
Deciphering Molecular Dynamics of Foot and Mouth Disease Virus (FMDV): A Looming Threat to Pakistan’s Dairy Industry

Abstract
:1. Introduction
2. Etiology
3. FMDV Serotypes
4. Insights into Genetic/Molecular Dynamics of FMD
5. Geographical Distribution of FMD
5.1. FMD in South Asia
5.2. The Situation of FMD in Pakistan
6. Phylogenetic Analysis of FMDV Serotypes
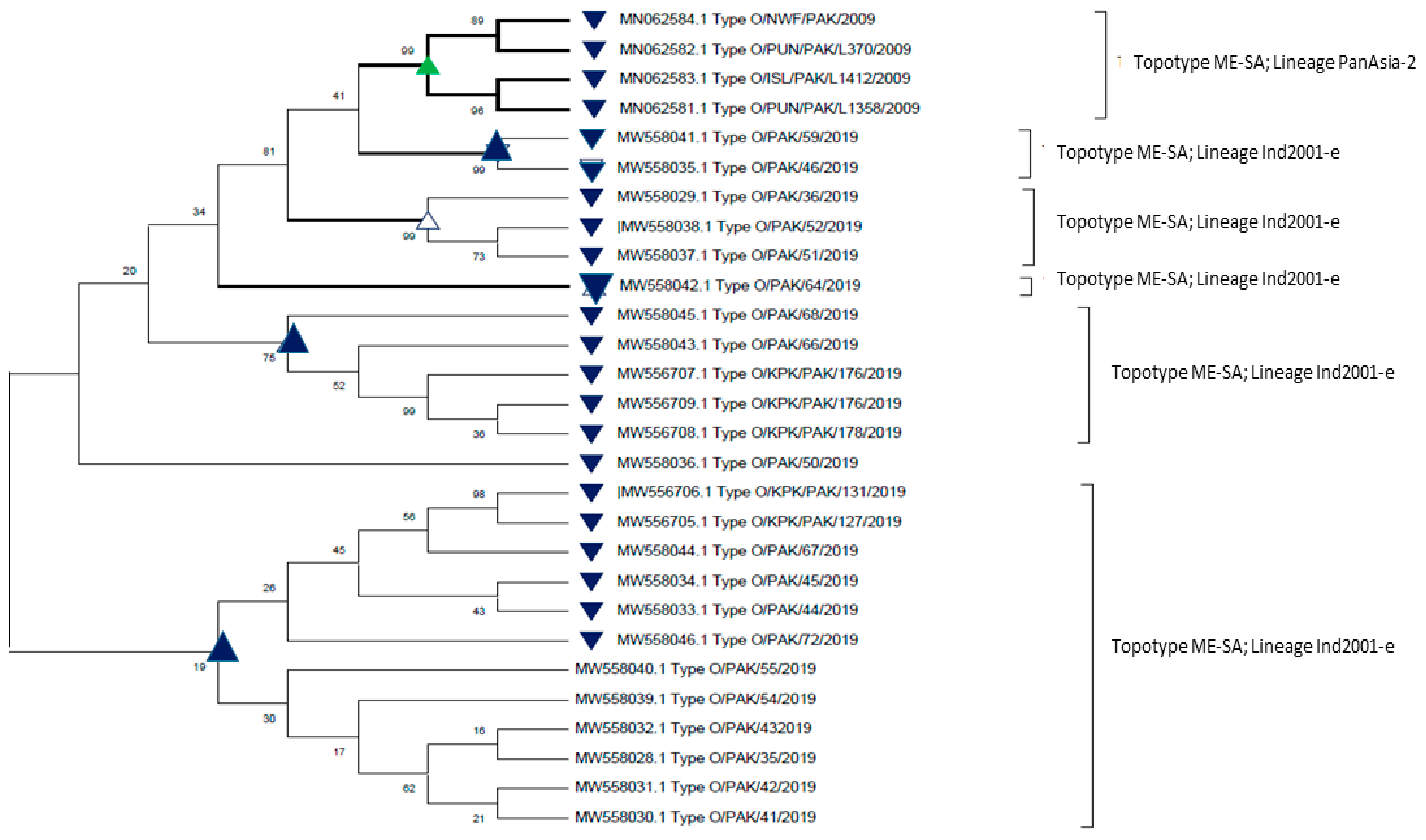
6.1. Serotype O
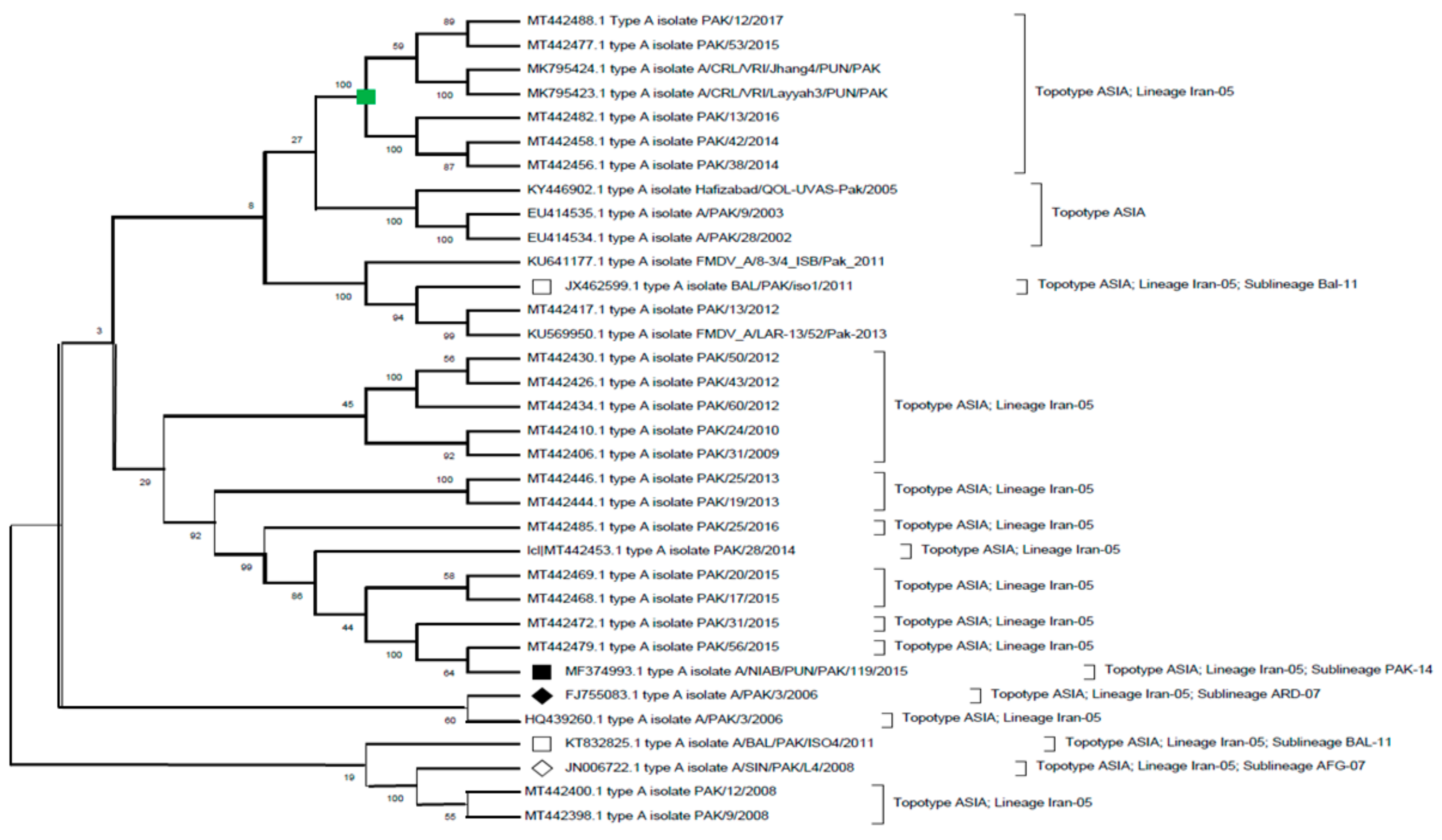
