Canopy Accumulation: Are Seagrass Meadows a Sink of Microplastics?

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
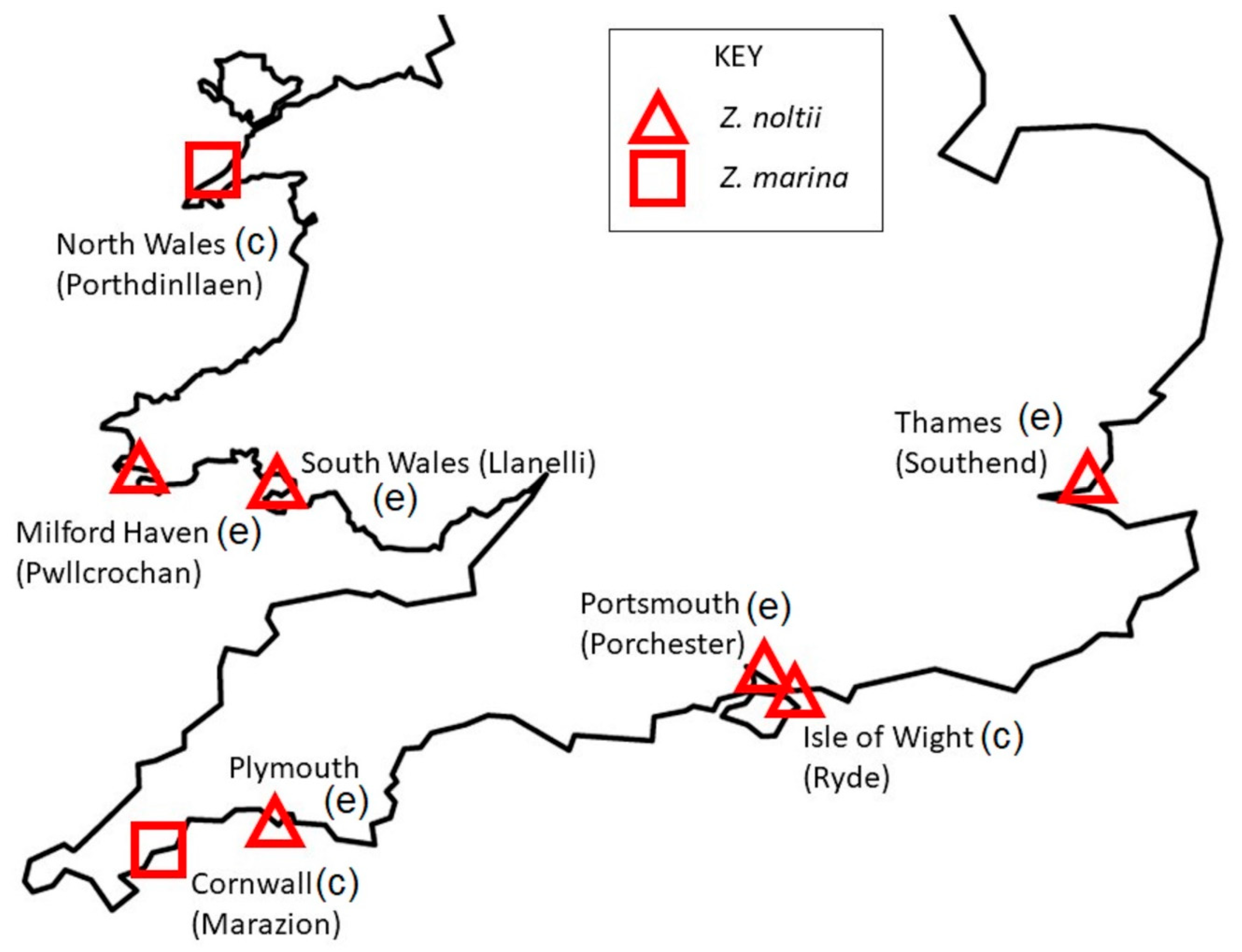
2.1. Sampling Locations
2.2. Sediment Sampling
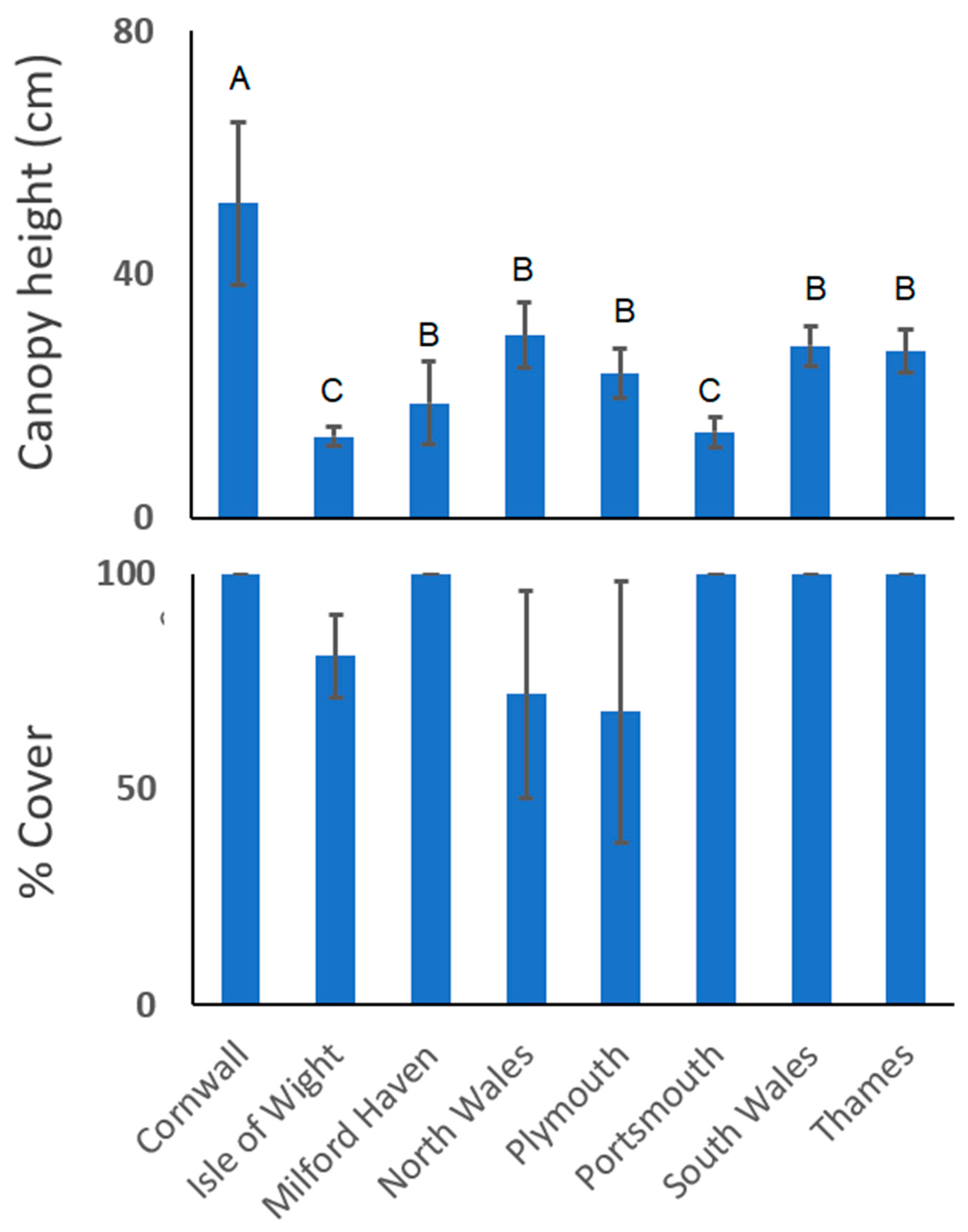
2.3. Seagrass Assessment
2.4. Preparation of Sample and Zinc Chloride Solution
2.5. Sediment Microplastic Isolation (SMI) Unit—Assembly and Purging
2.6. Extraction of Microplastic from Samples
2.7. Literature Review
2.8. Data Analysis
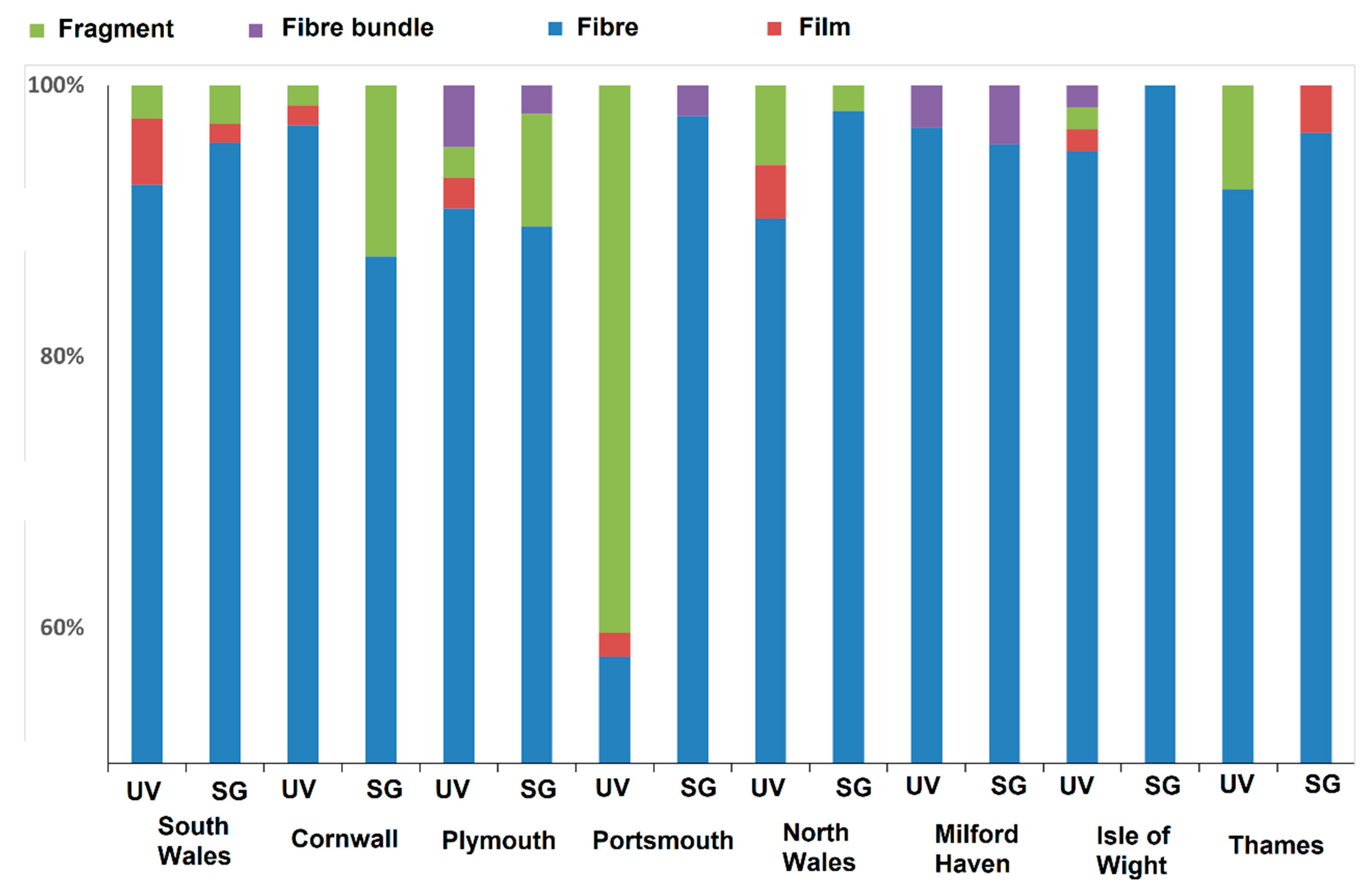
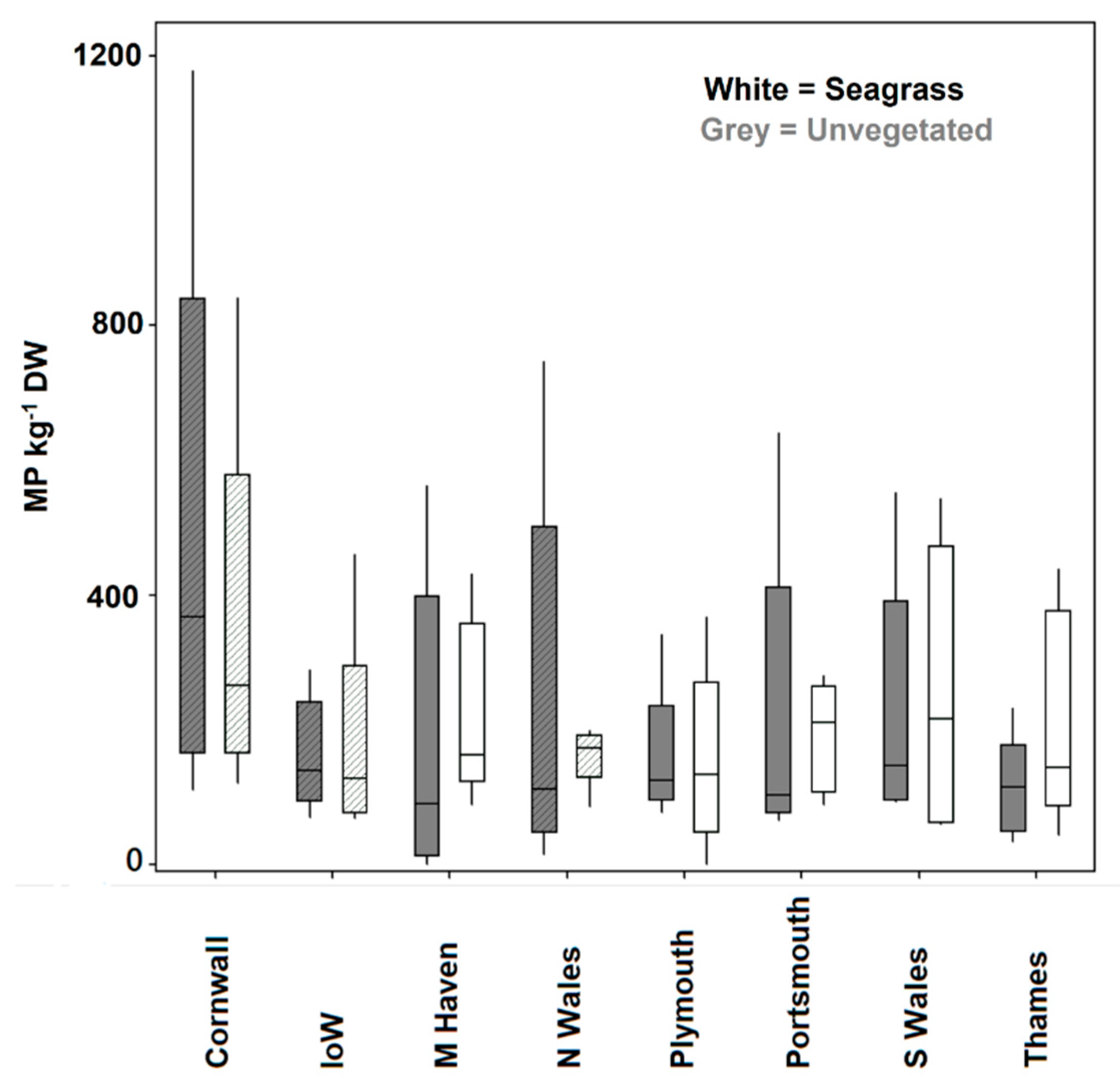
