
The Condition of Four Coral Reefs in Timor-Leste before and after the 2016–2017 Marine Heatwave



Abstract
:1. Introduction
1.1. Disease in the Context of Coral Reef Health
1.2. Water Quality and Coral Reefs along the North Coast of Timor-Leste
1.3. Global Impacts—Ocean Warming, Mass Coral Bleaching, and Mortality
1.4. Aims
2. Materials and Methods
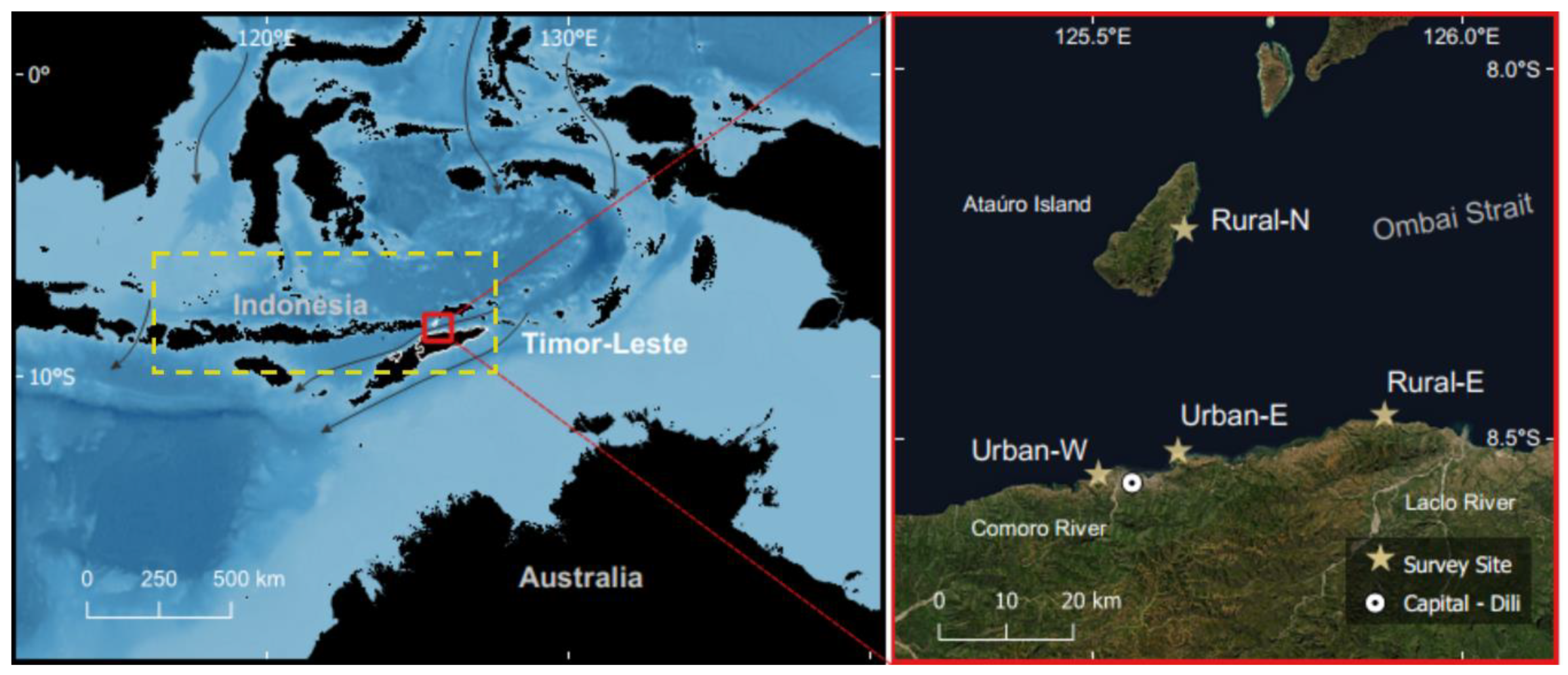
2.1. Study Site

2.2. Coral Community Composition and Coral Health Surveys
2.3. Measurement of Nutrient Concentrations and Stable Isotope Ratios
2.4. In Situ and Satellite Temperature Data
2.5. Statistical Analyses
3. Results
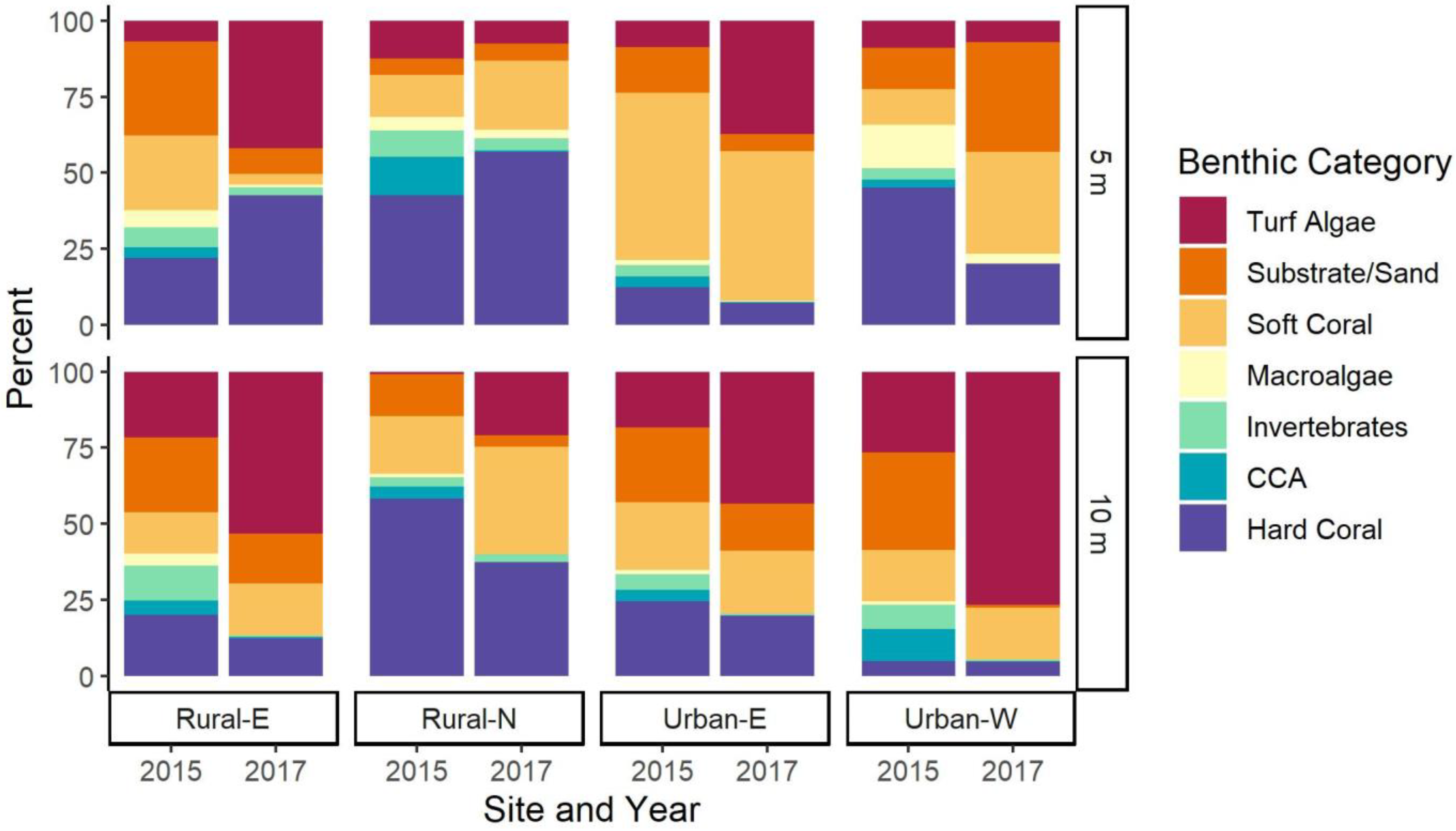
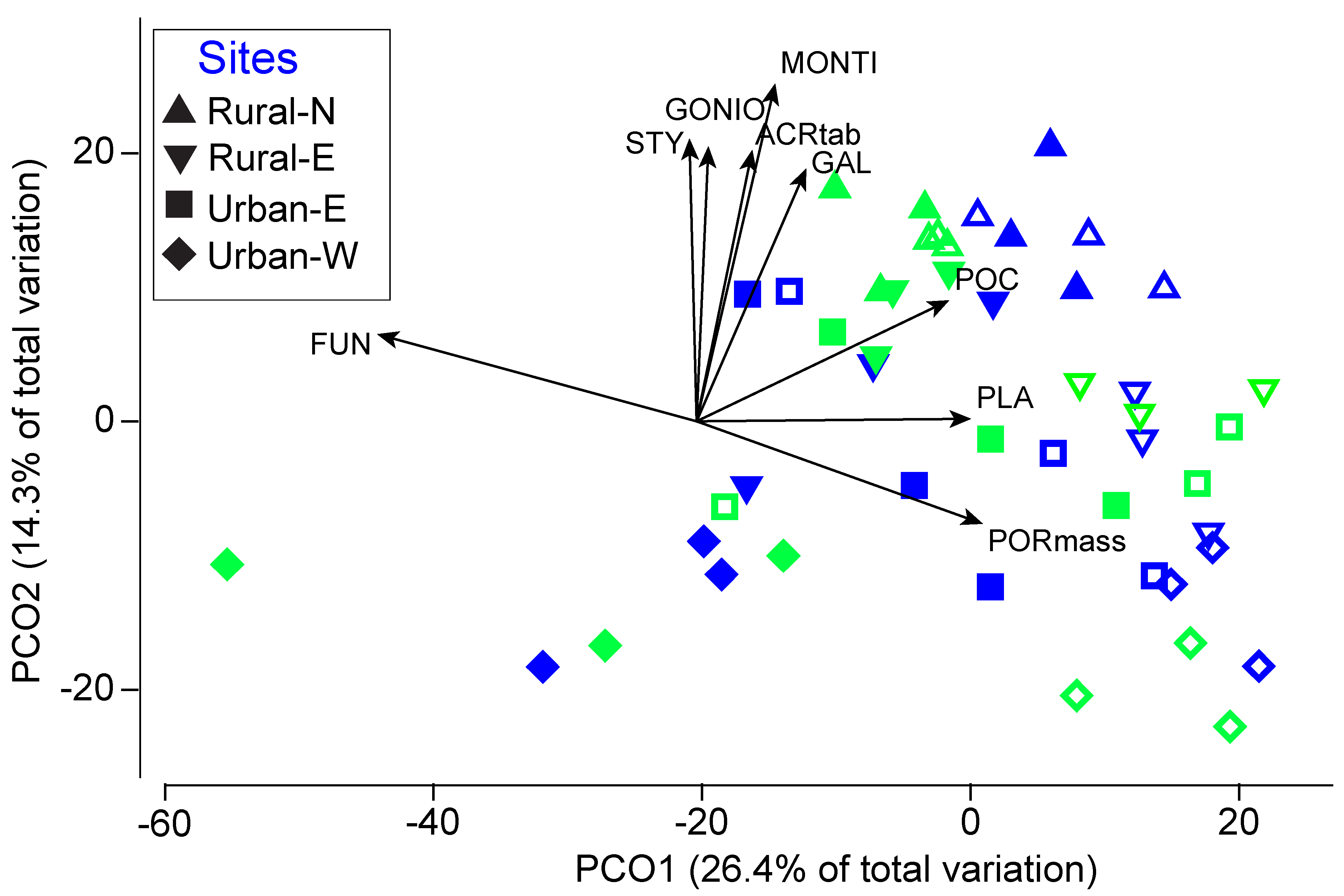
3.1. Coral Cover and Community Composition at Four Sites
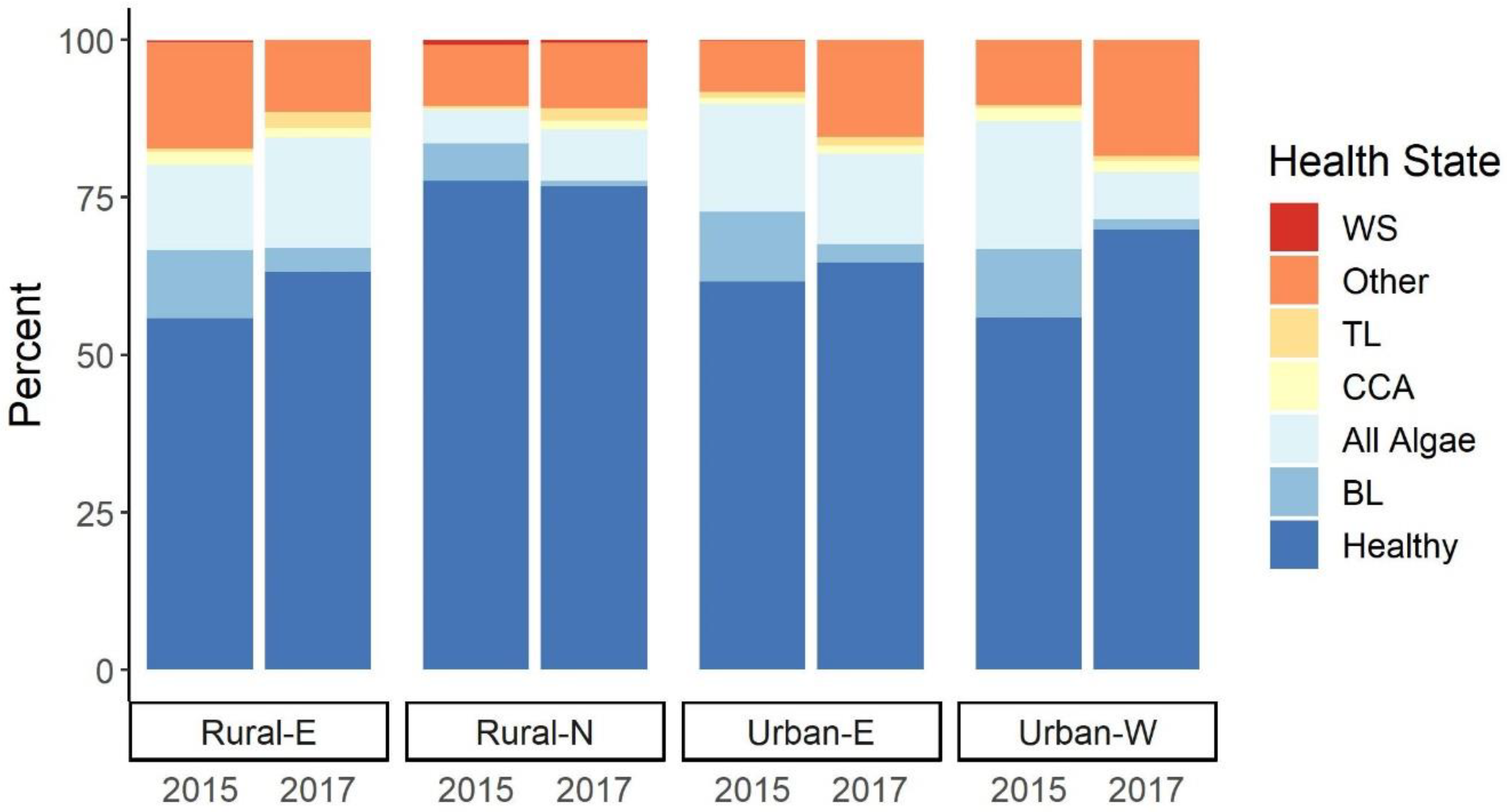
3.2. Prevalence of Coral Disease and Indicators of Compromised Health
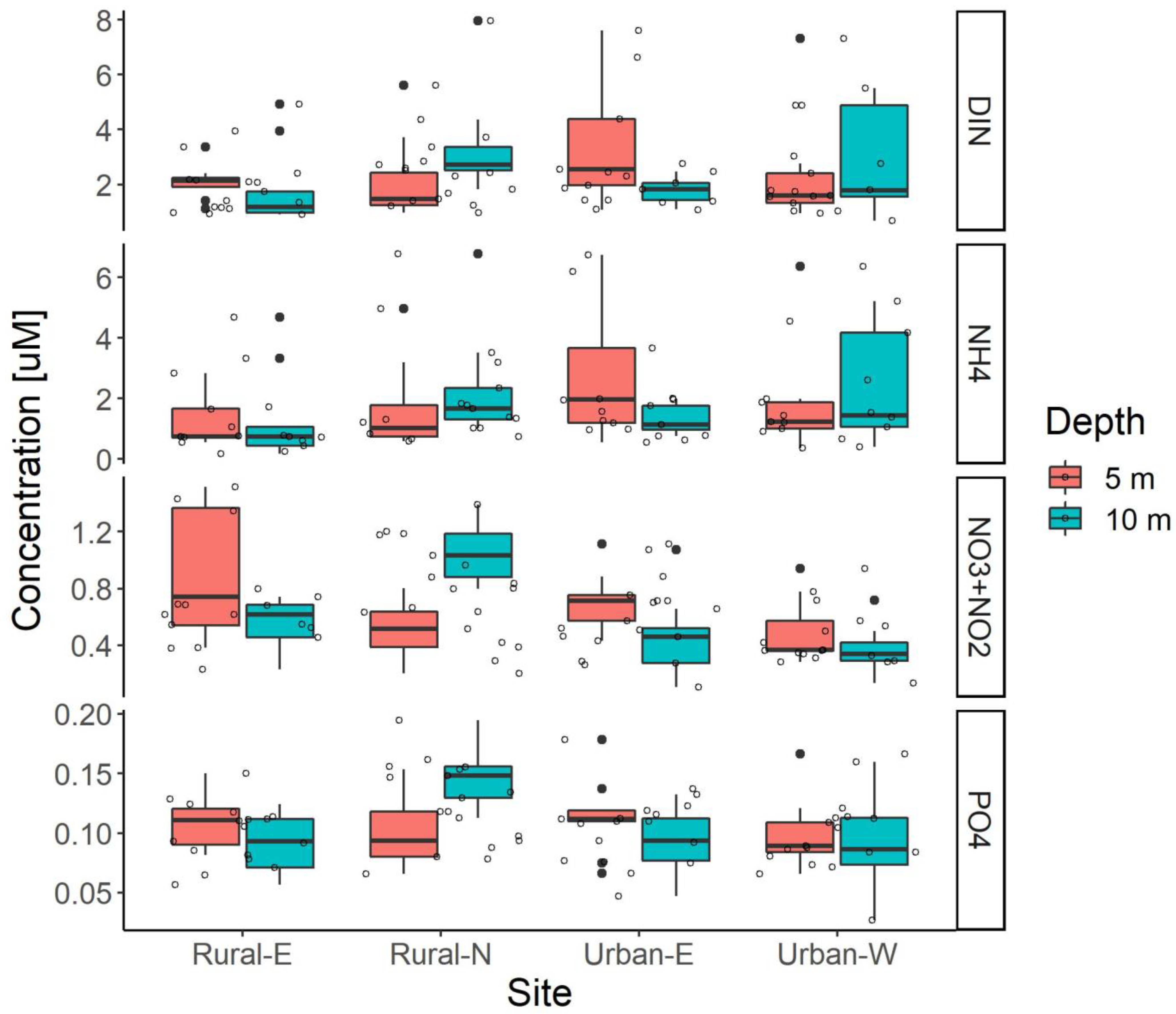
3.3. Water Quality
Nutrients and Stable Isotopes
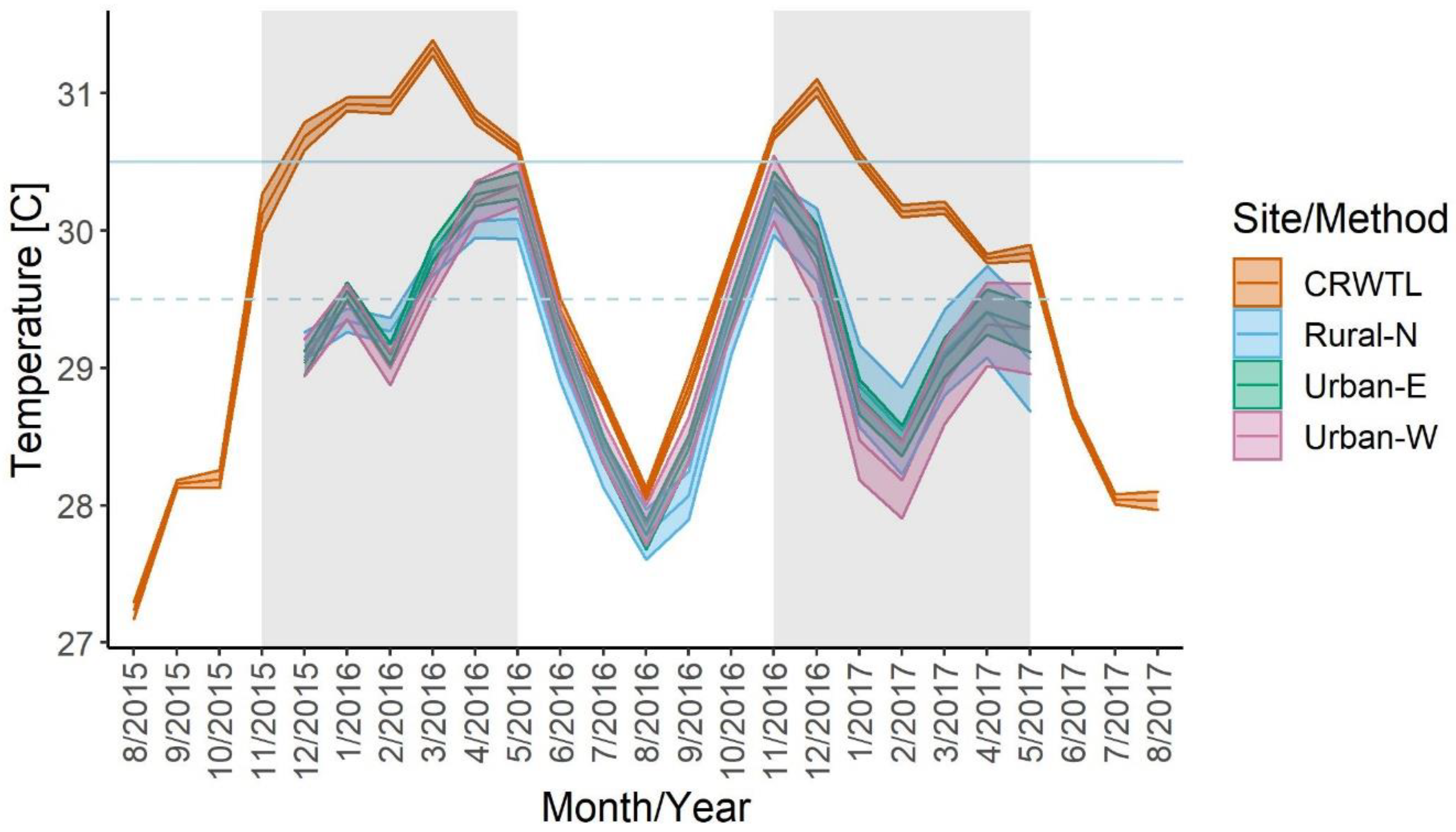
3.4. Temperature and the Prevalence of Bleaching
4. Discussion
4.1. Coral Community Composition and Human Impacts
4.2. The Health of Coral Reefs along the North Coast of Timor-Leste
4.3. Water Quality and Sources of Nutrients in Timor-Leste
4.4. Elevated Temperature and the Prevalence of Bleaching from Thermal Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burke, L.; Reytar, K.; Spalding, M.; Perry, A. Reefs at Risk Revisited; World Resources Institute: Washington, DC, USA, 2011; p. e0116200. [Google Scholar]
- Veron, J.E.N.; DeVantier, L.M.; Turak, E.; Green, A.L.; Kininmonth, S.; Stafford-Smith, M.; Peterson, N. Delineating the Coral Triangle. Galaxea J. Coral Reef Stud. 2009, 11, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Harvell, D.; Jordán-Dahlgren, E.; Merkel, S.; Rosenberg, E.; Raymundo, L.; Smith, G.; Weil, E.; Willis, B. Coral Disease, Environmental Drivers, and