
Inflammation and Preterm Birth: A Systematic Review


{kind=link}
{kind=link}
{kind=link}
Abstract
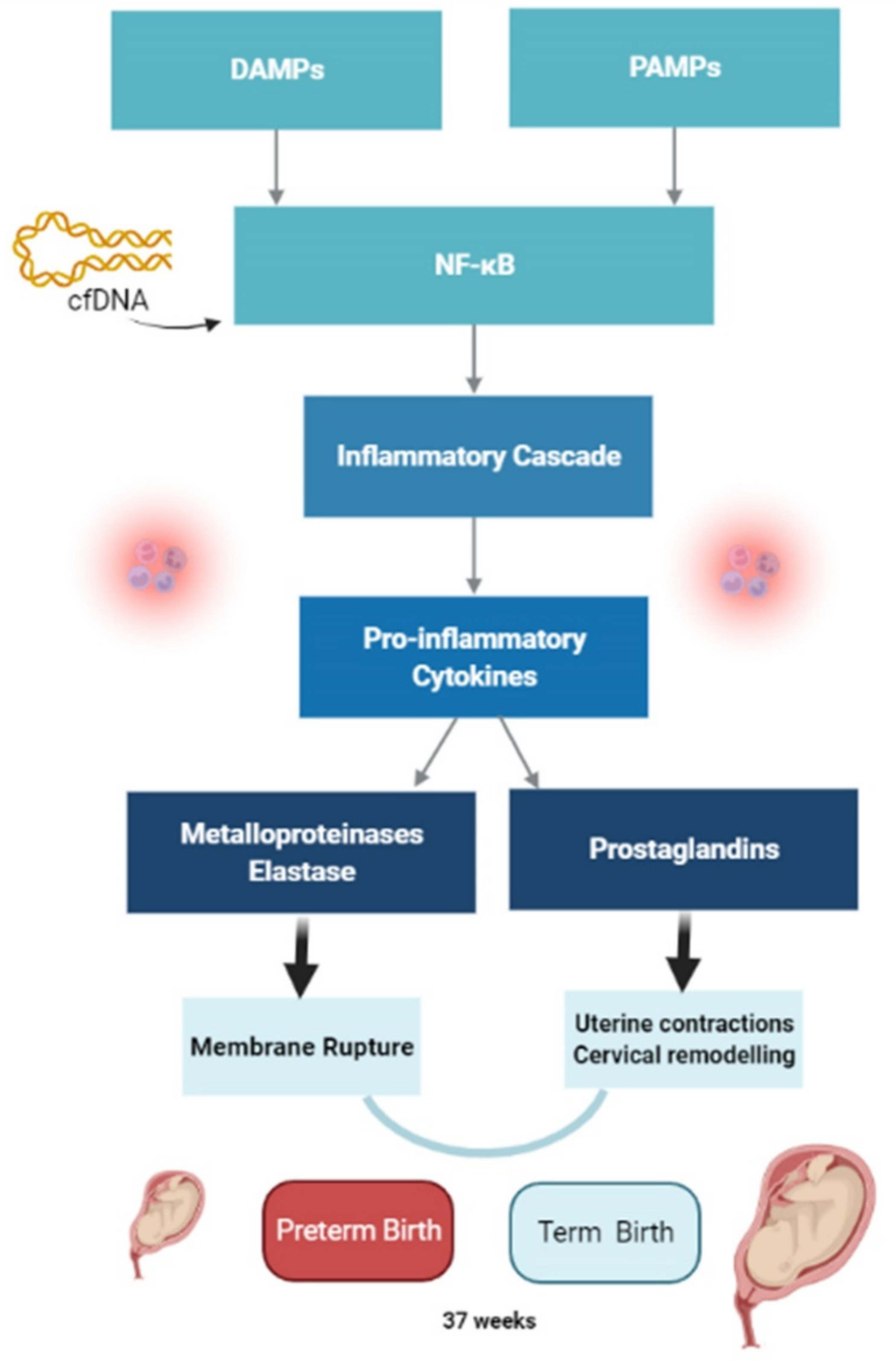
:1. Introduction
2. Materials and Methods
2.1. Eligibility Criteria
2.2. Information Sources and Search Strategy
2.3. Study Selection
2.4. Data Extraction
3. Results
- Risk of bias within studies and across studies
4. Discussion
4.1. Immuno-Inflammation and Normal Delivery
4.2. Immuno-Inflammation and Preterm Birth
- Genetic factors and PTB
4.3. Reproductive Hormones and Labour
4.4. New Approaches in PTB Treatment
- Drugs that interfere with nPRA/nPRB balance:
- (a)
- Selective progesterone receptor modulators (SPRMs) boost P4 anti-inflammatory activity that blocks labour, while at the same time preventing PR phosphorylation (of serine residue 344 and 345 in nPRA) [8], leading to a loss of P4 anti-inflammatory activity [14]. SPRMs are compounds that compete with P4 for binding to the nuclear PR ligand-binding pocket and induce mixed agonist/antagonist activity [16]. Nuclear PRs are presumably saturated by progesterone during pregnancy [14]. Since membrane PR have a lower binding affinity, they may be pharmacologically affected by additional P4 [14]. Moreover, it is possible that nuclear PR signalling in myometrial, endometrial and decidual cells might be affected by an interaction with membrane PR activity, altered by exogenous P4 [14].