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nordlund, L.M.; Koch, E.W.; Barbier, E.B.; Creed, J.C. Seagrass ecosystem services and their variability across genera and geographical regions. PLoS ONE 2016, 11, e0163091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unsworth, R.K.F.; Nordlund, L.M.; Cullen-Unsworth, L.C. Seagrass meadows support global fisheries production. Conserv. Lett. 2019, 12, e12566. [Google Scholar] [CrossRef]
- Fonseca, M.S.; Fisher, J.S.; Zieman, J.C. Influence of the seagrass, IZostera marina L., on current flow. Est. Coast. Shelf Sci. 1982, 15, 351–364. [Google Scholar] [CrossRef]
- Maxwell, P.S.; Eklöf, J.S.; van Katwijk, M.M.; O’Brien, K.R.; de la Torre-Castro, M.; Boström, C.; Bouma, T.J.; Krause-Jensen, D.; Unsworth, R.K.F.; van Tussenbroek, B.I.; et al. The fundamental role of ecological feedback mechanisms for the adaptive management of seagrass ecosystems—A review. Biol. Rev. 2017, 92, 1521–1538. [Google Scholar] [CrossRef] [Green Version]
- Orth, R.J.; Carruthers, T.J.B.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L.; Randall Hughes, A.; Kendrick, G.A.; Judson Kenworthy, W.; Olyarnik, S.; et al. A global crisis for seagrass ecosystems. Bioscience 2006, 56, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.B.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [Green Version]
- Cozzolino, L.; Nicastro, K.R.; Zardi, G.I.; de los Santos, C.B. Species-specific plastic accumulation in the sediment and canopy of coastal vegetated habitats. Sci. Total Environ. 2020, 723, 138018. [Google Scholar] [CrossRef]
- Statista. Global Plastic Production. Available online: https://www.statista.com/statistics/282732/global-production-of-plastics-since-1950/ (accessed on 1 May 2019).