6.2. Serotype A
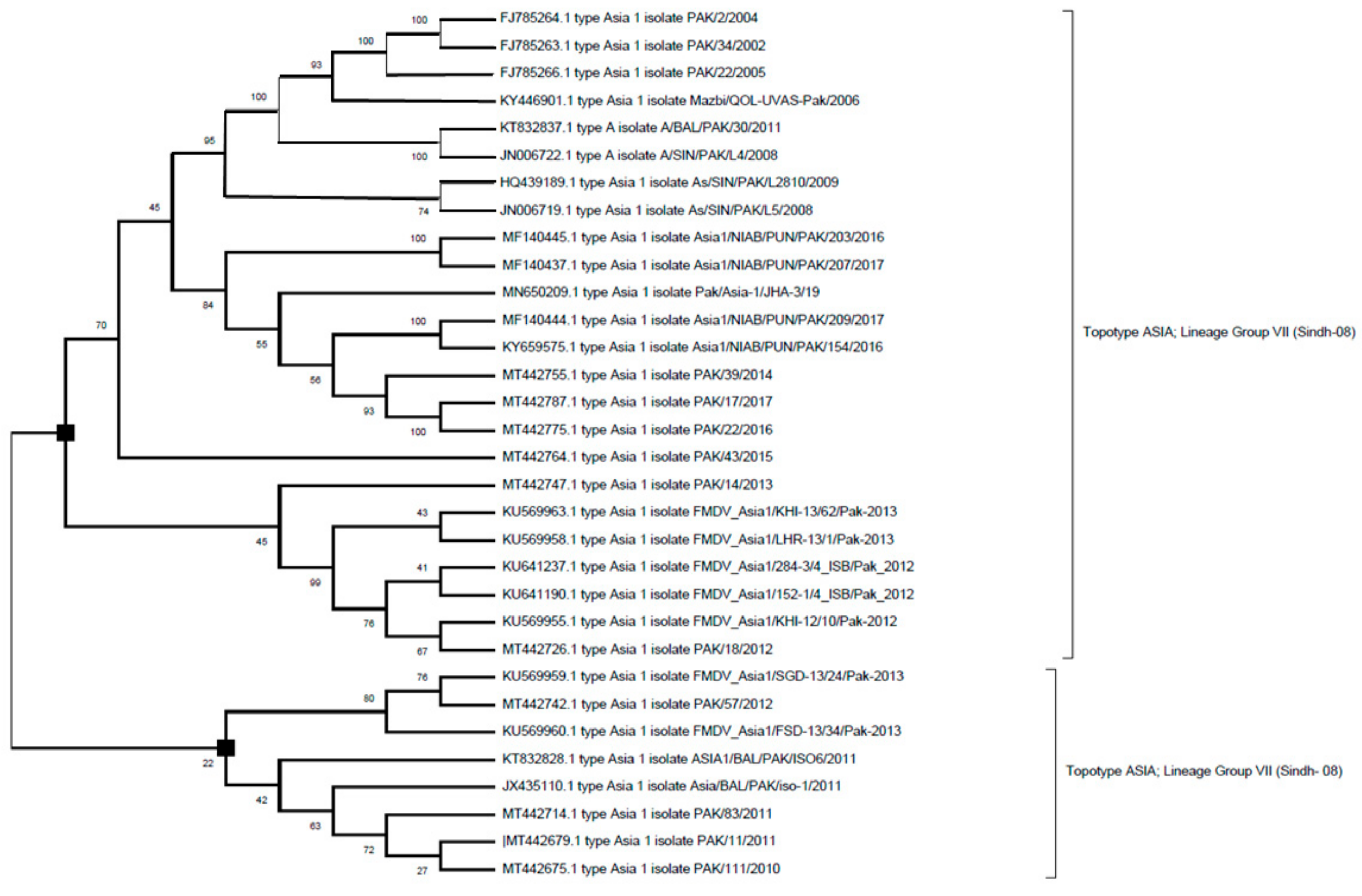
6.3. Serotype Asia-1
7. Sequence Analysis of FMDV Strains from Pakistan



8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References and Notes
- Alhaji, N.; Amin, J.; Aliyu, M.; Mohammad, B.; Babalobi, O.; Wungak, Y.; Odetokun, I. Economic impact assessment of foot-and-mouth disease burden and control in pastoral local dairy cattle production systems in Northern Nigeria: A cross-sectional survey. Prev. Vet. Med. 2020, 177, 104974. [Google Scholar] [CrossRef]
- Bulletins Archives—IDF—IDF Is the Leading Source of Scientific and Technical Expertise for All Stakeholders of the Dairy Chain. Available online: https://fil-idf.org/product-category/bulletin/ (accessed on 23 December 2021).
- Pakistan Finance Division. Pakistan Economic Survey 2018–2019; 2019. Available online: http://www.finance.gov.pk/survey/chapters_19/Economic_Survey_2018_19.pdf (accessed on 23 December 2021).
