholar] [CrossRef] [Green Version]
- Massobrio, P.; Pasquale, V.; Martinoia, S. Self-organized criticality in cortical assemblies occurs in concurrent scale-free and small-world networks. Sci. Rep. 2015, 5, 10578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontenele, A.J.; de Vasconcelos, N.A.; Feliciano, T.; Aguiar, L.A.; Soares-Cunha, C.; Coimbra, B.; Dalla Porta, L.; Ribeiro, S.; Rodrigues, A.J.; Sousa, N.; et al. Criticality between cortical states. Phys. Rev. Lett. 2019, 122, 208101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitzbichler, M.G.; Smith, M.L.; Christensen, S.R.; Bullmore, E. Broadband criticality of human brain network synchronization. PLoS Comput. Biol. 2009, 5, e1000314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephen, D.G.; Stepp, N.; Dixon, J.A.; Turvey, M. Strong anticipation: Sensitivity to long-range correlations in synchronization behavior. Phys. A Stat. Mech. Its Appl. 2008, 387, 5271–5278. [Google Scholar] [CrossRef]
- Aguilar-Velázquez, D.; Guzmán-Vargas, L. Synchronization and 1/f signals in interacting small-world networks. Chaos Solitons Fractals 2017, 104, 418–425. [Google Scholar] [CrossRef]
- Aguilar-Velázquez, D.; Guzmán-Vargas, L. Critical synchronization and 1/f noise in inhibitory/excitatory rich-club neural networks. Sci. Rep. 2019, 9, 1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, J.; Grilli, J.; Suweis, S.; Muñoz, M.A.; Banavar, J.R.; Maritan, A. Information-based fitness and the emergence of criticality in living systems. Proc. Natl. Acad. Sci. USA 2014, 111, 10095–10100. [Google Scholar] [CrossRef] [Green Version]
- Sporns, O. Networks of the Brain; MIT Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Hasenstaub, A.; Otte, S.; Callaway, E.; Sejnowski, T.J. Metabolic cost as a unifying principle governing neuronal biophysics. Proc. Natl. Acad. Sci. USA 2010, 107, 12329–12334. [Google Scholar] [CrossRef] [Green Version]
- Watts, D.J.; Strogatz, S.H. Collective dynamics of ‘small-world’ networks. Nature 1998, 393, 440. [Google Scholar] [CrossRef]
- Bullmore, E.; Sporns, O. The economy of brain network organization. Nat. Rev. Neurosci. 2012, 13, 336. [Google Scholar] [CrossRef] [PubMed]
- Eguíluz, V.M.; Chialvo, D.R.; Cecchi, G.A.; Baliki, M.; Apkarian, A.V. Scale-free brain functional networks. Phys. Rev. Lett. 2005, 94, 018102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Den Heuvel, M.P.; Sporns, O. Rich-club organization of the human connectome. J. Neurosci. 2011, 31, 15775–15786. [Google Scholar] [CrossRef]
- Nigam, S.; Shimono, M.; Ito, S.; Yeh, F.C.; Timme, N.; Myroshnychenko, M.; Lapish, C.C.; Tosi, Z.; Hottowy, P.; Smith, W.C.; et al. Rich-club organization in effective connectivity among cortical neurons. J. Neurosci. 2016, 36, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Mondragón, R.J. The rich-club phenomenon in the Internet topology. IEEE Commun. Lett. 2004, 8, 180–182. [Google Scholar] [CrossRef] [Green Version]
- Senden, M.; Deco, G.; De Reus, M.A.; Goebel, R.; Van Den Heuvel, M.P. Rich club organization supports a diverse set of functional network configurations. NeuroImage 2014, 96, 174–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonifazi, P.; Goldin, M.; Picardo, M.A.; Jorquera, I.; Cattani, A.; Bianconi, G.; Represa, A.; Ben-Ari, Y.; Cossart, R. GABAergic hub neurons orchestrate synchrony in developing hippocampal networks. Science 2009, 326, 1419–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barabási, A.L.; Albert, R. Emergence of scaling in random networks. Science 1999, 286, 509–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravasz, E.; Barabási, A.L. Hierarchical organization in complex networks. Phys. Rev. E 2003, 67, 026112. [Google Scholar] [CrossRef] [Green Version]
