

Age and Sexual Maturity Estimation of Stranded Striped Dolphins, Stenella coeruleoalba, Infected with Brucella ceti

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Dolphin Population
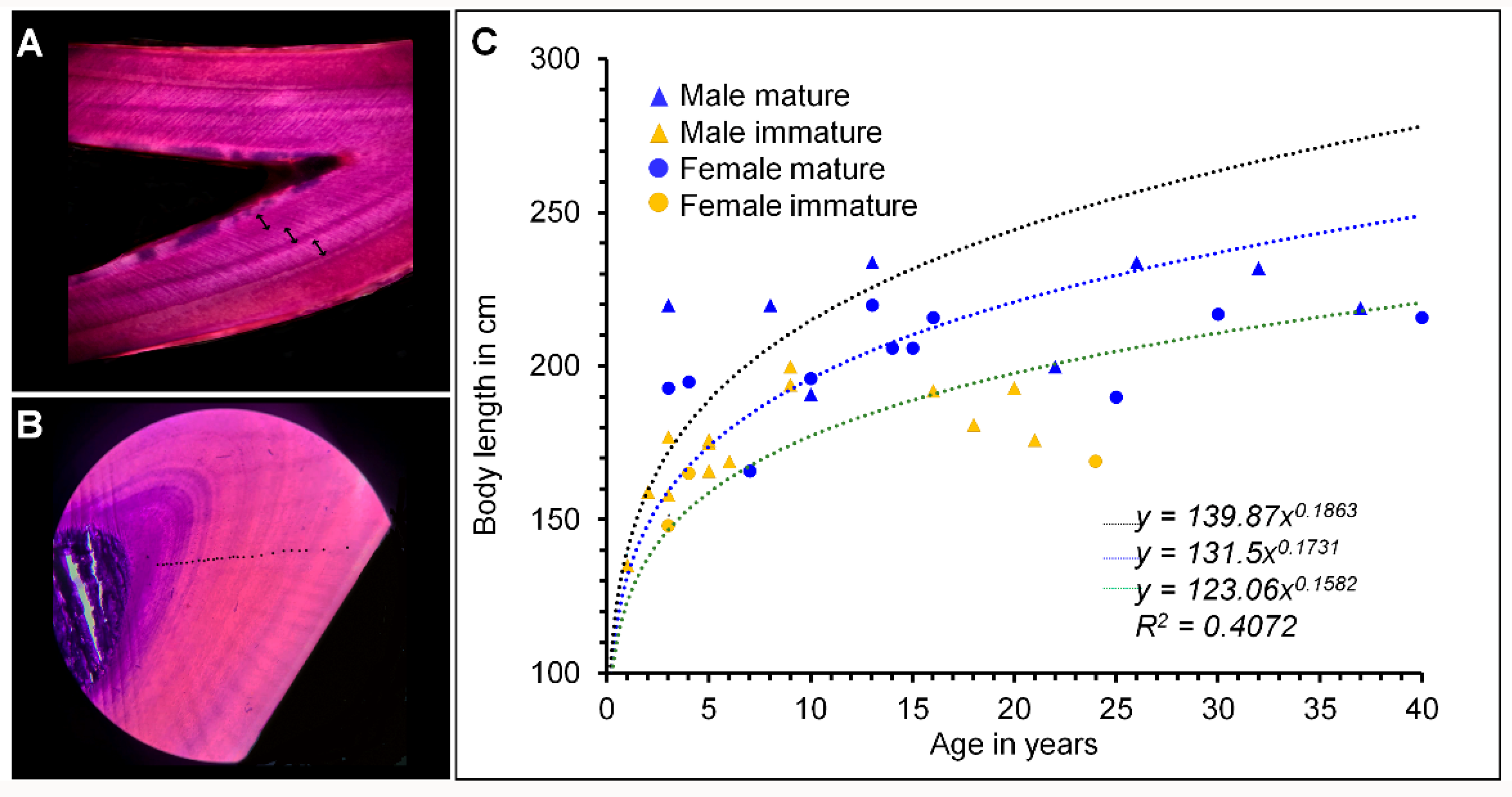
2.2. Growth Layer Groups (GLG) of Dentin
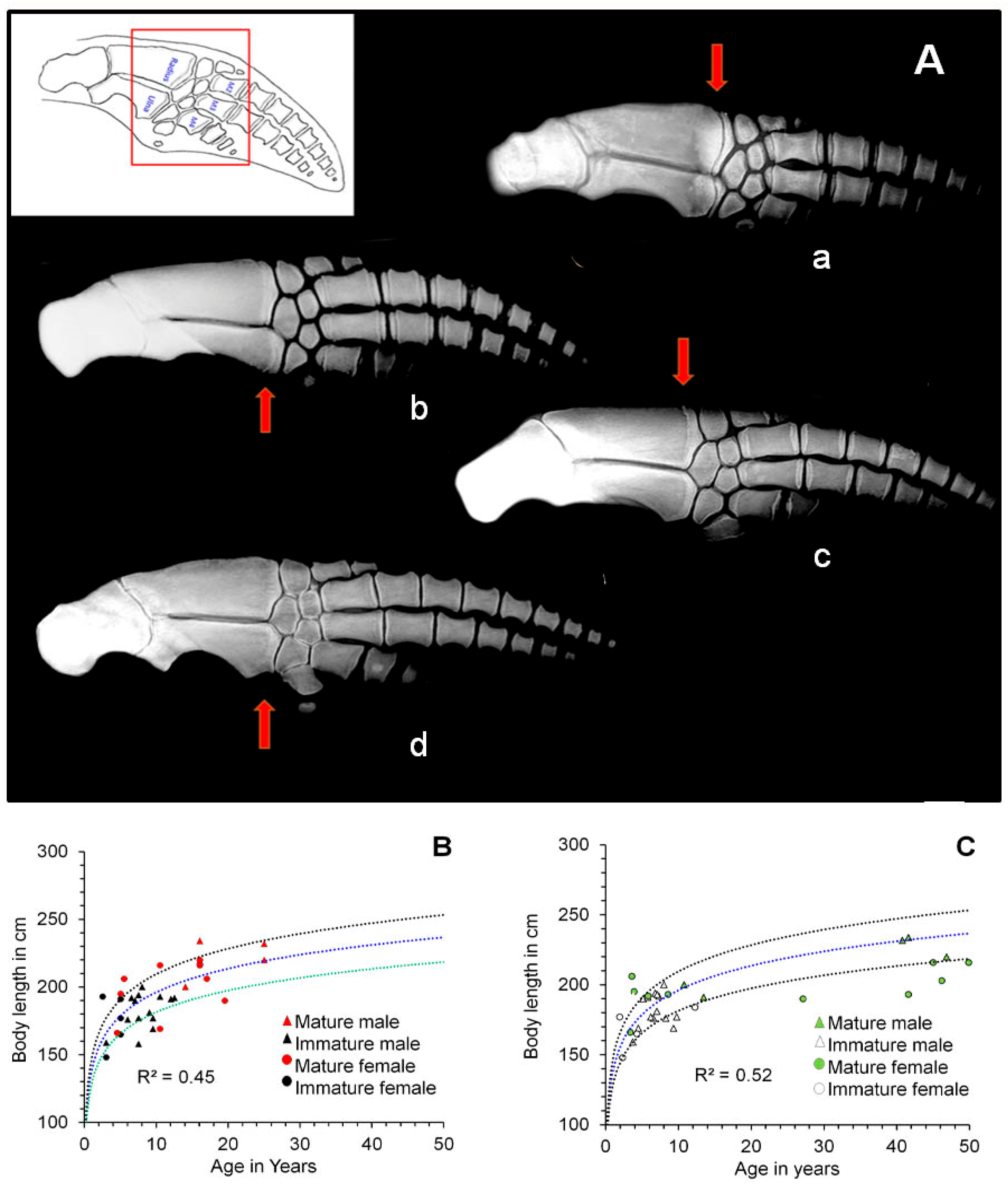
2.3. Bone Ossification
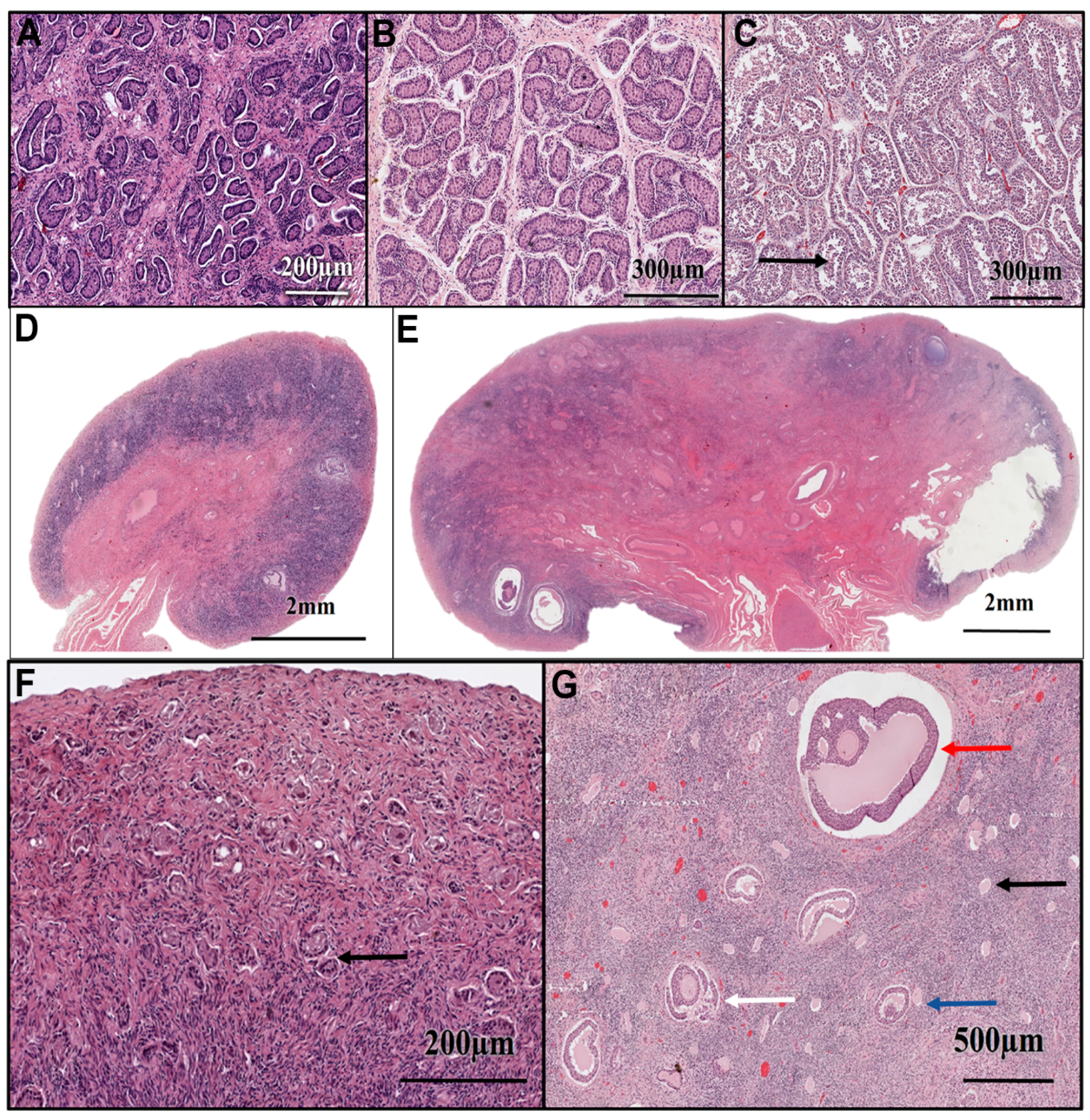
2.4. Gonadal Histology
2.5. Hormonal Levels
2.6. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Archer, F.I. Morphological and Genetic Variation of Striped Dolphins (Stenella coeruleoalba); University of California: San Diego, CA, USA, 1996. [Google Scholar]
- Archer, F.I.; Perrin, W.F. Stenella coeruleoalba. Mamm. Species 1999, 603, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Archer, F.I. Striped Dolphin: Stenella coeruleoalba. In Encyclopedia of Marine Mammals, 3rd ed.; Würsig, B., Thewissen, J.G.M., Kovacs, K.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 954–956. ISBN 9780128043271. [Google Scholar]
- Wursig, B.; Perrin, W.F.; Thewissen, J.G.M. Encyclopedia of Marine Mammals, 2nd ed.; Academic Press: Cambridge, MA, USA, 2009; ISBN 9780123735539. [Google Scholar]
- Hammond, P.S.; Bearzi, G.; Bjørge, A.; Forney, K.; Kaczmarski, L.; Kasuya, T.; Perrin, W.F.; Scott, M.; Wang, J.; Wells, R.S.; et al. Stenella coeruleoalba. IUCN Red List Threat. Species 2008, e.T20731A50374282. [Google Scholar] [CrossRef]
- Miyazaki, N. Growth and reproduction of Stenella coeruleoalba off the Pacific coast of Japan. Sci. Rep. Whales Res. Inst. 1977, 29, 1–48. [Google Scholar]
- Calzada, N.; Aguilar, A. Flipper development in the Mediterranean striped dolphin (Stenella coeruleoalba). Anat. Rec. 1996, 245, 708–714. [Google Scholar] [CrossRef]
- González-Barrientos, R.; Morales, J.A.; Hernández-Mora, G.; Barquero-Calvo, E.; Guzmán-Verri, C.; Chaves-Olarte, E.; Moreno, E. Pathology of striped dolphins (Stenella coeruleoalba) infected with Brucella ceti. J. Comp. Pathol. 2010, 142, 347–352. [Google Scholar] [CrossRef]