the Balance Between Coral and Microbial Associates. Oceanography 2007, 20, 172–195. [Google Scholar] [CrossRef] [Green Version]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral Reefs Under Rapid Climate Change and Ocean Acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, J.B.C.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A.; et al. Historical Overfishing and the Recent Collapse of Coastal Ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshw. Res. 1999, 50, 839–866. [Google Scholar] [CrossRef] [Green Version]
- Heron, S.F.; Maynard, J.A.; Van Hooidonk, R.; Eakin, C.M. Warming Trends and Bleaching Stress of the World’s Coral Reefs 1985–2012. Sci. Rep. 2016, 6, 38402. [Google Scholar] [CrossRef] [Green Version]
- Eakin, C.M.; Liu, G.; Gomez, A.M.; De La Cour, J.L.; Heron, S.F.; Skirving, W.J.; Geiger, E.F.; Marsh, B.L.; Tirak, K.V.; Strong, A.E.; et al. The Witch Is Dead (?)—Three Years of Global Coral Bleaching 2014–2017. Reef Encount. 2017, 32, 33–38. [Google Scholar]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C.; et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Ampou, E.E.; Johan, O.; Menkes, C.E.; Niño, F.; Birol, F.; Ouillon, S.; Andréfouët, S. Coral mortality induced by the 2015–2016 El-Niño in Indonesia: The effect of rapid sea level fall. Biogeosciences 2017, 14, 817–826. [Google Scholar] [CrossRef] [Green Version]
- Burke, L.; Reytar, K.; Spalding, M.; Perry, A. Reefs at Risk Revisited in the Coral Triangle; World Resources Institute: Washington, DC, USA, 2012; pp. 1–72. [Google Scholar]
- ADB. State of the Coral Triangle: Timor-Leste; Asian Development Bank: Mandaluyong City, Philippines, 2014; p. 57. [Google Scholar]
- Macaulay, J. Timor Leste: Newest and Poorest of Asian Nations. Geography 2003, 88, 40–46. [Google Scholar]