- Siddiky, M. Dairying in South Asian region: Opportunities, challenges and way forward. SAARC J. Agric. 2017, 15, 173–187. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J. Milk: The Mammary Gland and Its Secretion; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016; p. 89. [Google Scholar]
- Rushton, J. The Economics of Animal Health and Production; CABI: Wallingford, UK, 2009. [Google Scholar]
- Awel, S.; Dilba, G.; Abraha, B.; Zewde, D.; Wakjira, B.; Aliy, A. Seroprevalence and molecular detection of foot and mouth disease virus in dairy cattle around Addis Ababa, Central Ethiopia. Vet. Med. Res. Rep. 2021, 12, 187–197. [Google Scholar] [CrossRef]
- Lee, M.; Jo, H.; Park, S.; Ko, M.; Kim, S.; Kim, B.; Park, J. Advanced Foot-and-mouth disease vaccine platform for stimulation of simultaneous cellular and humoral immune responses. Vaccines 2020, 8, 254. [Google Scholar] [CrossRef]
- Azeem, A. A review on foot and mouth disease in dairy animals, etiology, pathogenesis and clinical findings. Pure Appl. Biol. 2020, 9, 821–832. [Google Scholar] [CrossRef]
- Upadhayay, U.; Ewam, P. Epidemiology and diagnosis of foot-and-mouth disease: A review. Indian J. Anim. Sci. 2012, 6, 543–551. [Google Scholar]
- Hietela, S.; Ardans, A. Molecular weapons against agricultural vulnerability and the war on terror. J. Vet. Med. Educ. 2003, 30, 155–156. [Google Scholar] [CrossRef] [PubMed]
- Samina, I. Host factors affecting the homologous and heterologous immune response of cattle to FMDV: Genetic background, age, virus strains and route of administration. Vaccines 1998, 16, 335–339. [Google Scholar] [CrossRef]
- Vallée, H.; Carré, H. Sur la pluralité du virus aphteux. CR Acad. Sci. Paris 1922, 174, 1498–1500. [Google Scholar]
- Abubakar, M. An evaluation of foot-and-mouth disease outbreak in relation to vaccination in a herd of cattle and buffaloes. Res. J. Vet. Pract. 2014, 2, 28–29. [Google Scholar] [CrossRef] [Green Version]
- Knight-Jones, T.J.; Rushton, J. The economic impacts of foot and mouth disease–What are they, how big are they and where do they occur? Prev. Vet. Med. 2013, 112, 161–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abubakar, M.; Khan, E.; Arshed, M.; Gonzales, J.; Ferrari, G.; Hussain, M.; Ali, Q. An appraisal on the occurrence of foot-and-mouth disease virus serotypes in cattle and buffaloes, Pakistan. Arch. Virol. 2015, 160, 1561–1564. [Google Scholar] [CrossRef]
- Feasibility Analysis for Cluster Development Based Agriculture Transformation. Cluster Development Based Agriculture Transformation Plan Vision-2025; Ministry of Planning Development and Special Initiatives: Islamabad, Pakistan, 2020.
- Brito, B.; Rodriguez, L.; Hammond, J.; Pinto, J.; Perez, A. Review of the global distribution of foot-and-mouth disease virus from 2007 to 2014. Transbound. Emerg. Dis. 2015, 64, 316–332. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, M. The causative agent of FMD. In Some RNA Viruses; IntechOpen Limited: London, UK, 2021; p. 79. [Google Scholar]
- Chakraborty, S. Foot-and-mouth disease, an economically important disease of animals. Adv. Anim. Vet. Sci. 2014, 2, 1–18. [Google Scholar] [CrossRef]
- Morelli, M.; Wright, C.; Knowles, N.; Juleff, N.; Paton, D.; King, D.; Haydon, D. Evolution of foot-and-mouth disease virus intra-sample sequence diversity during serial transmission in bovine hosts. Vet. Res. 2013, 44, 12. [Google Scholar] [CrossRef] [Green Version]
- Paton, D.J.; Di Nardo, A.; Knowles, N.J.; Wadsworth, J.; Pituco, E.M.; Cosivi, O.; Rivera, A.M.; Kassimi, L.B.; Brocchi, E.; de Clercq, K.; et al. The history of foot-and-mouth disease virus serotype C: The first known extinct serotype? Virus Evol. 2021, 7, veab009. [Google Scholar] [CrossRef]
- Rudreshappa, A.; Sanyal, A.; Mohapatra, J.; Subramaniam, S.; De, A.; Das, B.; Singanallur, N.; Jangam, A.; Muthukrishnan, M.; Villuppanoor, S.; et al. Emergence of antigenic variants with in serotype A foot and mouth disease virus in India and evaluation of a new vaccine candidate panel. Vet. Microbiol. 2012, 158, 405–409. [Google Scholar] [CrossRef]
- Ding, Y.; Chen, H.; Zhang, J.; Zhou, J.; Ma, L.; Zhang, L.; Gu, Y.; Liu, Y. An overview of control strategy and diagnostic technology for foot-and-mouth disease in China. Virol. J. 2013, 10, 78. [Google Scholar] [CrossRef] [Green Version]