- Granados-Zapata, A.; Robles-Malagamba, M.J.; González-Barrientos, R.; Kot, B.C.-W.; Barquero-Calvo, E.; Cordero-Chavaría, M.; Suárez-Esquivel, M.; Guzmán-Verri, C.; Palacios-Alfaro, J.D.; Tien-Sung, C.; et al. Pathological studies and postmortem computed tomography of dolphins with meningoencephalomyelitis and osteoarthritis caused by Brucella ceti. Oceans 2022, 3, 189–203. [Google Scholar] [CrossRef]
- Guzmán-Verri, C.; González-Barrientos, R.; Hernández-Mora, G.; Morales, J.-A.; Baquero-Calvo, E.; Chaves-Olarte, E.; Moreno, E. Brucella ceti and brucellosis in cetaceans. Front. Cell. Infect. Microbiol. 2012, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Mora, G.; González-Barrientos, R.; Morales, J.A.; Chaves-Olarte, E.; Guzmán-Verri, C.; Baquero-Calvo, E.; De-Miguel, M.J.; Marín, C.M.; Blasco, J.M.; Moreno, E. Neurobrucellosis in stranded dolphins, Costa Rica. Emerg. Infect. Dis. 2008, 14, 1430–1433. [Google Scholar] [CrossRef]
- Hernández-Mora, G.; Palacios-Alfaro, J.D.; González-Barrientos, R. Wildlife reservoirs of brucellosis: Brucella in aquatic environments. Rev.-Off. Int. Epizoot. 2013, 32, 89–103. [Google Scholar] [CrossRef]
- Hernández-Mora, G.; González-Barrientos, R.; Víquez-Ruíz, E.; Palacios-Alfaro, J.D.; Bettoni-Rodríguez, G.; Gendre, M.; Vincent, C.; Roca-Monge, K.; Ruiz-Villalobos, N.; Suárez-Esquivel, M.; et al. Brucella sp. sequence-type 27 associated with abortion in dwarf sperm whale Kogia sima. Eur. J. Wildl. Res. 2021, 67, 63. [Google Scholar] [CrossRef]
- Buckle, K.; Roe, W.D.; Howe, L.; Michael, S.; Duignan, P.J.; Burrows, E.; Ha, H.J.; Humphrey, S.; McDonald, W.L. Brucellosis in endangered Hector’s dolphins (Cephalorhynchus hectori). Vet. Pathol. 2017, 54, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Colegrove, K.M.; Venn-Watson, S.; Litz, J.; Kinsel, M.J.; Terio, K.A.; Fougeres, E.; Ewing, R.; Pabst, D.A.; McLellan, W.A.; Raverty, S.; et al. Fetal distress and in utero pneumonia in perinatal dolphins during the Northern Gulf of Mexico unusual mortality Event. Dis. Aquat. Organ. 2016, 119, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtiss, J.B.; Colegrove, K.M.; Dianis, A.; Rotstein, D.; Fauquier, D.; Rowles, T.; Niemeyer, M.; Kinsel, M.J.; Terio, K. Pathology of Brucella ceti infection in cetaceans according to sequence type. In Proceedings of the IAAAM Conference, Durban, South Africa, 18 May 2019. [Google Scholar]