- Barbosa, M.; Booth, S. Timor-Leste’s Fisheries Catches (1950–2009): Fisheries under Different Regimes. In Fisheries Catch Reconstructions: Islands, Part I; Zeller, D., Harper, S., Eds.; Fisheries Centre, University of British Columbia: Vancouver, BC, Canada, 2009; Volume 17, pp. 39–52. [Google Scholar]
- Kingsbury, D.; Soares, D.B.; Harris, V.; Fox, J.J.; Bateman, S.; Bergin, A. A Reliable Partner: Strengthening Australia—Timor-Leste Relations. Aust. Strateg. Policy Inst. Ltd. 2011, 39, 68. [Google Scholar]
- McWilliam, A. Perspectives on Customary Marine Tenures in East Timor. Asia Pac. J. Anthropol. 2002, 3, 6–32. [Google Scholar] [CrossRef]
- Tilley, A.; Burgos, A.; Duarte, A.; Lopes, J.D.R.; Eriksson, H.; Mills, D. Contribution of women’s fisheries substantial, but overlooked, in Timor-Leste. AMBIO 2021, 50, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Grantham, R.; Álvarez-Romero, J.G.; Mills, D.J.; Rojas, C.; Cumming, G.S. Spatiotemporal determinants of seasonal gleaning. People Nat. 2021, 3, 376–390. [Google Scholar] [CrossRef]
- RDTL. Timor-Leste Strategic Development Plan 2011–2030; Republica Democratica de Timor-Leste: Dili, Timor-Leste, 2011; pp. 1–215.
- JICA. The Study on Community-Based Integrated Watershed Management in Laclo and Comoro River Basins in the Democratic Republic of Timor-Leste; Japan International Cooperation Agency: Tokyo, Japan, 2010; pp. 1–43.
- Sandlund, O.T.; Bryceson, I.; de Carvalho, D.; Rio, N.; da Silva, J.; Silva, M.I. Assessing Environmental Needs and Priorities in Timor-Leste; United Nations Development Programme (UNDP) Commissioned Paper; UNDP: New York, NY, USA, 2001. [Google Scholar]
- Alongi, D.M.; Amaral, A.; de Carvalho, N.; McWilliam, A.; Rouwenhorst, J.; Tirendi, F.; Trott, L.; Wasson, R.J. River Catchments and Marine Productivity in Timor Leste: Caraulun and Laclo Catchments; South and North Coasts—Final Report; Ministry of Agriculture & Fisheries, Government of Timor Leste: Dili, Timor-Leste, 2012.