- (b)
- Histone deacetylase inhibitors (Trichostatin A): seems to promote nPRB expression in human myometrial samples [8].
- (c)
- Chloroquine: human myometrial cells express bitter taste receptors, and the activation of these receptors leads to muscle relaxation. Additionally, chloroquine, as a bitter tastant, is also able to prevent inflammation induced PTB [8].
- PGF2 α antagonists: recent research showed that, when used in combination with Atosiban, delivery was reduced 55% more efficiently than with Atosiban alone [5].
- TLR4 antagonists (naloxone, naltrexone, and rosiglitazone) diminish proinflammatory cytokines [8]. Naloxone (+)- is an isomer of the antagonist of opioid receptor naloxone (−)-. Naloxone (+)- has anti-inflammatory properties with the capacity to specifically antagonize TLR4; nevertheless, it has no opioid activity [6]. In animal models, it reduces PTB, foetal demise and cytokine expression in intrauterine tissues [6]. Additionally, it has the benefit of providing a greater blockage than other more distally located targets [6]. However, drugs targeting TLR4 may only be useful in PTB triggered by Gram-negative bacteria (which can elicit inflammation by another TLR, rendering some freedom to the birth pathway) [6].
- Cytokine-suppressive anti-inflammatory drugs (CSAIDs) inhibit intracellular pathways that result in NF-κB activation [6]. These drugs are more selective and effective than NSAIDs with no adverse neonatal effects (specifically abortion and renal impairment). The classes being studied are TAK1 inhibitors; P38 MAPK inhibitors and IKK complex inhibitors (of which TPCA 1 is the most promising) [6].
- NF-κB inhibitors: N-acetylcysteine is involved in reducing oxidative stress, with promising results in PTB reduction and an additional benefit in foetal neuroprotection. Sulfasalazine, a drug frequently used in inflammatory bowel disease, inhibits IKK kinases and seems to reduce inflammatory cytokine release in vitro [6].
- Drugs targeting specific proinflammatory cytokines:
- TNF-α inhibitors (infliximab and etanercept) are only studied in mice but raise concerns regarding the increased risk of neonatal infection [6].
- Polyunsaturated fatty acids (PUFA) and lipid metabolites are known to resolve oxidative stress and inflammation through arachidonic acid pathways. The balance between metabolites of omega-3 and omega-6 PUFA seems to be important in pregnancy maintenance and have a significant role in cervical effacement, dilation and initiation of labour. θ3 PUFA have anti-inflammatory actions but θ6 have proinflammatory properties [6]. The recent ORIP trial found that they did not reduce PTB [6]. However, the new molecules (specialized pro-resolving lipid mediators, such as lipoxin A4 and resolving E2) have not been studied in humans [6]. Moreover, plasma N-acylethanolamines were recently proposed as a single simple test that could be used to predict PTB in high-risk women [20].
- Polyphenols suppress the production of proinflammatory cytokines such as TNF-α and IL-1β. Resveratrol has shown good results, but the increase in foetal pancreatic mass makes its use undesirable [6].