- Doel, T. FMD Vaccines. Virus Res. 2003, 91, 81–99. [Google Scholar] [CrossRef]
- Knowles, N.; Samuel, A. Molecular epidemiology of foot-and-mouth disease virus. Virus Res. 2003, 91, 65–80. [Google Scholar] [CrossRef]
- Yang, M.; Holland, H.; Clavijo, A. Production of monoclonal antibodies against whole virus particles of foot-and-mouth disease virus serotype O and A and their potential use in quantification of intact virus for vaccine manufacture. Vaccines 2008, 26, 3377–3382. [Google Scholar] [CrossRef] [PubMed]
- Knight-Jones, T.; McLaws, M.; Rushton, J. Foot-and-mouth disease impact on smallholders—What do we know, what don’t we know and how can we find out more? Transbound. Emerg. Dis. 2016, 64, 1079–1094. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, M.; Parida, S. Foot and mouth disease vaccine strain selection: Current approaches and future perspectives. Expert Rev. Vaccines 2018, 17, 577–591. [Google Scholar] [CrossRef] [PubMed]
- WRLFMD. World Reference Laboratory for Foot and Mouth; OIE/FAO Reference Laboratory Contract Report; Annual Report; The Pirbright Institute: Pirbright, UK, 2016. [Google Scholar]
- Subramaniam, S.; Mohapatra, J.; Das, B.; Sanyal, A.; Pattnaik, B. Genetic and antigenic analysis of foot-and-mouth disease virus serotype O responsible for outbreaks in India during 2013. Infect. Genet. Evol. 2015, 30, 59–64. [Google Scholar] [CrossRef]
- Jamal, S.; Ferrari, G.; Ahmed, S.; Normann, P.; Belsham, G. Genetic diversity of foot-and-mouth disease virus serotype O in Pakistan and Afghanistan, 1997–2009. Infect. Genet. Evol. 2011, 11, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Namatovu, A.; Belsham, G.; Ayebazibwe, C.; Dhikusooka, M.; Wekesa, S.; Siegismund, H.; Muwanika, V.; Tjørnehøj, K. Challenges for serology-based characterization of foot-and-mouth disease outbreaks in endemic areas; identification of two separate lineages of serotype O FMDV in Uganda In 2011. Transbound. Emerg. Dis. 2013, 62, 522–534. [Google Scholar] [CrossRef] [Green Version]
- World Animal Health Information Database (WAHID)—Web Resources. Available online: https://webresources.articles411.com/world-animal-health-information-database-wahid (accessed on 23 December 2021).
- Forman, S.; Le Gall, F.; Belton, D.; Evans, B.; François, J.L.; Murray, G.; Sheesley, D.; Vandersmissen, A.; Yoshimura, S. Moving towards the global control of foot and mouth disease: An opportunity for donors. Rev. Sci. Tech. 2009, 28, 883. [Google Scholar] [CrossRef]
- Kumar, R.; Sreenivasa, B.; Tamilselvan, R. Construction and characterization of recombinant human adenovirus type 5 expressing foot-and-mouth disease virus capsid proteins of indian vaccine strain, O/IND/R2/75. Vet. World 2015, 8, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Ashkani, J.; Rees, D. The critical role of vp1 in forming the necessary cavities for receptor-mediated entry of FMDV to the host cell. Sci. Rep. 2016, 6, 27140. [Google Scholar] [CrossRef] [Green Version]
- Garcıía-Briones, M.; Blanco, E.; Chiva, C.; Andreu, D.; Ley, V.; Sobrino, F. immunogenicity and T cell recognition in swine of foot-and-mouth disease virus polymerase 3D. Virology 2004, 322, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Singh, I.; Deb, R.; Kumar, S.; Singh, R.; Andonissamy, J.; Smita, S.; Sengar, G.; Kumar, R.; Ojha, K.; Sahoo, N.; et al. Deciphering foot-and-mouth disease (FMD) virus–host tropism. J. Biomol. Struct. Dyn. 2019, 37, 4779–4789. [Google Scholar] [CrossRef]
- Domingo, E.; Baranowski, E.; Escarmís, C.; Sobrino, F. Foot-and-mouth disease virus. Comp. Immunol. Microbiol. Infect. Dis. 2002, 25, 297–308. [Google Scholar] [CrossRef]
- Diaz-San Segundo, F.; Medina, G.; Stenfeldt, C.; Arzt, J.; de los Santos, T. Foot-and-mouth disease vaccines. Vet. Microbiol. 2017, 206, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.; Barnett, P. Experimental evaluation of Foot-and-mouth disease vaccines for emergency use in ruminants and pigs: A review. Vet. Res. 2008, 40, 13. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Bai, X.; Li, P.; Zhang, M.; Bao, H.; Sun, P.; Lu, Z.; Cao, Y.; Chen, Y.; Li, D.; et al. Influence of foot-and-mouth disease virus O/CHN/Mya98/33-P strain leader protein on viral replication and host innate immunity. Viral Immunol. 2015, 28, 360–366. [Google Scholar] [CrossRef]