- Aronson, R.; Precht, W.F. White-band disease and the changing face of Caribbean coral reefs. Hydrobiologia 2001, 460, 25–38. [Google Scholar] [CrossRef]
- Myers, R.; Raymundo, L. Coral disease in Micronesian reefs: A link between disease prevalence and host abundance. Dis. Aquat. Org. 2009, 87, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Weil, E.; Irikawa, A.; Casareto, B.; Suzuki, Y. Extended geographic distribution of several Indo-Pacific coral reef diseases. Dis. Aquat. Org. 2012, 98, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Willis, B.L.; Page, C.A.; Dinsdale, E.A. Coral Disease on the Great Barrier Reef. In Coral Health and Disease; Springer: Berlin/Heidelberg, Germnay, 2004; pp. 69–104. [Google Scholar] [CrossRef]
- Beeden, R.; Willis, B.L.; Page, C.A.; Weil, E. Underwater Cards for Assessing Coral Health on Indo-Pacific Reefs; Coral Reef Targeted Research and Capacity Building for Management Program, Currie Communications: Melbourne, Australia, 2008. [Google Scholar]
- Raymundo, L.J.; Couch, C.S.; Bruckner, A.W.; Harvell, C.D. Coral Disease Handbook Guidelines for Assessment: Guidelines for Assessment Monitoring and Management; Currie Communications: Melbourne, Australia, 2008; ISBN 978-1-921317-01-9. [Google Scholar]
- Bruno, J.F.; Selig, E.R.; Casey, K.; Page, C.A.; Willis, B.L.; Harvell, C.D.; Sweatman, H.; Melendy, A.M. Thermal Stress and Coral Cover as Drivers of Coral Disease Outbreaks. PLoS Biol. 2007, 5, e124. [Google Scholar] [CrossRef]
- Pollock, F.J.; Lamb, J.; Field, S.N.; Heron, S.; Schaffelke, B.; Shedrawi, G.; Bourne, D.G.; Willis, B.L. Sediment and Turbidity Associated with Offshore Dredging Increase Coral Disease Prevalence on Nearby Reefs. PLoS ONE 2014, 9, e102498. [Google Scholar] [CrossRef] [Green Version]
- Raymundo, L.J.; Halford, A.R.; Maypa, A.P.; Kerr, A.M. Functionally diverse reef-fish communities ameliorate coral disease. Proc. Natl. Acad. Sci. USA 2009, 106, 17067–17070. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, R.M.; Kim, C.J.; Tracy, A.M.; Most, R.; Harvell, C.D. Linking sewage pollution and water quality to spatial patterns of Porites lobata growth anomalies in Puako, Hawaii. Mar. Pollut. Bull. 2016, 104, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef] [PubMed]
- Aronson, R.B.; Macintyre, I.G.; Wapnick, C.M.; O’Neill, M.W. Phase Shifts, Alternative States, and the Unprecedented Convergence of Two Reef Systems. Ecology 2004, 85, 1876–1891. [Google Scholar] [CrossRef]
- Cleary, D.F.R.; Suharsono; Hoeksema, B.W. Coral diversity across a disturbance gradient in the Pulau Seribu reef complex off Jakarta, Indonesia. In Marine, Freshwater, and Wetlands Biodiversity Conservation; Hawksworth, D.L., Bull, A.T., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 285–306. [Google Scholar]
- Baker, D.; MacAvoy, S.; Kim, K. Relationship between water quality, Δ15N, and aspergillosis of Caribbean sea fan corals. Mar. Ecol. Prog. Ser. 2007, 343, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Thurber, R.L.V.; Burkepile, D.E.; Fuchs, C.; Shantz, A.; McMinds, R.; Zaneveld, J.R. Chronic nutrient enrichment increases prevalence and severity of coral disease and bleaching. Glob. Change Biol. 2013, 20, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Voss, J.D.; Richardson, L.L. Nutrient enrichment enhances black band disease progression in corals. Coral Reefs 2006, 25, 569–576. [Google Scholar] [CrossRef]
- Wagner, D.; Kramer, P.; Van Woesik, R. Species composition, habitat, and water quality influence coral bleaching in southern Florida. Mar. Ecol. Prog. Ser. 2010, 408, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Wooldridge, S.A.; Done, T.J. Improved water quality can ameliorate effects of climate change on corals. Ecol. Appl. 2009, 19, 1492–1499. [Google Scholar] [CrossRef]
- Amato, D.W.; Bishop, J.M.; Glenn, C.R.; Dulai, H.; Smith, C.M. Impact of Submarine Groundwater Discharge on Marine Water Quality and Reef Biota of Maui. PLoS ONE 2016, 11, e0165825. [Google Scholar] [CrossRef] [Green Version]
- Dinsdale, E.A.; Pantos, O.; Smriga, S.; Edwards, R.; Angly, F.; Wegley, L.; Hatay, M.; Hall, D.; Brown, E.; Haynes, M.; et al. Microbial Ecology of Four Coral Atolls in the Northern Line Islands. PLoS ONE 2008, 3, e1584. [Google Scholar] [CrossRef] [Green Version]
- Osawa, Y.; Fujita, K.; Umezawa, Y.; Kayanne, H.; Ide, Y.; Nagaoka, T.; Miyajima, T.; Yamano, H. Human impacts on large benthic foraminifers near a densely populated area of Majuro Atoll, Marshall Islands. Mar. Pollut. Bull. 2010, 60, 1279–1287. [Google Scholar] [CrossRef]
- Smith, J.; Smith, C.; Hunter, C. An experimental analysis of the effects of herbivory and nutrient enrichment on benthic community dynamics on a Hawaiian reef. Coral Reefs 2001, 19, 332–342. [Google Scholar] [CrossRef]
- Aeby, G.S.; Williams, G.J.; Franklin, E.C.; Kenyon, J.; Cox, E.F.; Coles, S.; Work, T.M. Patterns of Coral Disease across the Hawaiian Archipelago: Relating Disease to Environment. PLoS ONE 2011, 6, e20370. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.F.; Petes, L.E.; Harvell, C.D.; Hettinger, A. Nutrient enrichment can increase the severity of coral diseases. Ecol. Lett. 2003, 6, 1056–1061. [Google Scholar] [CrossRef]
- Kaczmarsky, L.; Richardson, L.L. Do elevated nutrients and organic carbon on Philippine reefs increase the prevalence of coral disease? Coral Reefs 2011, 30, 253–257. [Google Scholar] [CrossRef]
- Costanzo, S.; O’Donohue, M.; Dennison, W.; Loneragan, N.; Thomas, M. A New Approach for Detecting and Mapping Sewage Impacts. Mar. Pollut. Bull. 2001, 42, 149–156. [Google Scholar] [CrossRef]
- Dailer, M.L.; Ramey, H.L.; Saephan, S.; Smith, C.M. Algal Δ15N values detect a wastewater effluent plume in nearshore and offshore surface waters and three-dimensionally model the plume across a coral reef on Maui, Hawai‘i, USA. Mar. Pollut. Bull. 2012, 64, 207–213. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Barile, P.J.; Littler, M.M.; Littler, D.S. Macroalgal blooms on southeast Florida coral reefs: II. Cross-shelf discrimination of nitrogen sources indicates widespread assimilation of sewage nitrogen. Harmful Algae 2005, 4, 1106–1122. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Barile, P.J.; Matzie, W.R. Anthropogenic Nutrient Enrichment of Seagrass and Coral Reef Communities in the Lower Florida Keys: Discrimination of Local versus Regional Nitrogen Sources. J. Exp. Mar. Biol. Ecol. 2004, 308, 23–58. [Google Scholar] [CrossRef]
- Lin, H.-J.; Wu, C.; Kao, S.; Kao, W.; Meng, P. Mapping anthropogenic nitrogen through point sources in coral reefs using Δ15N in macroalgae. Mar. Ecol. Prog. Ser. 2007, 335, 95–109. [Google Scholar] [CrossRef]
- Moynihan, M.; Baker, D.M.; Mmochi, A.J. Isotopic and microbial indicators of sewage pollution from Stone Town, Zanzibar, Tanzania. Mar. Pollut. Bull. 2012, 64, 1348–1355. [Google Scholar] [CrossRef] [PubMed]
- Redding, J.E.; Myers-Miller, R.L.; Baker, D.M.; Fogel, M.; Raymundo, L.J.; Kim, K. Link between sewage-derived nitrogen pollution and coral disease severity in Guam. Mar. Pollut. Bull. 2013, 73, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Savage, C.; Elmgren, R. Macroalgal (fucus vesiculosus) Δ15n values trace decrease in sewage influence. Ecol. Appl. 2004, 14, 517–526. [Google Scholar] [CrossRef]
- Sutherland, K.P.; Porter, J.W.; Turner, J.W.; Thomas, B.J.; Looney, E.E.; Luna, T.P.; Meyers, M.K.; Futch, J.C.; Lipp, E.K. Human sewage identified as likely source of white pox disease of the threatened Caribbean elkhorn coral, Acropora palmata. Environ. Microbiol. 2010, 12, 1122–1131. [Google Scholar] [CrossRef]
- Umezawa, Y.; Miyajima, T.; Kayanne, H.; Koike, I. Significance of groundwater nitrogen discharge into coral reefs at Ishigaki Island, southwest of Japan. Coral Reefs 2002, 21, 346–356. [Google Scholar] [CrossRef]
- Fry, B.; Baltz, D.M.; Benfield, M.C.; Fleeger, J.W.; Gace, A.; Haas, H.L.; Quiñones-Rivera, Z.J. Stable isotope indicators of movement and residency for brown shrimp (Farfantepenaeus aztecus) in coastal Louisiana marshscapes. Estuaries 2003, 26, 82–97. [Google Scholar] [CrossRef]
- Dailer, M.L.; Knox, R.S.; Smith, J.E.; Napier, M.; Smith, C.M. Using Δ15N values in algal tissue to map locations and potential sources of anthropogenic nutrient inputs on the island of Maui, Hawai‘i, USA. Mar. Pollut. Bull. 2010, 60, 655–671. [Google Scholar] [CrossRef] [PubMed]
- Gartner, A.; Lavery, P.; Smit, A. Use of d15N signatures of different functional forms of macroalgae and filter-feeders to reveal temporal and spatial patterns in sewage dispersal. Mar. Ecol. Prog. Ser. 2002, 235, 63–73. [Google Scholar] [CrossRef]
- Heaton, T. Isotopic studies of nitrogen pollution in the hydrosphere and atmosphere: A review. Chem. Geol. Isot. Geosci. Sect. 1986, 59, 87–102. [Google Scholar] [CrossRef]
- Tucker, J.; Sheats, N.; Giblin, A.; Hopkinson, C.; Montoya, J. Using stable isotopes to trace sewage-derived material through Boston Harbor and Massachusetts Bay. Mar. Environ. Res. 1999, 48, 353–375. [Google Scholar] [CrossRef]
- Montoya, J.P.; Carpenter, E.J.; Capone, D.G. Nitrogen fixation and nitrogen isotope abundances in zooplankton of the oligotrophic North Atlantic. Limnol. Oceanogr. 2002, 47, 1617–1628. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Li, X.B.; Titlyanov, E.A.; Ye, C.; Titlyanova, T.V.; Guo, Y.P.; Zhang, J. Linking macroalgal Δ15N-values to nitrogen sources and effects of nutrient stress on coral condition in an upwelling region. Bot. Mar. 2013, 56, 471–480. [Google Scholar] [CrossRef]
- Lamb, K.; Swart, P.; Altabet, M. Nitrogen and Carbon Isotopic Systematics of the Florida Reef Tract. Bull. Mar. Sci. 2012, 88, 119–146. [Google Scholar] [CrossRef] [Green Version]
- Leichter, J.J.; Wankel, S.; Paytan, A.; Hanson, K.; Miller, S.; Altabet, M.A. Nitrogen and oxygen isotopic signatures of subsurface nitrate seaward of the Florida Keys reef tract. Limnol. Oceanogr. 2007, 52, 1258–1267. [Google Scholar] [CrossRef] [Green Version]
- Sigman, D.M.; Altabet, M.A.; McCorkle, D.C.; Francois, R.; Fischer, G.E. The Δ15N of nitrate in the Southern Ocean: Nitrogen cycling and circulation in the ocean interior. J. Geophys. Res. Earth Surf. 2000, 105, 19599–19614. [Google Scholar] [CrossRef]
- Firstater, F.N.; Hidalgo, F.J.; Lomovasky, B.; Tarazona, J.; Flores, G.; Iribarne, O.O. Coastal upwelling may overwhelm the effect of sewage discharges in rocky intertidal communities of the Peruvian coast. Mar. Freshw. Res. 2010, 61, 309–319. [Google Scholar] [CrossRef]
- Radice, V.Z.; Hoegh-Guldberg, O.; Fry, B.; Fox, M.D.; Dove, S.G. Upwelling as the major source of nitrogen for shallow and deep reef-building corals across an oceanic atoll system. Funct. Ecol. 2019, 33, 1120–1134. [Google Scholar] [CrossRef]
- Radice, V.Z.; Fry, B.; Dove, S.G.; Hoegh-Guldberg, O. Biogeochemical variability and trophic status of reef water column following a coral bleaching event. Coral Reefs 2021, 40, 1–7. [Google Scholar] [CrossRef]
- NOAA Coral Reef Watch NOAA Coral Reef Watch Version 3.0 Daily Gobal 50km Satellite Virtual Station Time Series Data for Timor-Leste, 1 August 2015–31 August 2017. Available online: https://coralreefwatch.noaa.gov/product/vs/timeseries/coral_triangle.php#timor_leste (accessed on 15 September 2017).