- Van Cauwenberghe, L.; Devriese, L.; Galgani, F.; Robbens, J.; Janssen, C.R. Microplastics in sediments: A review of techniques, occurrence and effects. Mar. Environ. Res. 2015, 111, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Browne, M.A.; Galloway, T.S.; Thompson, R.C. Spatial Patterns of Plastic Debris along Estuarine Shorelines. Environ. Sci. Technol. 2010, 44, 3404–3409. [Google Scholar] [CrossRef]
- Derraik, J.G.B. The pollution of the marine environment by plastic debris: A review. Mar. Poll. Bull. 2002, 44, 842–852. [Google Scholar] [CrossRef]
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Plastic waste inputs from land into the ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef]
- Eerkes-Medrano, D.; Thompson, R.C.; Aldridge, D.C. Microplastics in freshwater systems: A review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Res. 2015, 75, 63–82. [Google Scholar] [CrossRef] [PubMed]
- Browne, M.A.; Crump, P.; Niven, S.J.; Teuten, E.; Tonkin, A.; Galloway, T.; Thompson, R. Accumulation of Microplastic on Shorelines Woldwide: Sources and Sinks. Environ. Sci. Technol. 2011, 45, 9175–9179. [Google Scholar] [CrossRef] [PubMed]
- Van Cauwenberghe, L.; Claessens, M.; Vandegehuchte, M.B.; Janssen, C.R. Microplastics are taken up by mussels (Mytilus edulis) and lugworms (Arenicola marina) living in natural habitats. Environ. Poll. 2015, 199, 10–17. [Google Scholar] [CrossRef]
- Kazmiruk, T.N.; Kazmiruk, V.D.; Bendell, L.I. Abundance and distribution of microplastics within surface sediments of a key shellfish growing region of Canada. PLoS ONE 2018, 13, e0196005. [Google Scholar] [CrossRef] [Green Version]
- van Raamsdonk, L.W.D.; van der Zande, M.; Koelmans, A.A.; Hoogenboom, R.L.A.P.; Peters, R.J.B.; Groot, M.J.; Peijnenburg, A.A.C.M.; Weesepoel, Y.J.A. Current Insights into Monitoring, Bioaccumulation, and Potential Health Effects of Microplastics Present in the Food Chain. Foods 2020, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Van Cauwenberghe, L.; Janssen, C.R. Microplastics in bivalves cultured for human consumption. Environ. Poll. 2014, 193, 65–70. [Google Scholar] [CrossRef]
- Gacia, E.; Duarte, C.M. Sediment Retention by a Mediterranean Posidonia oceanica Meadow: The Balance between Deposition and Resuspension. Est. Coast. Shelf Sci. 2001, 52, 505–514. [Google Scholar] [CrossRef]
- Fonseca, M.S.; Zieman, J.C.; Thayer, G.W.; Fisher, J.S. The role of current velocity in structuring eelgrass (Zostera marina L.) meadows. Est. Coast. Shelf Sci. 1983, 17, 367–380. [Google Scholar] [CrossRef]
- Huang, Y.; Xiao, X.; Xu, C.; Perianen, Y.D.; Hu, J.; Holmer, M. Seagrass beds acting as a trap of microplastics—Emerging hotspot in the coastal region? Environ. Poll. 2020, 257, 113450. [Google Scholar] [CrossRef] [PubMed]
- Ling, S.D.; Sinclair, M.; Levi, C.J.; Reeves, S.E.; Edgar, G.J. Ubiquity of microplastics in coastal seafloor sediments. Mar. Poll. Bull. 2017, 121, 104–110. [Google Scholar] [CrossRef]
- Jones, K.L.; Hartl, M.G.J.; Bell, M.C.; Capper, A. Microplastic accumulation in a Zostera marina L. bed at Deerness Sound, Orkney, Scotland. Mar. Poll. Bull. 2020, 152, 110883. [Google Scholar] [CrossRef]
- Seng, N.; Lai, S.; Fong, J.; Saleh, M.F.; Cheng, C.; Cheok, Z.Y.; Todd, P.A. Early evidence of microplastics on seagrass and macroalgae. Mar. Freshw. Res. 2020. [Google Scholar] [CrossRef]