- Jackson, T.; Sharma, A.; Ghazaleh, R.; Blakemore, W.; Ellard, F.; Simmons, D.; Newman, J.; Stuart, D.; King, A. Arginine-Glycine-Aspartic Acid-specific binding by foot-and-mouth disease viruses to the purified integrin Alpha(V)Beta3 in vitro. J. Virol. 1997, 71, 8357–8361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, I.; Malirat, V.; Falczuk, A. Evolving Perception On The Benefits Of Vaccination As A Foot And Mouth Disease Control Policy: Contributions Of South America. Expert Rev. Vaccines 2005, 4, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Pinto, J. Foot-and-Mouth Disease Situation Food and Agriculture Organization of the United Nations Monthly Report; FAO: Roma, Italy, 2017; pp. 21–25. [Google Scholar]
- Shmeiger, Z.; Miculitzki, M.; Gelman, B.; Vaxman, I.; Goshen, T. The effect of foot and mouth disease morbidity influencing periparturient diseases and culling on Nir Yitzhak dairy cattle farm. Isr. J. Vet. Med. 2021, 76, 27. [Google Scholar]
- Gizaw, D.; Tesfaye, Y.; Wood, B.; Di Nardo, A.; Shegu, D.; Muluneh, A.; Bilata, T.; Belayneh, R.; Fentie, A.; Asgdome, H.; et al. Molecular characterization of foot-and-mouth disease viruses circulating in Ethiopia between 2008 and 2019. Transbound. Emerg. Dis. 2020, 67, 2983–2992. [Google Scholar] [CrossRef]
- Lyons, N.; Alexander, N.; Stärk, K.; Dulu, T.; Sumption, K.; James, A.; Rushton, J.; Fine, P. Impact of foot-and-mouth disease on milk production on a large-scale dairy farm in Kenya. Prev. Vet. Med. 2015, 120, 177–186. [Google Scholar] [CrossRef]
- Lyons, N.; Stärk, K.; van Maanen, C.; Thomas, S.; Chepkwony, E.; Sangula, A.; Dulu, T.; Fine, P. Epidemiological analysis of an outbreak of foot-and-mouth disease (serotype SAT2) on a large dairy farm in Kenya using regular vaccination. Acta Tropica 2015, 143, 103–111. [Google Scholar] [CrossRef]
- Muroga, N.; Hayama, Y.; Yamamoto, T.; Kurogi, A.; Tsuda, T.; Tsutsui, T. The 2010 foot-and-mouth disease epidemic in Japan. J. Vet. Med. Sci. 2012, 74, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Haydon, D.; Kao, R.; Kitching, R. The UK foot-and-mouth disease outbreak—The aftermath. Nat. Rev. Microbiol. 2004, 2, 675–681. [Google Scholar] [CrossRef]
- O’Rourke, K.; Williamson, J. After columbus: Explaining europe’s overseas trade boom, 1500–1800. J. Econ. Hist. 2002, 62, 417–456. [Google Scholar] [CrossRef]
- Lombard, M.; Fussel, A. Antigen and vaccine banks: Technical requirements and the role of the european antigen bank in emergency foot and mouth disease vaccination. Revue Sci. et Tech. de l’OIE 2007, 26, 117–134. [Google Scholar] [CrossRef]
- Jamal, S.; Belsham, G. Foot-And-Mouth Disease: Past, present and future. Vet. Res. 2013, 44, 116. [Google Scholar] [CrossRef] [Green Version]
- Fodgeby, E. Review of Epizootology and Control of Foot-and-Mouth Disease in Europe from 1937 to 1961; European Commission for the Control of Foot-and-Mouth Disease, Food and Agriculture Organization of the United Nations: Rome, Italy, 1962; p. 108. [Google Scholar]
- Capstick, P.; Telling, R.; Chapman, W.; Stewart, D. Growth of a cloned strain of hamster kidney cells in suspended cultures and their susceptibility to the virus of foot-and-mouth disease. Nature 1962, 195, 1163–1164. [Google Scholar] [CrossRef] [PubMed]
- Bahnemann, H. Binary ethylenimine as an inactivant for foot-and-mouth disease virus and its application for vaccine production. Arch. Virol. 1975, 47, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Foot-and-Mouth Disease Situation Food and Agriculture Organization of the United Nations Monthly Report; FAO: Roma, Italy, 2019; p. 32.
- Bachanek-Bankowska, K.; Di Nardo, A.; Wadsworth, J.; Henry, E.K.; Parlak, Ü.; Timina, A.; Mischenko, A.; Qasim, I.A.; Abdollahi, D.; Sultana, M.; et al. Foot-and-Mouth Disease in the Middle East Caused by an A/ASIA/G-VII Virus Lineage, 2015–2016. Emerg. Infect. Dis. 2018, 24, 1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agriculture-Ministry of Finance—Government of Pakistan-pdf—Chapter 2 Agriculture Pakistan’s Agriculture Sector plays a Central Role in the Course Hero. Available online: https://www.coursehero.com/file/45332132/Agriculture-Ministry-of-Finance-Government-of-Pakistan-pdf/ (accessed on 23 December 2021).