- Dagleish, M.P.; Barley, J.; Finlayson, J.; Reid, R.J.; Foster, G. Brucella ceti Associated pathology in the testicle of a Harbour porpoise (Phocoena phocoena). J. Comp. Pathol. 2008, 139, 54–59. [Google Scholar] [CrossRef]
- Foster, G.; MacMillan, A.P.; Godfroid, J.; Howie, F.; Ross, H.M.; Cloeckaert, A.; Reid, R.J.; Brew, S.; Patterson, I.A.P. A Review of Brucella sp. infection of sea mammals with particular emphasis on isolates from Scotland. Vet. Microbiol. 2002, 90, 563–580. [Google Scholar] [CrossRef]
- Jauniaux, T.P.; Brenez, C.; Fretin, D.; Godfroid, J.; Haelters, J.; Jacques, T.; Kerckhof, F.; Mast, J.; Sarlet, M.; Coignoul, F.L. Brucella ceti infection in Harbor porpoise (Phocoena phocoena). Emerg. Infect. Dis. 2010, 16, 1966–1968. [Google Scholar] [CrossRef]
- Maquart, M.; Le Flèche, P.; Foster, G.; Tryland, M.; Ramisse, F.; Djønne, B.; Al Dahouk, S.; Jacques, I.; Neubauer, H.; Walravens, K.; et al. MLVA-16 typing of 295 marine Brucella isolates from different animal and geographic origins identify 7 major groups within Brucella ceti and Brucella pinnipedialis. BMC Microbiol. 2009, 9, 145. [Google Scholar] [CrossRef]
- Ohishi, K.; Fujise, Y.; Maruyama, T. Brucella spp. in the Western North Pacific and Antarctic cetaceans: A review. J. Cetacean Res. Manag. 2008, 10, 67–72. [Google Scholar]
- West, K.L.; Levine, G.; Jacob, J.; Jensen, B.; Sanchez, S.; Colegrove, K.; Rotstein, D. Coinfection and vertical transmission of Brucella and Morbillivirus in a neonatal sperm whale (Physeter macrocephalus) in Hawaii, USA. J. Wildl. Dis. 2015, 51, 227–232. [Google Scholar] [CrossRef]
- May-Collado, L.J.; Amador-Caballero, M.; Casas, J.J.; Gamboa-Poveda, M.P.; Garita-Alpízar, F.; Gerrodette, T.; González-Barrientos, R.; Hernández-Mora, G.; Palacios, D.M.; Palacios-Alfaro, J.D.; et al. Ecology and conservation of cetaceans of Costa Rica and Panama. In Advances in Marine Vertebrate Research in Latin America; Rossi-Santos, M., Finkl, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 293–319. [Google Scholar] [CrossRef]