- Goss, H.; Jaskiel, J.; Rotjan, R. Thalassia testudinum as a potential vector for incorporating microplastics into benthic marine food webs. Mar. Poll. Bull. 2018, 135, 1085–1089. [Google Scholar] [CrossRef]
- Pietrelli, L.; Di Gennaro, A.; Menegoni, P.; Lecce, F.; Poeta, G.; Acosta, A.T.R.; Battisti, C.; Iannilli, V. Pervasive plastisphere: First record of plastics in egagropiles (Posidonia spheroids). Environ. Poll. 2017, 229, 1032–1036. [Google Scholar] [CrossRef]
- Remy, F.; Collard, F.; Gilbert, B.; Compère, P.; Eppe, G.; Lepoint, G. When Microplastic Is Not Plastic: The Ingestion of Artificial Cellulose Fibers by Macrofauna Living in Seagrass Macrophytodetritus. Environ. Sci. Technol. 2015, 49, 11158–11166. [Google Scholar] [CrossRef] [PubMed]
- UNEP-WCMC; Short, F.T. Global Distribution of Seagrasses (Version 6.0). Sixth Update to the Data Layer Used in Green and SHORT (2003). Cambridge (UK): UN Environment World Conservation Monitoring Centre. 2018. Available online: http://data.unep-wcmc.org/datasets/7 (accessed on 1 December 2020).
- Ganthy, F.; Soissons, L.; Sauriau, P.G.; Verney, R.; Sottolichio, A. Effects of short flexible seagrass Zostera noltei on flow, erosion and deposition processes determined using flume experiments. Sedimentology 2015, 62, 997–1023. [Google Scholar] [CrossRef] [Green Version]
- Lanuru, M.; Fitri, R. Sediment deposition in South Sulawesi sea-grass bed. Mar. Res. Indones. 2008, 33, 221–224. [Google Scholar] [CrossRef]
- Martin, J.; Lusher, A.; Thompson, R.C.; Morley, A. The Deposition and Accumulation of Microplastics in Marine Sediments and Bottom Water from the Irish Continental Shelf. Sci. Rep. 2017, 7, 10772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, J.; Sobral, P. Plastic marine debris on the Portuguese coastline: A matter of size? Mar. Poll. Bull. 2011, 62, 2649–2653. [Google Scholar] [CrossRef]
- Löder, M.G.J.; Gerdts, G. Methodology Used for the Detection and Identification of Microplastics—A Critical Appraisal. In Marine Anthropogenic Litter; Bergmann, M., Gutow, L., Klages, M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 201–227. [Google Scholar]
- Dethier, M.N.; Graham, E.S.; Cohen, S.; Tear, L.M. Visual versus random-point percent cover estimates: ‘objective’ is not always better. Mar. Ecol. Prog. Ser. 1993, 96, 93–100. [Google Scholar] [CrossRef]
- McKenzie, L.J.; Campbell, S.J.; Roder, C.A. Seagrass-Watch: Manual for Mapping & Monitoring Seagrass Resources by Community (Citizen) Volunteers; Queensland Fisheries Service, Department of Primary Industries: Cairns, Switzerland, 2001; p. 100. [Google Scholar]
- Wentworth, C.K. A Scale of Grade and Class Terms for Clastic Sediments. J. Geol. 1922, 30, 377–392. [Google Scholar] [CrossRef]
- Hidalgo-Ruz, V.; Gutow, L.; Thompson, R.C.; Thiel, M. Microplastics in the Marine Environment: A Review of the Methods Used for Identification and Quantification. Environ. Sci. Technol. 2012, 46, 3060–3075. [Google Scholar] [CrossRef] [PubMed]
- Imhof, H.K.; Schmid, J.; Niessner, R.; Ivleva, N.P.; Laforsch, C. A novel, highly efficient method for the separation and quantification of plastic particles in sediments of aquatic environments. Limnol. Oceanogr. Methods 2012, 10, 524–537. [Google Scholar] [CrossRef]
- Coppock, R.L.; Cole, M.; Lindeque, P.K.; Queirós, A.M.; Galloway, T.S. A small-scale, portable method for extracting microplastics from marine sediments. Environ. Poll. 2017, 230, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Baltic, C.C. Guide to Microplastic Identification; Marine and Environmental Research Institute: Blue Hill, ME, USA, 2017. [Google Scholar]