- Blench, R.; Chapman, R.; Slaymaker, T. A Study of the Role of Livestock in Poverty Reduction Strategy Papers; Food and Agriculture Organization of the United Nation: Rome, Italy, 2003; pp. 1–57. [Google Scholar]
- Jamal, S.; Ahmed, S.; Hussain, M.; Ali, Q. Status of foot-and-mouth disease in Pakistan. Arch. Virol. 2010, 155, 1487–1491. [Google Scholar] [CrossRef]
- Final Evaluation of the Project, Development of a Framework for the Progressive Control of Foot and Mouth Disease in Pakistan; FAO: Rome, Italy, 2016. Available online: http://www.fao.org/evaluation/digest/evaluation-detail/es/c/416084/ (accessed on 23 December 2021).
- Bachanek-Bankowska, K.; Di Nardo, A.; Wadsworth, J.; King, D.; Knowles, N. A47 reconstructing the evolutionary history of pandemic foot-and-mouth disease viruses: The impact of recombination within the emerging O/ME-SA/Ind-2001 lineage. Virus Evol. 2019, 5, vez002-046. [Google Scholar] [CrossRef] [Green Version]
- Zulfiqar, M. Draft Report for Development of National Disease Control Policy for Foot and Mouth Disease in Pakistan under the FAO Project Support for Emergency Prevention and Control of Main Trans-Boundary Animal Diseases in Pakistan Rinderpest; FMD PPR: Draft report Pakistan. 2003; p. 90.
- Abdul-Hamid, N.; Hussein, N.; Wadsworth, J.; Radford, A.; Knowles, N.; King, D. Phylogeography of foot-and-mouth disease virus types O and A In Malaysia and surrounding countries. Infect. Genet. Evol. 2011, 11, 320–328. [Google Scholar] [CrossRef]
- Burman, A.; Clark, S.; Abrescia, N.; Fry, E.; Stuart, D.; Jackson, T. Specificity of the VP1 GH loop of foot-and-mouth disease virus for Alphav integrins. J. Virol. 2006, 80, 9798–9810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, A.; Khan, Q.; Waheed, U.; Arshad, M.; Asif, M.; Farooq, M. RT-pcr evaluation for identification and sequence analysis of foot-and-mouth disease serotype O from 2006 To 2007 In Punjab, Pakistan. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Knowles, N.; Samuel, A.; Davies, P.; Midgley, R.; Valarcher, J. Pandemic strain of foot-and-mouth disease virus serotype O. Emerg. Infect. Dis. 2005, 11, 1887–1893. [Google Scholar] [CrossRef] [PubMed]
- Hicks, H.; Wadsworth, J.; Azhar, M.; Afzal, M.; Manzoor, S.; Abubakar, M.; Khan, E.; King, D.; Knowles, N. Genome sequences of foot-and-mouth disease virus O/ME-SA/Ind-2001E strains isolated in Pakistan. Microbiol. Resour. Announc. 2020, 9, e00242-14. [Google Scholar] [CrossRef] [PubMed]
- Rashtibaf, M.; Sharifi, K.; Zibaee, S.; Dehghani, H. A survey on the frequency of foot-and-mouth disease virus carriers in cattle in North-East of Iran by RT-PCR: Implications for revising disease control strategy. Transbound. Emerg. Dis. 2012, 59, 482–489. [Google Scholar] [CrossRef]
- Klein, J.; Hussain, M.; Ahmad, M.; Normann, P.; Afzal, M.; Alexandersen, S. Genetic characterisation of the recent foot-and-mouth disease virus subtype A/IRN/2005. Virol. J. 2007, 4, 122. [Google Scholar] [CrossRef] [Green Version]
- Samuel, A.; Knowles, N. Foot-and-mouth disease type O viruses exhibit genetically and geographically distinct evolutionary lineages (topotypes). J. Gen. Virol. 2001, 82, 609–621. [Google Scholar] [CrossRef]
- Mohapatra, J.; Subramaniam, S.; Pandey, L.; Pawar, S.; De, A.; Das, B.; Sanyal, A.; Pattnaik, B. Phylogenetic structure of serotype A foot-and-mouth disease virus: Global diversity and the Indian perspective. J. Gen. Virol. 2011, 92, 873–879. [Google Scholar] [CrossRef]
- Jamal, S.; Belsham, G. Molecular epidemiology, evolution and phylogeny of foot-and-mouth disease virus. Infect. Genet. Evol. 2018, 59, 84–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, W.; Habib, M. Molecular Evaluation of Vp1 Coding Region of Foot and Mouth Disease Virus Circulating in Pakistan. Sarhad J. Agric. 2018, 34, 979–985. [Google Scholar] [CrossRef]