- Ortiz-Wolford, J.; Corona-Figueroa, M.F.; Dávila, V.; Cabrera, A.A. Cetacean stranding records along the Pacific coastline of Guatemala, 2007–2021: Implications for management, conservation, and research. Mar. Policy 2021, 134, 104827. [Google Scholar] [CrossRef]
- Portillo, R.I.; Sandoval, J.E.; Pineda, L.; Navas, E.M.; Mejía, M.G.P.; Fuentes, R.E.M. Registros de varamientos de cetáceos en El Salvador entre 1995–2019. Real. Y Reflexión 2021, 54, 218–240. [Google Scholar] [CrossRef]
- Suárez-Esquivel, M.; Baker, K.S.; Ruiz-Villalobos, N.; Hernández-Mora, G.; Barquero-Calvo, E.; González-Barrientos, R.; Castillo-Zeledón, A.; Jiménez-Rojas, C.; Chacón-Díaz, C.; Cloeckaert, A.; et al. Brucella genetic variability in wildlife marine mammals populations relates to host preference and ocean distribution. Genome Biol. Evol. 2017, 9, 1901–1912. [Google Scholar] [CrossRef] [PubMed]
- Garofolo, G.; Petrella, A.; Lucifora, G.; Di Francesco, G.; Di Guardo, G.; Pautasso, A.; Iulini, B.; Varello, K.; Giorda, F.; Goria, M.; et al. Occurrence of Brucella ceti in striped dolphins from Italian seas. PLoS ONE 2020, 15, e0240178. [Google Scholar] [CrossRef] [PubMed]
- Isidoro-Ayza, M.; Ruiz-Villalobos, N.; Pérez, L.; Guzmán-Verri, C.; Muñoz, P.M.; Alegre, F.; Barberán, M.; Chacón-Díaz, C.; Chaves-Olarte, E.; González-Barrientos, R.; et al. Brucella ceti infection in dolphins from the Western Mediterranean sea. BMC Vet. Res. 2014, 10, 206. [Google Scholar] [CrossRef] [Green Version]
- Davison, N.J.; Cranwell, M.P.; Perrett, L.L.; Dawson, C.E.; Stubberfield, E.J.; Jarvis, D.S.; Jepson, P.D. Meningoencephalitis associated with Brucella species in a live-stranded striped dolphin (Stenella coeruleoalba) in south-west England. Vet. Rec. Open 2009, 165, 86. [Google Scholar] [CrossRef]
- Gales, N.J. Mass stranding of striped dolphin, Stenella coeruleoalba, at Augusta, Western Australia: Notes on clinical pathology and general observations. J. Wildl. Dis. 1992, 28, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, A.; Yamada, T.K.; Tajima, Y.; Kunisue, T.; Amano, M.; Matsuishi, F. Diet of mass-stranded striped dolphins (Stenella coeruleoalba) in Southern Japan (East China Sea). Mammal Study 2020, 46, 17–24. [Google Scholar] [CrossRef]