- Kandel, E.R. In Search of Memory: The Emergence of a New Science of Mind; WW Norton & Company: New York, NY, USA, 2007. [Google Scholar]
- Attwell, D.; Laughlin, S.B. An energy budget for signaling in the grey matter of the brain. J. Cereb. Blood Flow Metab. 2001, 21, 1133–1145. [Google Scholar] [CrossRef]
- Buzsáki, G.; Kaila, K.; Raichle, M. Inhibition and brain work. Neuron 2007, 56, 771–783. [Google Scholar] [CrossRef] [Green Version]
- Izhikevich, E.M. Simple model of spiking neurons. IEEE Trans. Neural Netw. 2003, 14, 1569–1572. [Google Scholar] [CrossRef] [Green Version]
- Hansel, D.; Mato, G.; Meunier, C.; Neltner, L. On numerical simulations of integrate-and-fire neural networks. Neural Comput. 1998, 10, 467–483. [Google Scholar] [CrossRef]
- Pauli, R.; Weidel, P.; Kunkel, S.; Morrison, A. Reproducing polychronization: A guide to maximizing the reproducibility of spiking network models. Front. Neuroinform. 2018, 12, 46. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Sjöström, P.J.; Reigl, M.; Nelson, S.; Chklovskii, D.B. Highly nonrandom features of synaptic connectivity in local cortical circuits. PLoS Biol. 2005, 3, e68. [Google Scholar]
- Bruns, D.; Jahn, R. Real-time measurement of transmitter release from single synaptic vesicles. Nature 1995, 377, 62. [Google Scholar] [CrossRef]
- Colizza, V.; Flammini, A.; Serrano, M.A.; Vespignani, A. Detecting rich-club ordering in complex networks. Nat. Phys. 2006, 2, 110. [Google Scholar] [CrossRef]
- Beggs, J.M.; Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J. Neurosci. 2004, 24, 5216–5229. [Google Scholar] [CrossRef] [Green Version]
- Zapperi, S.; Lauritsen, K.B.; Stanley, H.E. Self-organized branching processes: Mean-field theory for avalanches. Phys. Rev. Lett. 1995, 75, 4071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollander, M.; Wolfe, D.A. Nonparametric Statistical Methods; Wiley: New York, NY, USA, 1973. [Google Scholar]
- Clauset, A.; Shalizi, C.R.; Newman, M.E. Power-law distributions in empirical data. SIAM Rev. 2009, 51, 661–703. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, D.J.; Berger, J.O.; Johannesson, M.; Nosek, B.A.; Wagenmakers, E.J.; Berk, R.; Bollen, K.A.; Brembs, B.; Brown, L.; Camerer, C.; et al. Redefine statistical significance. Nat. Hum. Behav. 2018, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Sornette, D. Critical Phenomena in Natural Sciences: Chaos, Fractals, Selforganization and Disorder: Concepts and Tools; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Lennie, P. The cost of cortical computation. Curr. Biol. 2003, 13, 493–497. [Google Scholar] [CrossRef] [Green Version]
- Rossi, R.; Ahmed, N. The network data repository with interactive graph analytics and visualization. In Proceedings of the Twenty-Ninth AAAI Conference on Artificial Intelligence, Austin, TX, USA, 25–30 January 2015. [Google Scholar]
- Shih, C.T.; Sporns, O.; Yuan, S.L.; Su, T.S.; Lin, Y.J.; Chuang, C.C.; Wang, T.Y.; Lo, C.C.; Greenspan, R.J.; Chiang, A.S. Connectomics-based analysis of information flow in the Drosophila brain. Curr. Biol. 2015, 25, 1249–1258. [Google Scholar] [CrossRef] [Green Version]
- Erdös, P.; Rényi, A. On random graphs. Publ. Math. 1959, 6, 290–297. [Google Scholar]
- Kim, J.Z.; Soffer, J.M.; Kahn, A.E.; Vettel, J.M.; Pasqualetti, F.; Bassett, D.S. Role of graph architecture in controlling dynamical networks with applications to neural systems. Nat. Phys. 2018, 14, 91. [Google Scholar] [CrossRef]