- Suaria, G.; Achtypi, A.; Perold, V.; Lee, J.R.; Pierucci, A.; Bornman, T.G.; Aliani, S.; Ryan, P.G. Microfibers in oceanic surface waters: A global characterization. Sci. Adv. 2020, 6, eaay8493. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Lawrence Erlbaum Associates: New Jersey, NJ, USA, 1988. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Usinglme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Lefcheck, J.S. piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Sanchez-Vidal, A.; Canals, M.; de Haan, W.P.; Romero, J.; Veny, M. Seagrasses provide a novel ecosystem service by trapping marine plastics. Sci. Rep. 2021, 11, 254. [Google Scholar] [CrossRef]
- Hartline, N.L.; Bruce, N.J.; Karba, S.N.; Ruff, E.O.; Sonar, S.U.; Holden, P.A. Microfiber Masses Recovered from Conventional Machine Washing of New or Aged Garments. Environ. Sci. Technol. 2016, 50, 11532–11538. [Google Scholar] [CrossRef] [Green Version]
- Bessa, F.; Barría, P.; Neto, J.M.; Frias, J.P.G.L.; Otero, V.; Sobral, P.; Marques, J.C. Occurrence of microplastics in commercial fish from a natural estuarine environment. Mar. Poll. Bull. 2018, 128, 575–584. [Google Scholar] [CrossRef]
- de los Santos, C.B.; Krång, A.-S.; Infantes, E. Microplastic retention by marine vegetated canopies: Simulations with seagrass meadows in a hydraulic flume. Environ. Poll. 2021, 269, 116050. [Google Scholar] [CrossRef] [PubMed]
- Jabeen, K.; Su, L.; Li, J.; Yang, D.; Tong, C.; Mu, J.; Shi, H. Microplastics and mesoplastics in fish from coastal and fresh waters of China. Environ. Poll. 2017, 221, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Lusher, A.L.; McHugh, M.; Thompson, R.C. Occurrence of microplastics in the gastrointestinal tract of pelagic and demersal fish from the English Channel. Mar. Poll. Bull. 2013, 67, 94–99. [Google Scholar] [CrossRef]
- Chakraborty, P.; Sarkar, A.; Vudamala, K.; Naik, R.; Nath, B.N. Organic matter—A key factor in controlling mercury distribution in estuarine sediment. Mar. Chem. 2015, 173, 302–309. [Google Scholar] [CrossRef]
- Schorer, M. Pollutant and organic matter content in sediment particle size fractions. Int. Assoc. Hydrol. Sci. 1997, 243, 59–68. [Google Scholar]
- Alomar, C.; Estarellas, F.; Deudero, S. Microplastics in the Mediterranean Sea: Deposition in coastal shallow sediments, spatial variation and preferential grain size. Mar. Environ. Res. 2016, 115, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hardesty, B.D.; Harari, J.; Isobe, A.; Lebreton, L.; Maximenko, N.; Potemra, J.; van Sebille, E.; Vethaak, A.D.; Wilcox, C. Using Numerical Model Simulations to Improve the Understanding of Micro-plastic Distribution and Pathways in the Marine Environment. Front. Mar. Sci. 2017, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Röhr, M.E.; Holmer, M.; Baum, J.K.; Björk, M.; Chin, D.; Chalifour, L.; Cimon, S.; Cusson, M.; Dahl, M.; Deyanova, D.; et al. Blue Carbon Storage Capacity of Temperate Eelgrass (Zostera marina) Meadows. Glob. Biogeochem. Cycles 2018, 32, 1457–1475. [Google Scholar] [CrossRef] [Green Version]
- Bertelli, C.M.; Unsworth, R.K.F. Protecting the hand that feeds us: Seagrass (Zostera marina) serves as commercial juvenile fish habitat. Mar. Poll. Bull. 2014, 83, 425–429. [Google Scholar] [CrossRef]