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef]
- Strong, A.E.; Liu, G.; Skirving, W.; Eakin, C.M. NOAA’s Coral Reef Watch program from satellite observations. Ann. GIS 2011, 17, 83–92. [Google Scholar] [CrossRef]
- Liu, G.; Strong, A.E.; Skirving, W.; Arzayus, L.F. Overview of NOAA Coral Reef Watch Program’s Near-Real-Time Satellite Global Coral Bleaching Monitoring Activities. In Proceedings of the 10th International Coral Reef Symposium, Okinawa, Japan; 2006; pp. 1783–1793. [Google Scholar]
- Baird, A.H.; Marshall, P.A. Mortality, growth and reproduction in scleractinian corals following bleaching on the Great Barrier Reef. Mar. Ecol. Prog. Ser. 2002, 237, 133–141. [Google Scholar] [CrossRef]
- Marshall, P.A.; Baird, A.H. Bleaching of corals on the Great Barrier Reef: Differential susceptibilities among taxa. Coral Reefs 2000, 19, 155–163. [Google Scholar] [CrossRef]
- Harvell, C.D.; Kim, K.; Burkholder, J.M.; Colwell, R.R.; Epstein, P.R.; Grimes, D.J.; Hofmann, E.E.; Lipp, E.K.; Osterhaus, A.D.M.E.; Overstreet, R.M.; et al. Emerging Marine Diseases--Climate Links and Anthropogenic Factors. Science 1999, 285, 1505–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvell, C.D.; Mitchell, C.E.; Ward, J.R.; Altizer, S.; Dobson, A.P.; Ostfeld, R.S.; Samuel, M.D. Climate Warming and Disease Risks for Terrestrial and Marine Biota. Science 2002, 296, 2158–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodnicke, O.B.; Bourne, D.G.; Heron, S.F.; Pears, R.J.; Stella, J.S.; Smith, H.A.; Willis, B.L. Unravelling the links between heat stress, bleaching and disease: Fate of tabular corals following a combined disease and bleaching event. Coral Reefs 2019, 38, 591–603. [Google Scholar] [CrossRef]
- van der Zande, R.M.; Achlatis, M.; Bender-Champ, D.; Kubicek, A.; Dove, S.; Hoegh-Guldberg, O. Paradise lost: End-of-century warming and acidification under business-as-usual emissions have severe consequences for symbiotic corals. Glob. Change Biol. 2020, 26, 2203–2219. [Google Scholar] [CrossRef]
- Gordon, A.; Sprintall, J.; Van Aken, H.; Susanto, R.D.; Wijffels, S.; Molcard, R.; Ffield, A.; Pranowo, W.; Wirasantosa, S. The Indonesian throughflow during 2004–2006 as observed by the INSTANT program. Dyn. Atmos. Oceans 2010, 50, 115–128. [Google Scholar] [CrossRef]
- Taufiqurrahman, E.; Wahyudi, A.J.; Masumoto, Y. The Indonesian Throughflow and its Impact on Biogeochemistry in the Indonesian Seas. ASEAN J. Sci. Technol. Dev. 2020, 37, 29–35. [Google Scholar] [CrossRef]
- Sprintall, J.; Wijffels, S.; Molcard, R.; Jaya, I. Direct estimates of the Indonesian Throughflow entering the Indian Ocean: 2004–2006. J. Geophys. Res. Earth Surf. 2009, 114, 114. [Google Scholar] [CrossRef]
- Susanto, D.; Wei, Z.; Adi, R.; Zhang, Q.; Fang, G.; Fan, B.; Supangat, A.; Agustiadi, T.; Li, S.; Trenggono, M.; et al. Oceanography Surrounding Krakatau Volcano in the Sunda Strait, Indonesia. Oceanography 2016, 29, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Wirasatriya, A.; Susanto, R.D.; Kunarso, K.; Jalil, A.R.; Ramdani, F.; Puryajati, A.D. Northwest monsoon upwelling within the Indonesian seas. Int. J. Remote Sens. 2021, 42, 5433–5454. [Google Scholar] [CrossRef]
- RDTL. Population and Housing Census 2015 Preliminary Results; Democratic Republic of Timor-Leste (RDTL): Dili, Timor-Leste, 2015; p. 39.
- DNMG; BOM; CSIRO. Current and Future Climate of Timor-Leste; Timor-Leste National Directorate of Meteorology and Geophysics: Dili, Timor-Leste, 2015; pp. 1–8.
- United Nations Development Program; Democratic Republic of Timor-Leste. National Coastal Vulnerability Assessment and Designing of Integrated Coastal Management and Adaptation Strategic Plan for Timor-Leste; United Nations Development Programme: Dili, Timor-Leste, 2018; p. 73.
- PIFSC. Interdisciplinary Baseline Ecosystem Assessment Surveys to Inform Ecosystem-Based Management Planning in Timor-Leste: Final Report; NOAA Pacific Islands Fisheries Science Center: Honolulu, HI, USA, 2017; p. 234.
- English, S.; Wilkinson, C.; Baker, V. Survey Manual for Tropical Marine Resources; Australian Institute of Marine Science: Townsville, Australia, 1997.
- Zvuloni, A.; Armoza-Zvuloni, R.; Loya, Y. Structural deformation of branching corals associated with the vermetid gastropod Dendropoma maxima. Mar. Ecol. Prog. Ser. 2008, 363, 103–108. [Google Scholar] [CrossRef]
- The R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Anderson, M.; Gorley, R.N.; Clarke, K. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. Getting Started with PRIMER V7; PRIMER-E: Plymouth, UK, 2015. [Google Scholar]
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. 2020. Available online: https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 6 March 2022).
- Pinheiro, J.; Bates, D.; Debroy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Brown, K.; Bender-Champ, D.; Bryant, D.E.; Dove, S.; Hoegh-Guldberg, O. Human activities influence benthic community structure and the composition of the coral-algal interactions in the central Maldives. J. Exp. Mar. Biol. Ecol. 2017, 497, 33–40. [Google Scholar] [CrossRef]
- Bruno, J.F.; Valdivia, A. Coral reef degradation is not correlated with local human population density. Sci. Rep. 2016, 6, 29778. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.E.; Brainard, R.; Carter, A.; Grillo, S.; Edwards, C.; Harris, J.; Lewis, L.; Obura, D.; Rohwer, F.; Sala, E.; et al. Re-evaluating the health of coral reef communities: Baselines and evidence for human impacts across the central Pacific. Proc. R. Soc. B Boil. Sci. 2016, 283, 20151985. [Google Scholar] [CrossRef]
- Wedding, L.M.; Lecky, J.; Gove, J.M.; Walecka, H.R.; Donovan, M.K.; Williams, G.J.; Jouffray, J.-B.; Crowder, L.B.; Erickson, A.; Falinski, K.; et al. Advancing the integration of spatial data to map human and natural drivers on coral reefs. PLoS ONE 2018, 13, e0189792. [Google Scholar] [CrossRef] [Green Version]
- Erdmann, M.V.; Mohan, C. (Eds.) A Rapid Marine Biological Assessment of Timor-Leste, RAP Bulletin of Biological Assessment 66; Coral Triangle Support Partnership, Conservation International: Dili, Timor-Leste, 2013; ISBN 978-1-934151-56-3. [Google Scholar]
- Boggs, G.; Edyvane, K.; de Carvalho, N.; Penny, S.; Rouwenhorst, J.; Brocklehurst, P.; Cowie, I.; Barreto, C.; Amaral, A.; Monteiro, J.; et al. Marine and Coastal Habitat Mapping in Timor Leste (North Coast)—Final Report; Ministry of Agriculture & Fisheries, Government of Timor Leste: Dili, Timor-Leste, 2012.