- Vargas-Castro, I.; Crespo-Picazo, J.L.; Rivera-Arroyo, B.; Sánchez, R.; Marco- Cabedo, V.; Jiménez-Martinez, M.Á.; Fayos, M.; Sergio, Á.; Garcia- Párraga, D.; Sánchez-Vizcaino, J.M. Alpha and gammaherpesviruses in stranded striped dolphins (Stenella coeruleoalba) from Spain: First molecular detection of gammaherpesvirus infection in central nervous system of odontocetes. BMC Vet. Res. 2020, 16, 288. [Google Scholar] [CrossRef]
- Barraclough, A.; Sanz-Requena, R.; Marti-Bonmati, L.; Schmitt, T.L.; Jensen, E.; Garcia- Párraga, D. Radiographic assessment of pectoral flipper bone maturation in bottlenose dolphins (Tursiops truncatus), as a novel technique to accurately estimate chronological age. PLoS ONE 2019, 14, e0222722. [Google Scholar] [CrossRef]
- Nielsen, N.H.; Víkingsson, G.A.; Hansen, S.H.; Ditlevsen, S.; Heide-Jørgensen, M.P. Two techniques of age estimation in cetaceans: GLGs in teeth, earplugs, and measuring the AAR rate in eye lens nucleus. NAMMCO Sci. Publ. 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, N.; Nishiwaki, M. School structure of the striped dolphin off the Pacific coast of Japan. Sci. Rep. Whales Res. Inst. 1978, 30, 65–115. [Google Scholar]
- Ogden, J.A.; Conlogue, G.J.; Rhodin, A.G.J. Roentgenographic indicators of skeletal maturity in marine mammals (Cetacea). Skelet. Radiol. 1981, 7, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Dierauf, L.; Gulland, F.M. CRC Handbook of Marine Mammal Medicine: Health, Disease, and Rehabilitation, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2001; ISBN 1-4200-4163-0. [Google Scholar]
- Miller, D.L. Reproductive Biology and Phylogeny of Cetacea: Whales, Porpoises and Dolphins, 1st ed.; CRC Press: Boca Raton, FL, USA, 2007; ISBN 9780429063626. [Google Scholar]
- Geraci, J.R.; Lounsbury, V.J. Marine Mammals Ashore: A Field Guide for Strandings; National Aquarium: Baltimore, MD, USA, 2005; ISBN 0-9774609-0-8. [Google Scholar]
- United States Department of Agriculture USDA. National Veterinary Services Laboratories, Guidelines for Necropsy. 2021. Available online: https://www.aphis.usda.gov/animal_health/lab_info_services/downloads/NecropsyGuideline.pdf (accessed on 7 May 2021).