- Aguilar-Velázquez, D.; Reyes-Ramírez, I. A wavelet analysis of multiday extreme ozone and its precursors in Mexico city during 2015–2016. Atmos. Environ. 2018, 188, 112–119. [Google Scholar] [CrossRef]
- Towlson, E.K.; Vértes, P.E.; Ahnert, S.E.; Schafer, W.R.; Bullmore, E.T. The rich club of the C. elegans neuronal connectome. J. Neurosci. 2013, 33, 6380–6387. [Google Scholar] [CrossRef] [Green Version]
- Faber, S.P.; Timme, N.M.; Beggs, J.M.; Newman, E.L. Computation is concentrated in rich clubs of local cortical networks. Netw. Neurosci. 2019, 3, 384–404. [Google Scholar] [CrossRef]
- Izhikevich, E.M. Which model to use for cortical spiking neurons? IEEE Trans. Neural Netw. 2004, 15, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Buckner, R.L.; Sepulcre, J.; Talukdar, T.; Krienen, F.M.; Liu, H.; Hedden, T.; Andrews-Hanna, J.R.; Sperling, R.A.; Johnson, K.A. Cortical hubs revealed by intrinsic functional connectivity: Mapping, assessment of stability, and relation to Alzheimer’s disease. J. Neurosci. 2009, 29, 1860–1873. [Google Scholar] [CrossRef] [Green Version]
- van den Heuvel, M.P.; Sporns, O. Network hubs in the human brain. Trends Cognit. Sci. 2013, 17, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Friston, K. The free-energy principle: A unified brain theory? Nat. Rev. Neurosci. 2010, 11, 127. [Google Scholar] [CrossRef]
- Mangalam, M.; Carver, N.S.; Kelty-Stephen, D.G. Global broadcasting of local fractal fluctuations in a bodywide distributed system supports perception via effortful touch. Chaos Solitons Fractals 2020, 135, 109740. [Google Scholar] [CrossRef]
- Yu, L.; Yu, Y. Energy-efficient neural information processing in individual neurons and neuronal networks. J. Neurosci. Res. 2017, 95, 2253–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poil, S.S.; Hardstone, R.; Mansvelder, H.D.; Linkenkaer-Hansen, K. Critical-state dynamics of avalanches and oscillations jointly emerge from balanced excitation/inhibition in neuronal networks. J. Neurosci. 2012, 32, 9817–9823. [Google Scholar] [CrossRef]
- Socolar, J.E.; Kauffman, S.A. Scaling in ordered and critical random Boolean networks. Phys. Rev. Lett. 2003, 90, 068702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haldeman, C.; Beggs, J.M. Critical branching captures activity in living neural networks and maximizes the number of metastable states. Phys. Rev. Lett. 2005, 94, 058101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favela, L.H. Radical embodied cognitive neuroscience: Addressing “grand challenges” of the mind sciences. Front. Hum. Neurosci. 2014, 8, 796. [Google Scholar] [CrossRef] [Green Version]
- Aldana, M.; Balleza, E.; Kauffman, S.; Resendiz, O. Robustness and evolvability in genetic regulatory networks. J. Theor. Biol. 2007, 245, 433–448. [Google Scholar] [CrossRef] [PubMed]
- Bertschinger, N.; Natschläger, T. Real-time computation at the edge of chaos in recurrent neural networks. Neural Comput. 2004, 16, 1413–1436. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilar-Velázquez, D. Critical Neural Networks Minimize Metabolic Cost. Physics 2021, 3, 42-58. https://doi.org/10.3390/physics3010005
Aguilar-Velázquez D. Critical Neural Networks Minimize Metabolic Cost. Physics. 2021; 3(1):42-58. https://doi.org/10.3390/physics3010005
Chicago/Turabian StyleAguilar-Velázquez, Daniel. 2021. "Critical Neural Networks Minimize Metabolic Cost" Physics 3, no. 1: 42-58. https://doi.org/10.3390/physics3010005
APA StyleAguilar-Velázquez, D. (2021). Critical Neural Networks Minimize Metabolic Cost. Physics, 3(1), 42-58. https://doi.org/10.3390/physics3010005