- Kim, C.J.S. Drivers of Coral Reef Composition, Cryptic Marine Biodiversity, and Coral Health along the North Coast of Timor-Leste. Ph.D. Thesis, The University of Queensland, St. Lucia, Australia, 2021. [Google Scholar]
- Teh, L.S.L.; Teh, L.C.L.; Sumaila, U.R. A Global Estimate of the Number of Coral Reef Fishers. PLoS ONE 2013, 8, e65397. [Google Scholar] [CrossRef]
- Da Costa, M.D.; Lopes, M.; Ximenes, A.; Ferreira, A.D.R.; Spyckerelle, L.; Williams, R.; Nesbitt, H.; Erskine, W. Household food insecurity in Timor-Leste. Food Secur. 2013, 5, 83–94. [Google Scholar] [CrossRef]
- Cesar, H.; Burke, L.; Pet-Soede, L. The Economics of Worldwide Coral Reef Degradation; Cesar Environmental Economics Consulting, Arnhem, and WWF-Netherlands: Arnhem, The Netherlands, 2003; p. 23. [Google Scholar]
- Gillett, R. Fisheries in the Economies of the Pacific Island Countries and Territories; Asia Development Bank: Mandaluyong, Philippines, 2009. [Google Scholar]
- Andréfouët, S.; Guillaume, M.M.M.; Delval, A.; Rasoamanendrika, F.M.A.; Blanchot, J.; Bruggemann, J.H. Fifty years of changes in reef flat habitats of the Grand Récif of Toliara (SW Madagascar) and the impact of gleaning. Coral Reefs 2013, 32, 757–768. [Google Scholar] [CrossRef]
- Ashworth, J.S.; Ormond, R.F.; Sturrock, H.T. Effects of reef-top gathering and fishing on invertebrate abundance across take and no-take zones. J. Exp. Mar. Biol. Ecol. 2004, 303, 221–242. [Google Scholar] [CrossRef]
- Woodland, D.; Hooper, J. The effect of human trampling on coral reefs. Biol. Conserv. 1977, 11, 1–4. [Google Scholar] [CrossRef]
- Ayling, A.M.; Ayling, A.L.; Edyvane, K.S.; Penny, S.; de Carvalho, N.; Fernandes, A.; Amaral, A.L. Preliminary Biological Resource Survey of Fringing Reefs in the Proposed Nino Konis Santana Marine Park, Timor-Leste. Rep. North. Territ. Dep. Nat. Resour. Environ. Arts. Palmerst. North. Territ. 2009, 830. [Google Scholar]
- Maynard, J.A.; Van Hooidonk, R.; Eakin, C.M.; Puotinen, M.; Garren, M.; Williams, G.J.; Heron, S.; Lamb, J.; Weil, E.; Willis, B.L.; et al. Projections of climate conditions that increase coral disease susceptibility and pathogen abundance and virulence. Nat. Clim. Change 2015, 5, 688–694. [Google Scholar] [CrossRef]
- Amaro, C.; Biosca, E.G. Vibrio vulnificus biotype 2, pathogenic for eels, is also an opportunistic pathogen for humans. Appl. Environ. Microbiol. 1996, 62, 1454–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervino, J.M.; Hayes, R.L.; Polson, S.W.; Polson, S.C.; Goreau, T.J.; Martinez, R.J.; Smith, G.W. Relationship of Vibrio Species Infection and Elevated Temperatures to Yellow Blotch/Band Disease in Caribbean Corals. Appl. Environ. Microbiol. 2004, 70, 6855–6864. [Google Scholar] [CrossRef] [Green Version]
- Linkous, D.A.; Oliver, J.D. Pathogenesis ofVibrio vulnificus. FEMS Microbiol. Lett. 1999, 174, 207–214. [Google Scholar] [CrossRef]
- Milton, D.L.; Norqvist, A.; Wolf-Watz, H. Cloning of a metalloprotease gene involved in the virulence mechanism of Vibrio anguillarum. J. Bacteriol. 1992, 174, 7235–7244. [Google Scholar] [CrossRef] [Green Version]
- Sussman, M.; Willis, B.L.; Victor, S.; Bourne, D.G. Coral Pathogens Identified for White Syndrome (WS) Epizootics in the Indo-Pacific. PLoS ONE 2008, 3, e2393. [Google Scholar] [CrossRef] [Green Version]
- Ushijima, B.; Videau, P.; Burger, A.H.; Shore, A.; Runyon, C.M.; Sudek, M.; Aeby, G.S.; Callahan, S.M. Vibrio coralliilyticus Strain OCN008 Is an Etiological Agent of Acute Montipora White Syndrome. Appl. Environ. Microbiol. 2014, 80, 2102–2109. [Google Scholar] [CrossRef] [Green Version]
- Ushijima, B.; Smith, A.; Aeby, G.S.; Callahan, S.M. Vibrio owensii Induces the Tissue Loss Disease Montipora White Syndrome in the Hawaiian Reef Coral Montipora capitata. PLoS ONE 2012, 7, e46717. [Google Scholar] [CrossRef] [Green Version]
- Aeby, G.S.; Ross, M.; Williams, G.J.; Lewis, T.D.; Work, T.M. Disease dynamics of Montipora white syndrome within Kaneohe Bay, Oahu, Hawaii: Distribution, seasonality, virulence, and transmissibility. Dis. Aquat. Org. 2010, 91, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haapkylä, J.; Unsworth, R.; Seymour, A.; Melbourne-Thomas, J.; Flavell, M.; Willis, B.; Smith, D. Spatio-temporal coral disease dynamics in the Wakatobi Marine National Park, South-East Sulawesi, Indonesia. Dis. Aquat. Org. 2009, 87, 105–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aeby, G.S. Outbreak of coral disease in the Northwestern Hawaiian Islands. Coral Reefs 2005, 24, 481. [Google Scholar] [CrossRef]
- Roff, G.; Hoegh-Guldberg, O.; Fine, M. Intra-colonial response to Acroporid “white syndrome” lesions in tabular Acropora spp. (Scleractinia). Coral Reefs 2006, 25, 255–264. [Google Scholar] [CrossRef]
- Haapkylä, J.; Seymour, A.; Trebilco, J.; Smith, D. Coral disease prevalence and coral health in the Wakatobi Marine Park, south-east Sulawesi, Indonesia. J. Mar. Biol. Assoc. UK 2007, 87, 403–414. [Google Scholar] [CrossRef]
- Johan, O.; Bengen, D.G.; Zamani, N.P.; Suharsono; Sweet, M. The Distribution and Abundance of Black Band Disease and White Syndrome in Kepulauan Seribu, Indonesia. HAYATI J. Biosci. 2015, 22, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Muller, E.M.; Raymundo, L.J.; Willis, B.L.; Haapkylä, J.; Yusuf, S.; Wilson, J.R.; Harvell, D.C. Coral Health and Disease in the Spermonde Archipelago and Wakatobi, Sulawesi. J. Indones. Coral Reefs 2012, 1, 147–159. [Google Scholar]
- Muller, E.M.; Van Woesik, R. Caribbean coral diseases: Primary transmission or secondary infection? Glob. Chang. Biol. 2012, 18, 3529–3535. [Google Scholar] [CrossRef]
- Aeby, G.S.; Bourne, D.G.; Wilson, B.; Work, T. Coral Diversity and the Severity of Disease Outbreaks: A Cross-Regional Comparison ofAcroporaWhite Syndrome in a Species-Rich Region (American Samoa) with a Species-Poor Region (Northwestern Hawaiian Islands). J. Mar. Biol. 2011, 2011, 490198. [Google Scholar] [CrossRef] [Green Version]
- Elton, C.S. The Biology of Invasions by Animals and Plants; John Wiley and Sons: New York, NY, USA, 1958. [Google Scholar]
- Plank, J.E. Plant Diseases-Epidemics and Control; Academic Press: New York, NY, USA, 1963. [Google Scholar]
- Hobbs, J.-P.A.; Frisch, A.J.; Newman, S.; Wakefield, C.B. Selective Impact of Disease on Coral Communities: Outbreak of White Syndrome Causes Significant Total Mortality of Acropora Plate Corals. PLoS ONE 2015, 10, e0132528. [Google Scholar] [CrossRef] [Green Version]
- Sheridan, C.; Grosjean, P.; Leblud, J.; Palmer, C.V.; Kushmaro, A.; Eeckhaut, I. Sedimentation rapidly induces an immune response and depletes energy stores in a hard coral. Coral Reefs 2014, 33, 1067–1076. [Google Scholar] [CrossRef]
- Heron, S.F.; Willis, B.L.; Skirving, W.J.; Eakin, C.M.; Page, C.A.; Miller, I.R. Summer Hot Snaps and Winter Conditions: Modelling White Syndrome Outbreaks on Great Barrier Reef Corals. PLoS ONE 2010, 5, e12210. [Google Scholar] [CrossRef] [Green Version]
- Altizer, S.; Ostfeld, R.S.; Johnson, P.T.J.; Kutz, S.; Harvell, C.D. Climate Change and Infectious Diseases: From Evidence to a Predictive Framework. Science 2013, 341, 514–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alongi, D.M.; da Silva, M.; Wasson, R.J.; Wirasantosa, S. Sediment discharge and export of fluvial carbon and nutrients into the Arafura and Timor Seas: A regional synthesis. Mar. Geol. 2013, 343, 146–158. [Google Scholar] [CrossRef]
- Barneah, O.; Brickner, I.; Hooge, M.; Weis, V.; LaJeunesse, T.; Benayahu, Y. Three party symbiosis: Acoelomorph worms, corals and unicellular algal symbionts in Eilat (Red Sea). Mar. Biol. 2007, 151, 1215–1223. [Google Scholar] [CrossRef]
- Naumann, M.S.; Mayr, C.; Struck, U.; Wild, C. Coral mucus stable isotope composition and labeling: Experimental evidence for mucus uptake by epizoic acoelomorph worms. Mar. Biol. 2010, 157, 2521–2531. [Google Scholar] [CrossRef]
- Wijgerde, T.; Schots, P.; Van Onselen, E.; Janse, M.; Karruppannan, E.; Verreth, J.; Osinga, R. Epizoic acoelomorph flatworms impair zooplankton feeding by the scleractinian coral Galaxea fascicularis. Biol. Open 2013, 2, 10–17. [Google Scholar] [CrossRef] [Green Version]
- McClanahan, T. Primary succession of coral-reef algae: Differing patterns on fished versus unfished reefs. J. Exp. Mar. Biol. Ecol. 1997, 218, 77–102. [Google Scholar] [CrossRef]
- Vermeij, M.J.A.; Van Moorselaar, I.; Engelhard, S.; Hörnlein, C.; Vonk, S.M.; Visser, P.M. The Effects of Nutrient Enrichment and Herbivore Abundance on the Ability of Turf Algae to Overgrow Coral in the Caribbean. PLoS ONE 2010, 5, e14312. [Google Scholar] [CrossRef] [Green Version]
- Leichter, J.J.; Stewart, H.L.; Miller, S.L. Episodic nutrient transport to Florida coral reefs. Limnol. Oceanogr. 2003, 48, 1394–1407. [Google Scholar] [CrossRef] [Green Version]
- Risk, M.J.; Lapointe, B.; Sherwood, O.A.; Bedford, B.J. The use of Δ15N in assessing sewage stress on coral reefs. Mar. Pollut. Bull. 2009, 58, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Burgett, J. Summary of Algal Community Changes Observed on the Southwest Arm of Rose Atoll from 1995–2002; USFWS: Honolulu, HI, USA, 2012.