- Kasuya, T. Effect of exploitation on reproductive parameters of the spotted and striped dolphins off the Pacific coast of Japan. Sci. Rep. Whales Res. Inst. 1985, 36, 107–138. [Google Scholar]
- Read, F.L.; Hohn, A.A.; Lockyer, C.H. A review of age estimation methods in marine mammals with special reference to monodontids. NAMMCO Sci. Publ. 2018, 10. [Google Scholar] [CrossRef]
- Pérez, A. Validación de la Técnica de Corte Crudo Para la Determinación de Grupo de Líneas de Crecimiento (GLG) en Dientes de Delfines Rayados (Stenella coeruleoalba) Encallados por Brucella ceti en Costa Rica de 2004–2015; Universidad Técnica Nacional: Quesada, Costa Rica, 2015. [Google Scholar]
- Malatesta, M. Histological and histochemical methods—Theory and practice. Eur. J. Histochem. 2016, 60, 2639. [Google Scholar] [CrossRef] [Green Version]
- Ito, H.; Miyazaki, N. Skeletal development of the striped dolphin (Stenella coeruleoalba) in Japanese waters. J. Mamm. Soc. Jap. 1990, 14, 79–96. [Google Scholar] [CrossRef]
- Bacha, W.J., Jr.; Bacha, L.M. Color Atlas of Veterinary Histology, 3rd ed.; John Wiley & Sons: West Sussex, UK, 2012; ISBN 9788872873007. [Google Scholar]
- Harrison, R.J. Reproduction and reproductive organs. In The Biology of Marine Mammals; Academic Press: New York, NY, USA, 1969; pp. 253–348. [Google Scholar]
- Perrin, W.F.; Reilly, S.B. Reproductive Parameters of Dolphins and Small Whales of the Family Delphinidae; Bibliogov: Columbus, OH, USA, 2013; ISBN 9781287045595. [Google Scholar]
- Suárez-Esquivel, M.; Castro-Ramírez, L. Tetraiodothyronine, triiodothyronine, and cortisol serum levels in Costa Rican dogs using an immunoassay analyzer. Rev. Cienc. Vet. 2014, 30, 25–37. [Google Scholar]
- Yoshioka, M.; Aida, K.; Hanyu, I. Correlation of serum progesterone levels with reproductive status in female striped dolphins and short-finned pilot whales. Nippon. Suisan Gakkaishi 1989, 55, 475–478. [Google Scholar] [CrossRef]
- Barratclough, A.; Gomez, F.M.; Morey, J.S.; Megan, J.M.; Parry, C.; Schwacke, L.; Jensen, E.D.; Smith, C.R. Biochemical and hematological biomarkers of reproductive failure in bottlenose dolphins Tursiops truncatus. Dis. Aquat. Organ. 2021, 144, 197–208. [Google Scholar] [CrossRef]
- Calzada, N.; Aguilar, A.; Lockyer, C.; Grau, E. Patterns of growth and physical maturity in western Mediterranean striped dolphin, Stenella coeruleoalba. Can. J. Zool. 1997, 75, 632–637. [Google Scholar] [CrossRef]
- Siciliano, S.; Ramos, R.; Di Beneditto, A.P.; Santos, M.; Fragoso, A.B.; Brito, J.; Azevedo, A.F.; Vicente, A.F.; Zampirolli, E.; Alvarenga, F.; et al. Age and growth of some delphinids in South-Eastern Brazil. J. Mar. Biolog. 2007, 87, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Guarino, F.M.; Di-Nocera; Galiero, F.G.; Iaccarino, D.; Giglio, S.; Madeo, E.; Pollaro, F.; Mezzasalma, M.; Iavarone, I.; Odierna, G.; et al. Age estimation and growth of striped dolphins Stenella coeruleoalba stranded along the coasts of south-western Italy. Eur. Zool. J. 2021, 88, 417–424. [Google Scholar] [CrossRef]
- Kasuya, T. Life History and reproductive biology of the short-finned pilot whale, Globicephala macrorhychus, off the Pacific coast of Japan. Rep. Int. Whal. Common 1984, 6, 259–310. [Google Scholar]
- Au, D.W.K.; Perryman, W.L. Dolphin habitats in the Eastern Tropical Pacific. Fish. Bull. 1985, 83, 623–643. [Google Scholar]
- Rasmusson, E.M.; Arkin, P.A. A global view of large-scale precipitation variability. J. Clim. 1993, 6, 1495–1522. [Google Scholar] [CrossRef]
- Fiedler, P.C. The annual cycle and biological effects of the Costa Rica dome. Deep Sea Res. Part I Oceanogr. Res. Pap. 2002, 49, 321–338. [Google Scholar] [CrossRef]