- Kitching, R. The role of the world reference laboratories for foot-and-mouth disease and for rinderpest. Ann. N. Y. Acad. Sci. 2006, 916, 139–146. [Google Scholar] [CrossRef]
- Brooksby, J.; Rogers, J. Methods Used in Typing the Virus of Foot-and-Mouth Disease at Pirbright, 1950–1955: Methods of Typing and Cultivation of Foot-and-Mouth Disease Virus; European Productivity Agency of the Organization of European Cooperation (OEEC), The Pirbright Institute: Pirbright, UK, 1957; pp. 31–34. [Google Scholar]
- Yasin, S.; Huq, M. Foot-and-Mouth Disease in Pakistan. Off. Int. Epizoot. 1960, 54, 378–388. [Google Scholar]
- Sheikh, S. An outbreak of malignant Foot-and-mouth disease at government Dajjal cattle breeding farm, Qadirabad, District Montgomery. Healer 1960, 5, 3–7. [Google Scholar]
- Ullah, A.; Jamal, S.; Romey, A.; Gorna, K.; Kakar, M.; Abbas, F.; Ahmad, J.; Zientara, S.; Bakkali Kassimi, L. Genetic characterization of serotypes A and Asia-1 Foot-and-mouth disease viruses in Balochistan, Pakistan, in 2011. Transbound. Emerg. Dis. 2016, 64, 1569–1578. [Google Scholar] [CrossRef]
- Jamal, S.; Ferrari, G.; Ahmed, S.; Normann, P.; Belsham, G. Molecular characterization of serotype Asia-1 foot-and-mouth disease viruses in Pakistan and Afghanistan; emergence of a new genetic group and evidence for a novel recombinant virus. Infect. Genet. Evol. 2011, 11, 2049–2062. [Google Scholar] [CrossRef]
- Mahapatra, M.; Yuvaraj, S.; Madhanmohan, M.; Subramaniam, S.; Pattnaik, B.; Paton, D.; Srinivasan, V.; Parida, S. Antigenic and genetic comparison of foot-and-mouth disease virus serotype O Indian vaccine strain, O/IND/R2/75 against currently circulating viruses. Vaccines 2015, 33, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Felsenstein, J. Phylogenies from restriction sites: A maximum-likelihood approach. Evolution 1992, 46, 159–173. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Virus Pool | Geographic Location | Prevalent Serotype | References |
|---|---|---|---|
| Pool 1 | South-East, Eastern Asia | A, O, Asia-1 | [30] |
| Pool 2 | Southern Asia | A, O, Asia-1 | [31] |
| Pool 3 | Middle East and Eurasia | A, O, Asia-1 | [32] |
| Pool 4 | Eastern Africa | O, A, SAT-1, SAT-2 | [33] |
| Pool 5 | Central and Western Africa | O, A, SAT-1, SAT-2 | [34] |
| Pool 6 | Southern Africa | SAT 1, 2 and 3 | [34] |
| Pool 7 | South America | FMD free | [35] |
| Virus Pool | Country | Serotypes | Topotypes | Lineages |
|---|---|---|---|---|
| 3 | Pakistan | O | Middle East–South Asia (ME–SA) | PanAsia-2 Ind-2001e |
| A | ASIA | Iran-05 | ||
| ASIA | ASIA | Sindh-08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abubakar, M.; Syed, Z.; Manzoor, S.; Arshed, M.J. Deciphering Molecular Dynamics of Foot and Mouth Disease Virus (FMDV): A Looming Threat to Pakistan’s Dairy Industry. Dairy 2022, 3, 123-136. https://doi.org/10.3390/dairy3010010
Abubakar M, Syed Z, Manzoor S, Arshed MJ. Deciphering Molecular Dynamics of Foot and Mouth Disease Virus (FMDV): A Looming Threat to Pakistan’s Dairy Industry. Dairy. 2022; 3(1):123-136. https://doi.org/10.3390/dairy3010010
Chicago/Turabian StyleAbubakar, Muhammad, Zainab Syed, Shumaila Manzoor, and Muhammad Javed Arshed. 2022. "Deciphering Molecular Dynamics of Foot and Mouth Disease Virus (FMDV): A Looming Threat to Pakistan’s Dairy Industry" Dairy 3, no. 1: 123-136. https://doi.org/10.3390/dairy3010010
APA StyleAbubakar, M., Syed, Z., Manzoor, S., & Arshed, M. J. (2022). Deciphering Molecular Dynamics of Foot and Mouth Disease Virus (FMDV): A Looming Threat to Pakistan’s Dairy Industry. Dairy, 3(1), 123-136. https://doi.org/10.3390/dairy3010010