- Lilley, R.; Unsworth, R.K.F. Atlantic Cod (Gadus morhua) benefits from the availability of seagrass (Zostera marina) nursery habitat. Glob. Ecol. Conserv. 2014, 2, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Furness, E.; Unsworth, R.K.F. Demersal Fish Assemblages in NE Atlantic Seagrass and Kelp. Diversity 2020, 12, 366. [Google Scholar] [CrossRef]
- Nordlund, L.; Unsworth, R.; Gullström, M.; Cullen-Unsworth, L. Global significance of seagrass fishery activity. Fish Fish. 2018, 19, 399–412. [Google Scholar] [CrossRef]
- Green, A.E.; Unsworth, R.K.F.; Chadwick, M.A.; Jones, P.J. Historical analysis exposes catastrophic seagrass loss for the United Kingdom. Front. Plant Sci. 2021. In Press. [Google Scholar]
- Deudero, S.; Alomar, C. Mediterranean marine biodiversity under threat: Reviewing influence of marine litter on species. Mar. Poll. Bull. 2015, 98, 58–68. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seagrass | Unvegetated | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source | Media | Location | No. of Locations | Location Type | Dominant Seagrass Species | Sub- (S) or Inter-tidal (I) | n | MP Abundance (MP kg−1 DW) | % of Samples Contain MP | Dominant MP Form | MP (MP kg−1 DW) | % of Samples Contain MP | Dominant MP Form | Effect Size (Cohens d) |
| Tahir et al. 2019 | ZnBr | Indonesia | 3 | Remote island | Halophila, Cymodocea, Enhalus, Thalassia, Syringodium, Halodule | S | 81 | 131 ± 100 | 27 | Fibres Filaments | NA | NA | NA | 1.31 |
| Huang et al. 2020 | ZnCl2 | China | 2 | Coastal | Enhalus acodoides | I | 12 (3 each) | 196.7 ± 16.1 | 100 | Fibres Filaments | 93.3 ± 15.3 | 100 | Fibres Filaments | 6.5 |
| 780.2 ± 147.0 | 267.1 ± 60.5 | 4.9 | ||||||||||||
| Jones et al. 2020 | NaCl | Scotland | 1 | Island | Zostera marina | S | 25 | 300 ± 30 (20) | 100 | Fibres | 110 ± 20 (5) | 100 | Fibres | 7.6 |
| Cozzolino et al. 2020 | NaCl | Portugal | 1 | Lagoon | Z. marina | I | 40 (10 each) | 18.2 ± 15.5 (0–11) | 100 | Fibres | 29.8 ± 14.9 (3–15) | 100 | Fibres | −0.76 |
| Cymodocea nodosa, Z. marina | S | 35.2 ± 26.7 (1–21) | 20.4 ± 14.5 (1–10) | 0.71 | ||||||||||
| Plee and Pomory 2020 | NaCl | USA | 1 | Lagoon | NA | S | 6 | 24 ± 6 | 100 | Fibres Fragments | 15 ± 3 | 100 | Fibres | 2 |
| Our study | ZnCl | England and Wales | 8 | Coastal and Estuarine | Z.marina Z. noltii | I | 80 (40 each) | 215 ± 163 | 98 | Fibres Filaments | 221 ± 236 | 98 | Fibres Filaments | −0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Unsworth, R.K.F.; Higgs, A.; Walter, B.; Cullen-Unsworth, L.C.; Inman, I.; Jones, B.L. Canopy Accumulation: Are Seagrass Meadows a Sink of Microplastics? Oceans 2021, 2, 162-178. https://doi.org/10.3390/oceans2010010
Unsworth RKF, Higgs A, Walter B, Cullen-Unsworth LC, Inman I, Jones BL. Canopy Accumulation: Are Seagrass Meadows a Sink of Microplastics? Oceans. 2021; 2(1):162-178. https://doi.org/10.3390/oceans2010010
Chicago/Turabian StyleUnsworth, Richard K. F., Alex Higgs, Bettina Walter, Leanne C. Cullen-Unsworth, Isabella Inman, and Benjamin L. Jones. 2021. "Canopy Accumulation: Are Seagrass Meadows a Sink of Microplastics?" Oceans 2, no. 1: 162-178. https://doi.org/10.3390/oceans2010010
APA StyleUnsworth, R. K. F., Higgs, A., Walter, B., Cullen-Unsworth, L. C., Inman, I., & Jones, B. L. (2021). Canopy Accumulation: Are Seagrass Meadows a Sink of Microplastics? Oceans, 2(1), 162-178. https://doi.org/10.3390/oceans2010010