- Perrin, W.F. Variation of spotted and spinner porpoise (Genus Stenella) in the Eastern Pacific and Hawaii. In UC San Diego: SIO; University of California Press, Ltd.: London, UK, 1975. [Google Scholar]
- Perrin, W.F. Subspecies of Stenella longirostris (Mammalia: Cetacea: Delphinidae). Proc. Biol. Soc. Wash. 1990, 103, 453–463. [Google Scholar]
- Perrin, W.F.; Akin, P.A.; Kashiwada, J.V. Geographic variation in external morphology of the spinner dolphin Stenella longirostris in the Eastern Pacific and implications for conservation. Fish. Bull. 1991, 89, 411–428. [Google Scholar]
- Perryman, W.L.; Lynn, M.S. Examination of stock and school structure of striped dolphin (Stenella coeruleoalba) in the Eastern Pacific from aerial photogrammetry. Fish. Bull. 1994, 92, 122–131. [Google Scholar]
- Perrin, W.F.; Scott, M.D.; Walker, G.J.; Cass, V.L. Review of Geographical Stocks of Tropical Dolphins (Stenella spp. and Delphinus Delphis) in the Eastern Pacific Item; NOAA Technical Report; NOAA: Washington, DC, USA, 1984. [Google Scholar]
- Fasick, J.I. Subspecific Variation in Coastal and Offshore Forms of Some Delphinid Species; University of Maryland: Baltimore, MD, USA, 1993. [Google Scholar]
- Crawford, R.P.; Adams, L.G.; Williams, J.D. Relationship of days in gestation at exposure and development of brucellosis in strain 19-vaccinated heifers. Am. J. Vet. Res. 1988, 49, 1037–1039. [Google Scholar]
- Gould, G.N.; Parkes, A.S.; Hammond, J.; Wilson, G.S.; Folley, S.J.; Steele-Bodger, H.W.; Scorgie, N.J.; Rowson, L.E.A. Brucellosis and sterility. J. R. Soc. Med. 1942, 35, 635–642. [Google Scholar] [CrossRef]
- Wilesmith, J.W. The persistence of Brucella abortus infection in calves: A retrospective study of heavily infected herds. Vet. Rec. 1978, 103, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Kasuya, T. Reconsideration of life history parameters of the spotted and striped dolphins based on cemental layers. Sci. Rep. Whales Res. Inst. 1976, 28, 73–106. [Google Scholar]
- Plön, S.; Bernard, R. Testis, spermatogenesis, and testicular cycles. Reproductive Biology and Phylogeny of Cetacea: Whales, Porpoises, and Dolphins; Miller, D.L., Ed.; CRC Press: Boca Raton, FL, USA, 2007; Volume 7, p. 215. [Google Scholar]
- Eberhardt, L.L.; Siniff, D.B. Population dynamics and marine mammal management policies. J. Fish. Res. Board Can. 1977, 34, 183–190. [Google Scholar] [CrossRef]
- DeMaster, D.P.; Edwards, E.F.; Wade, P.; Sisson, J.E. Status of dolphin stocks in the Eastern Tropical Pacific. In Wildlife 2001: Populations; McCullough, D.R., Barrett, R.H., Eds.; Springer: Dordrecht, The Netherlands, 1992; p. 101. ISBN 9781851668762. [Google Scholar]
- Reyna, R.; Traynor, K.; Hines, G.; Boots, L.R.; Aziz, R. Repeated freezing and thawing do not generally alter assay results for several commonly studied reproductive hormones. Fertil. Steril. 2001, 76, 823–825. [Google Scholar] [CrossRef]
- Hegstad-Davies, R.L. A review of sample handling considerations for reproductive and thyroid hormone measurement in serum or plasma. Theriogenology 2006, 66, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Ohsumi, S. Comparison of maturity and accumulation rate of corpora albicans between the left and right ovaries in Cetacea. Sci. Rep. Whales Res. Inst. 1964, 18, 123–148. [Google Scholar]
- Karakosta, C.V.; Jepson, P.D.; Ohira, H.; Moore, A.; Bennett, P.M.; Holt, W.V. Testicular and ovarian development in the Harbour Porpoise (Phocoena phocoena). J. Zool. 1999, 249, 111–121. [Google Scholar] [CrossRef]
- Aguilar, A.; Borrell, A. Abnormally high polychlorinated biphenyl levels in striped dolphins (Stenella coeruleoalba) affected by the 1990–1992 Mediterranean epizootic. Sci. Total Environ. 1994, 154, 237–247. [Google Scholar] [CrossRef]