- Thacker, R.; Ginsburg, D.; Paul, V. Effects of herbivore exclusion and nutrient enrichment on coral reef macroalgae and cyanobacteria. Coral Reefs 2001, 19, 318–329. [Google Scholar] [CrossRef]
- Littler, M.M.; Littler, D.S.; Brooks, B. Harmful algae on tropical coral reefs: Bottom-up eutrophication and top-down herbivory. Harmful Algae 2006, 5, 565–585. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Kiyashko, S.I.; Titlyanova, T.V.; Van Huyen, P.; Yakovleva, I.M. Identifying nitrogen sources for macroalgal growth in variously polluted coastal areas of southern Vietnam. Bot. Mar. 2011, 54, 367–376. [Google Scholar] [CrossRef]
- Costanzo, S.D.; Udy, J.; Longstaff, B.; Jones, A. Using nitrogen stable isotope ratios (Δ15N) of macroalgae to determine the effectiveness of sewage upgrades: Changes in the extent of sewage plumes over four years in Moreton Bay, Australia. Mar. Pollut. Bull. 2005, 51, 212–217. [Google Scholar] [CrossRef]
- Thornber, C.S.; Kinlan, B.; Graham, M.H.; Stachowicz, J.J. Population ecology of the invasive kelp Undaria pinnatifida in California: Environmental and biological controls on demography. Mar. Ecol. Prog. Ser. 2004, 268, 69–80. [Google Scholar] [CrossRef]
- Smith, J.; Runcie, J.; Smith, C. Characterization of a large-scale ephemeral bloom of the green alga Cladophora sericea on the coral reefs of West Maui, Hawai’i. Mar. Ecol. Prog. Ser. 2005, 302, 77–91. [Google Scholar] [CrossRef]
- Fourqurean, J.W.; Moore, T.O.; Fry, B.; Hollibaugh, J.T. Spatial and Temporal Variation in C:N:P Ratios, Δ15N, and Δ13C of Eelgrass Zostera Marina as Indicators of Ecosystem Processes, Tomales Bay, California, USA. Mar. Ecol. Prog. Ser. 1997, 157, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Rauenzahn, J.L.; Heron, S.; Eakin, C.M.; Skirving, W.J.; Christensen, T.R.L.; Strong, A.E.; Li, J. NOAA Coral Reef Watch 50 Km Satellite Sea Surface Temperature-Based Decision Support System for Coral Bleaching Management; NOAA/NERDIS: College Park, MD, USA, 2013.
- Sprintall, J.; Gordon, A.L.; Wijffels, S.; Feng, M.; Hu, S.; Koch-Larrouy, A.; Phillips, H.; Nugroho, D.; Napitu, A.; Pujiana, K.; et al. Detecting Change in the Indonesian Seas. Front. Mar. Sci. 2019, 6, 257. [Google Scholar] [CrossRef]
- Shinoda, T.; Han, W.; Metzger, E.J.; Hurlburt, H.E. Seasonal Variation of the Indonesian Throughflow in Makassar Strait. J. Phys. Oceanogr. 2012, 42, 1099–1123. [Google Scholar] [CrossRef]
- Wirasatriya, A.; Prasetyawan, I.B.; Triyono, C.D.; Muslim; Maslukah, L. Effect of ENSO on the variability of SST and Chlorophyll-a in Java Sea. IOP Conf. Ser. Earth Environ. Sci. 2018, 116, 012063. [Google Scholar] [CrossRef] [Green Version]
- Dewi, Y.W.; Wirasatriya, A.; Sugianto, D.N.; Helmi, M.; Marwoto, J.; Maslukah, L. Effect of ENSO and IOD on the Variability of Sea Surface Temperature (SST) in Java Sea. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020; Volume 530, p. 012007. [Google Scholar]
- Randall, C.J.; Toth, L.T.; Leichter, J.J.; Maté, J.L.; Aronson, R.B. Upwelling buffers climate change impacts on coral reefs of the eastern tropical Pacific. Ecology 2020, 101, e02918. [Google Scholar] [CrossRef] [PubMed]
- Glynn, P.W.; Maté, J.L.; Baker, A.C.; Calderón, M.O. Coral Bleaching and Mortality in Panama and Ecuador during the 1997–1998 El Niño–Southern Oscillation Event: Spatial/Temporal Patterns and Comparisons with the 1982–1983 Event. Bull. Mar. Sci. 2001, 69, 79–109. [Google Scholar]
- Feely, R.A.; Sabine, C.L.; Hernandez-Ayon, J.M.; Ianson, D.; Hales, B. Evidence for Upwelling of Corrosive "Acidified" Water onto the Continental Shelf. Science 2008, 320, 1490–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leichter, J.; Genovese, S. Intermittent upwelling and subsidized growth of the scleractinian coral Madracis mirabilis on the deep fore-reef slope of Discovery Bay, Jamaica. Mar. Ecol. Prog. Ser. 2006, 316, 95–103. [Google Scholar] [CrossRef]
- Rixen, T.; Jiménez, C.; Cortés, J. Impact of upwelling events on the sea water carbonate chemistry and dissolved oxygen concentration in the Gulf of Papagayo (Culebra Bay), Costa Rica: Implications for coral reefs. RBT 2015, 60, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.T.; Cai, W.; Jin, F.-F.; Santoso, A.; Wu, L.; Guilyardi, E.; An, S.-I. Response of El Niño sea surface temperature variability to greenhouse warming. Nat. Clim. Change 2014, 4, 786–790. [Google Scholar] [CrossRef]
- Cai, W.; Wang, G.; Dewitte, B.; Wu, L.; Santoso, A.; Takahashi, K.; Yang, Y.; Carréric, A.; McPhaden, M.J. Increased variability of eastern Pacific El Niño under greenhouse warming. Nature 2018, 564, 201–206. [Google Scholar] [CrossRef]
- Beyer, H.L.; Kennedy, E.V.; Beger, M.; Chen, C.A.; Cinner, J.E.; Darling, E.S.; Eakin, C.M.; Gates, R.D.; Heron, S.F.; Knowlton, N.; et al. Risk-sensitive planning for conserving coral reefs under rapid climate change. Conserv. Lett. 2018, 11, e12587. [Google Scholar] [CrossRef]
- Kim, C.J. Coral Reef Management and Tara Bandu on Ataúro Island: An Ecologist’s Perspective. In Proceedings of the 2020 Timor-Leste Studies Association-Portugal Conference, TLSA-PT, Dili, Timor-Leste, 8 April 2021. [Google Scholar]
- Lamb, J.B.; Willis, B.L.; Fiorenza, E.A.; Couch, C.S.; Howard, R.; Rader, D.N.; True, J.D.; Kelly, L.A.; Ahmad, A.; Jompa, J.; et al. Plastic waste associated with disease on coral reefs. Science 2018, 359, 460–462. [Google Scholar] [CrossRef]
- Lamb, J.B.; van de Water, J.A.J.M.; Bourne, D.G.; Altier, C.; Hein, M.Y.; Fiorenza, E.A.; Abu, N.; Jompa, J.; Harvell, C.D. Seagrass ecosystems reduce exposure to bacterial pathogens of humans, fishes, and invertebrates. Science 2017, 355, 731–733. [Google Scholar] [CrossRef] [PubMed]
- Sabdono, A.; Radjasa, O.K.; Trianto, A.; Sarjito; Munasik; Wijayanti, D.P. Preliminary study of the effect of nutrient enrichment, released by marine floating cages, on the coral disease outbreak in Ka-rimunjawa, Indonesia. Reg. Stud. Mar. Sci. 2019, 30, 100704. [Google Scholar] [CrossRef] [Green Version]
- Raymundo, L.J.; Licuanan, W.L.; Kerr, A.M. Adding insult to injury: Ship groundings are associated with coral disease in a pristine reef. PLoS ONE 2018, 13, e0202939. [Google Scholar] [CrossRef] [Green Version]
- Raymundo, L.J.; Diaz, R.; Miller, A.; Reynolds, T. Baseline Surveys of Proposed and Established Marine Sanctuaries on Bantayan Island, Northern Cebu; University of Guam: Mangilao, Guam, 2011; p. 64. [Google Scholar]
- Kaczmarsky, L. Coral disease dynamics in the central Philippines. Dis. Aquat. Org. 2006, 69, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymundo, L.J.; Rosell, K.B.; Reboton, C.T.; Kaczmarsky, L. Coral diseases on Philippine reefs: Genus Porites is a dominant host. Dis. Aquat. Org. 2005, 64, 181–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymundo, L. Porites ulcerative white spot disease: Description, prevalence, and host range of a new coral disease affecting Indo-Pacific reefs. Dis. Aquat. Org. 2003, 56, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.; Sweet, M.; Wood, E.; Bythell, J. Baseline coral disease surveys within three marine parks in Sabah, Borneo. PeerJ 2015, 3, 1391. [Google Scholar] [CrossRef] [Green Version]
- Green, E.P.; Bruckner, A.W. The significance of coral disease epizootiology for coral reef conservation. Biol. Conserv. 2000, 96, 347–361. [Google Scholar] [CrossRef]
- Williams, G.J.; Work, T.M.; Aeby, G.S.; Knapp, I.S.; Davy, S.K. Gross and microscopic morphology of lesions in Cnidaria from Palmyra Atoll, Central Pacific. J. Invertebr. Pathol. 2011, 106, 165–173. [Google Scholar] [CrossRef]
- Yasuda, N.; Nakano, Y.; Yamashiro, H.; Hidaka, M. Skeletal structure and progression of growth anomalies in Porites australiensis in Okinawa, Japan. Dis. Aquat. Org. 2012, 97, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Aeby, G.S. The Potential Effect on the Ability of a Coral Intermediate Host to Regenerate Has Had on the Evolution of Its Association with a Marine Parasite. In Proceedings of the 7th International Coral Reef Symposium, Guam, FSM, USA, 22–27 June 1992; pp. 809–815. [Google Scholar]
- Aeby, G.S. Corals in the genus Porites are susceptible to infection by a larval trematode. Coral Reefs 2003, 22, 216. [Google Scholar] [CrossRef]
- Bergsma, G.S. Tube-dwelling coral symbionts induce significant morphological change in Montipora. Symbiosis 2009, 49, 143–150. [Google Scholar] [CrossRef]
- Floros, C.; Samways, M.; Armstrong, B. Polychaete (Spirobranchus giganteus) loading on South African corals. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 289–298. [Google Scholar] [CrossRef]
- Scott, P.J.B.; Risk, M.J. The effect of Lithophaga (Bivalvia: Mytilidae) boreholes on the strength of the coral Porites lobata. Coral Reefs 1988, 7, 145–151. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Farenzena, Z.T. Tissue loss in corals infested by acoelomorph flatworms (Waminoa sp.). Coral Reefs 2012, 31, 869. [Google Scholar] [CrossRef] [Green Version]
- Haapkylä, J.; Seymour, A.S.; Barneah, O.; Brickner, I.; Hennige, S.; Suggett, D.; Smith, D. Association of Waminoa sp. (Acoela) with corals in the Wakatobi Marine Park, South-East Sulawesi, Indonesia. Mar. Biol. 2009, 156, 1021–1027. [Google Scholar] [CrossRef]
- Rodríguez-Villalobos, J.; Work, T.; Calderón-Aguilera, L.; Reyes-Bonilla, H.; Hernández, L. Explained and unexplained tissue loss in corals from the Tropical Eastern Pacific. Dis. Aquat. Org. 2015, 116, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Keats, D.W.; Chamberlain, Y.M.; Baba, M. Pneophyllum conicum (Dawson) comb. nov. (Rhodophyta, Corallinaceae), a Widespread Indo-Pacific Non-Geniculate Coralline Alga that Overgrows and Kills Live Coral. Bot. Mar. 1997, 40, 263–280. [Google Scholar] [CrossRef]
- Finckh, A.E. Biology of the Reef-Forming Organisms at Funafuti Atoll. In The Atoll of Funafuti; Royal Society: London, UK, 1904; pp. 125–150. [Google Scholar]
- Smith, T.; Nemeth, R.; Blondeau, J.; Calnan, J.; Kadison, E.; Herzlieb, S. Assessing coral reef health across onshore to offshore stress gradients in the US Virgin Islands. Mar. Pollut. Bull. 2008, 56, 1983–1991. [Google Scholar] [CrossRef]
- Kuffner, I.B.; Walters, L.J.; Becerro, M.A.; Paul, V.J.; Ritson-Williams, R.; Beach, K.S. Inhibition of coral recruitment by macroalgae and cyanobacteria. Mar. Ecol. Prog. Ser. 2006, 323, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Cetz-Navarro, N.P.; Espinoza-Avalos, J.; Hernández-Arana, H.A.; Carricart-Ganivet, J.P. Biological Responses of the Coral Montastraea annularis to the Removal of Filamentous Turf Algae. PLoS ONE 2013, 8, e54810. [Google Scholar] [CrossRef] [Green Version]
- Loh, T.-L.; Pawlik, J. Friend or foe? No evidence that association with the sponge Mycale laevis provides a benefit to corals of the genus Montastraea. Mar. Ecol. Prog. Ser. 2012, 465, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Rützler, K.; Muzik, K. Terpios Hoshinota, a New Cyanobacteriosponge Threatening Pacific Reefs. Sci. Mar. 1993, 57, 395–403. [Google Scholar]
- Bak, R.; Lambrechts, D.; Joenje, M.; Nieuwland, G.; Van Veghel, M. Long-term changes on coral reefs in booming populations of a competitive colonial ascidian. Mar. Ecol. Prog. Ser. 1996, 133, 303–306. [Google Scholar] [CrossRef]
- Littler, M.M.; Littler, D.S. A Colonial Tunicate Smothers Corals and Corraline Algae on the Great Astrolabe Reef, Fiji. Coral Reefs 1995, 14, 148–149. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Ángel, B.; Godwin, L.S.; Asher, J.; Brainard, R.E. Invasive didemnid tunicate spreading across coral reefs at remote Swains Island, American Sāmoa. Coral Reefs 2009, 28, 53. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, J.; Raghukumar, C. Pink-line syndrome, a physiological crisis in the scleractinian coral Porites lutea. Mar. Biol. 2006, 149, 347–356. [Google Scholar] [CrossRef]
- Eyre, B. Nutrient Biogeochemistry in the Tropical Moresby River Estuary System North Queensland, Australia. Estuar. Coast. Shelf Sci. 1994, 39, 15–31. [Google Scholar] [CrossRef]
- Mitchell, A.W.; Bramley, R.; Johnson, A.K.L. Export of nutrients and suspended sediment during a cyclone-mediated flood event in the Herbert River catchment, Australia. Mar. Freshw. Res. 1997, 48, 79–88. [Google Scholar] [CrossRef]
- Robertson, A.I.; Dixon, P.; Alongi, D.M. The Influence of Fluvial Discharge on Pelagic Production in the Gulf of Papua, Northern Coral Sea. Estuar. Coast. Shelf Sci. 1998, 46, 319–331. [Google Scholar] [CrossRef]
- Alongi, D.M.; Boto, K.G.; Robertson, A.I. Nitrogen and phosphorus cycles. In Ecosystems at the Land-Sea Margin: Drainage Basin to Coastal Sea; Robertson, A.I., Alongi, D.M., Eds.; American Geophysical Union: Washington, DC, USA, 1992; Volume 41, pp. 251–292. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algae | Effect | df | F-Value | p-Value | Rural-E | Rural-N | Urban-E | Urban-W |
|---|---|---|---|---|---|---|---|---|
| Halimeda spp. | Site | 3 | 3.8199 | 0.0121 * | 4.26‰ | 4.31‰ | 4.03‰ | 4.26‰ |
| Depth | 1 | 0.5442 | 0.4624 | ±0.06 | ±0.07 | ±0.06 | ±0.07 | |
| Site x Depth | 3 | 1.3801 | 0.2531 | ab | b | a | b | |
| Chlorodesmis spp. | Site | 1 | 10.0028 | 0.0064 * | 4.57‰ | - | 4.11‰ | 4.47‰ |
| Depth | 1 | 0.1747 | 0.6819 | ±0.15 | ±0.08 | ±0.10 | ||
| Site x Depth | 1 | 2.4127 | 0.1412 | ab | a | b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.J.S.; Roelfsema, C.; Dove, S.; Hoegh-Guldberg, O. The Condition of Four Coral Reefs in Timor-Leste before and after the 2016–2017 Marine Heatwave. Oceans 2022, 3, 147-171. https://doi.org/10.3390/oceans3020012
Kim CJS, Roelfsema C, Dove S, Hoegh-Guldberg O. The Condition of Four Coral Reefs in Timor-Leste before and after the 2016–2017 Marine Heatwave. Oceans. 2022; 3(2):147-171. https://doi.org/10.3390/oceans3020012
Chicago/Turabian StyleKim, Catherine J. S., Chris Roelfsema, Sophie Dove, and Ove Hoegh-Guldberg. 2022. "The Condition of Four Coral Reefs in Timor-Leste before and after the 2016–2017 Marine Heatwave" Oceans 3, no. 2: 147-171. https://doi.org/10.3390/oceans3020012
APA StyleKim, C. J. S., Roelfsema, C., Dove, S., & Hoegh-Guldberg, O. (2022). The Condition of Four Coral Reefs in Timor-Leste before and after the 2016–2017 Marine Heatwave. Oceans, 3(2), 147-171. https://doi.org/10.3390/oceans3020012