- Munson, L.; Calzada, N.; Kennedy, S.; Sorensen, T.B. Luteinized ovarian cysts in Mediterranean striped dolphins. J. Wildl. Dis. 1998, 34, 656–660. [Google Scholar] [CrossRef] [Green Version]
- Hall, A.J.; McConnell, B.J.; Schwacke, L.H.; Ylitalo, G.M.; Williams, R.; Rowles, T.K. Predicting the effects of polychlorinated biphenyls on cetacean populations through impacts on immunity and calf Survival. Environ. Pollut. 2018, 233, 407–418. [Google Scholar] [CrossRef] [Green Version]
- McFee, W.E.; Wu, D.; Colegrove, K.; Terio, K.; Balthis, L.; Young, R. Occurrence of Brucella ceti in stranded bottlenose dolphins Tursiops truncatus coincides with calving season. Dis. Aquat. Org. 2020, 141, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Schwacke, L.H.; Smith, C.R.; Townsend, F.I.; Wells, R.S.; Hart, L.B.; Balmer, B.C.; Collier, T.K.; De Guise, S.; Fry, M.M.; Guillette, L.J., Jr.; et al. Health of common bottlenose dolphins (Tursiops truncatus) in Barataria Bay, Louisiana, following the Deepwater Horizon Oil Spill. Environ. Sci. Technol. 2014, 48, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.R.; Rowles, T.K.; Hart, L.B.; Townsend, F.I.; Wells, R.S.; Zolman, E.S.; Balmer, B.C.; Quigley, B.; Ivančić, M.; McKercher, W.; et al. Slow recovery of Barataria Bay dolphin health following the Deepwater Horizon oil spill (2013–2014), with evidence of persistent lung disease and impaired stress response. Endanger. Species Res. 2017, 33, 127–142. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Mora, G.; Manire, C.A.; González-Barrientos, R.; Barquero-Calvo, E.; Guzmán-Verri, C.; Staggs, L.; Thompson, R.; Chaves-Olarte, E.; Moreno, E. Serological diagnosis of Brucella infections in Odontocetes. Clin. Vaccine Immunol. 2009, 16, 906–915. [Google Scholar] [CrossRef] [PubMed]
- George, T. Assessing Brucella ceti Infections in Oregon and Washington Dolphins that Stranded with Histopathological Lesions Resembling Neurobrucellosis 2006–2014; Evergreen State College: Olympia, WA, USA, 2015. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roca-Monge, K.; González-Barrientos, R.; Suárez-Esquivel, M.; Palacios-Alfaro, J.D.; Castro-Ramírez, L.; Jiménez-Soto, M.; Cordero-Chavarría, M.; García-Párraga, D.; Barratclough, A.; Moreno, E.; et al. Age and Sexual Maturity Estimation of Stranded Striped Dolphins, Stenella coeruleoalba, Infected with Brucella ceti. Oceans 2022, 3, 494-508. https://doi.org/10.3390/oceans3040033
Roca-Monge K, González-Barrientos R, Suárez-Esquivel M, Palacios-Alfaro JD, Castro-Ramírez L, Jiménez-Soto M, Cordero-Chavarría M, García-Párraga D, Barratclough A, Moreno E, et al. Age and Sexual Maturity Estimation of Stranded Striped Dolphins, Stenella coeruleoalba, Infected with Brucella ceti. Oceans. 2022; 3(4):494-508. https://doi.org/10.3390/oceans3040033
Chicago/Turabian StyleRoca-Monge, Karol, Rocío González-Barrientos, Marcela Suárez-Esquivel, José David Palacios-Alfaro, Laura Castro-Ramírez, Mauricio Jiménez-Soto, Minor Cordero-Chavarría, Daniel García-Párraga, Ashley Barratclough, Edgardo Moreno, and et al. 2022. "Age and Sexual Maturity Estimation of Stranded Striped Dolphins, Stenella coeruleoalba, Infected with Brucella ceti" Oceans 3, no. 4: 494-508. https://doi.org/10.3390/oceans3040033
APA StyleRoca-Monge, K., González-Barrientos, R., Suárez-Esquivel, M., Palacios-Alfaro, J. D., Castro-Ramírez, L., Jiménez-Soto, M., Cordero-Chavarría, M., García-Párraga, D., Barratclough, A., Moreno, E., & Hernández-Mora, G. (2022). Age and Sexual Maturity Estimation of Stranded Striped Dolphins, Stenella coeruleoalba, Infected with Brucella ceti. Oceans, 3(4), 494-508. https://doi.org/10.3390/oceans3040033