5. Final Comment
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Keelan, J.A. Intrauterine inflammatory activation, functional progesterone withdrawal, and the timing of term and preterm birth. J. Reprod. Immunol. 2018, 125, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Kalagiri, R.R.; Carder, T.; Choudhury, S.; Vora, N.; Ballard, A.R.; Govande, V.; Drever, N.; Beeram, M.R.; Uddin, M.N. Inflammation in Complicated Pregnancy and Its Outcome. Am. J. Perinatol. 2016, 33, 1337–1356. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Behnia, F.; Polettini, J.; Richardson, L.S. Novel pathways of inflammation in human fetal membranes associated with preterm birth and preterm pre-labor rupture of the membranes. Semin. Immunopathol. 2020, 42, 431–450. [Google Scholar] [CrossRef]
- Olson, D.M.; Mijovic, J.E.; Sadowsky, D.W. Control of human parturition. Semin. Perinatol. 1995, 19, 52–63. [Google Scholar] [CrossRef]
- Leimert, K.B.; Xu, W.; Princ, M.M.; Chemtob, S.; Olson, D.M. Inflammatory Amplification: A Central Tenet of Uterine Transition for Labor. Front. Cell Infect. Microbiol. 2021, 11, 660983. [Google Scholar] [CrossRef] [PubMed]
- Triggs, T.; Kumar, S.; Mitchell, M. Experimental drugs for the inhibition of preterm labor. Expert. Opin. Investig. Drugs 2020, 29, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Boyle, A.K.; Rinaldi, S.F.; Norman, J.E.; Stock, S.J. Preterm birth: Inflammation, fetal injury and treatment strategies. J. Reprod. Immunol. 2017, 119, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Zierden, H.C.; Shapiro, R.L.; DeLong, K.; Carter, D.M.; Ensign, L.M. Next generation strategies for preventing preterm birth. Adv. Drug Deliv. Rev. 2021, 174, 190–209. [Google Scholar] [CrossRef]
- Singh, N.; Bonney, E.; McElrath, T.; Lamont, R.F.; Preterm Birth International collaborative (PREBIC). Prevention of preterm birth: Proactive and reactive clinical practice-are we on the right track? Placenta 2020, 98, 6–12. [Google Scholar] [CrossRef]
- Bonney, E.A.; Johnson, M.R. The role of maternal T cell and macrophage activation in preterm birth: Cause or consequence? Placenta 2019, 79, 53–61. [Google Scholar] [CrossRef]
- Di Renzo, G.C.; Cabero Roura, L.; Facchinetti, F.; Helmer, H.; Hubinont, C.; Jacobsson, B.; Jorgensen, J.S.; Lamont, R.F.; Mikhailov, A.; Papantoniou, N.; et al. Preterm Labor and Birth Management: Recommendations from the European Association of Perinatal Medicine. J. Matern. Fetal Neonatal. Med. 2017, 30, 2011–2030. [Google Scholar] [CrossRef]
- Di Renzo, G.C.; Tosto, V.; Giardina, I. The biological basis and prevention of preterm birth. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 52, 13–22. [Google Scholar] [CrossRef]
- Gilman-Sachs, A.; Dambaeva, S.; Salazar Garcia, M.D.; Hussein, Y.; Kwak-Kim, J.; Beaman, K. Inflammation induced preterm labor and birth. J. Reprod. Immunol. 2018, 129, 53–58. [Google Scholar] [CrossRef]
- Green, E.S.; Arck, P.C. Pathogenesis of preterm birth: Bidirectional inflammation in mother and fetus. Semin. Immunopathol. 2020, 42, 413–429. [Google Scholar] [CrossRef]
- Herrera, C.A.; Stoerker, J.; Carlquist, J.; Stoddard, G.J.; Jackson, M.; Esplin, S.; Rose, N.C. Cell-free DNA, inflammation, and the initiation of spontaneous term labor. Am. J. Obstet. Gynecol. 2017, 217, 583.e1–583.e8. [Google Scholar] [CrossRef]
- Mesiano, S.A.; Peters, G.A.; Amini, P.; Wilson, R.A.; Tochtrop, G.P.; van Den Akker, F. Progestin therapy to prevent preterm birth: History and effectiveness of current strategies and development of novel approaches. Placenta 2019, 79, 46–52. [Google Scholar] [CrossRef]
- Strauss, J.F., 3rd; Romero, R.; Gomez-Lopez, N.; Haymond-Thornburg, H.; Modi, B.P.; Teves, M.E.; Pearson, L.N.; York, T.P.; Schenkein, H.A. Spontaneous preterm birth: Advances toward the discovery of genetic predisposition. Am. J. Obstet. Gynecol. 2018, 218, 294–314.e2. [Google Scholar] [CrossRef]
- Vora, B.; Wang, A.; Kosti, I.; Huang, H.; Paranjpe, I.; Woodruff, T.J.; MacKenzie, T.; Sirota, M. Meta-Analysis of Maternal and Fetal Transcriptomic Data Elucidates the Role of Adaptive and Innate Immunity in Preterm Birth. Front. Immunol. 2018, 9, 993. [Google Scholar] [CrossRef]
- Combs, C.A.; Gravett, M.; Garite, T.J.; Hickok, D.E.; Lapidus, J.; Porreco, R.; Rael, J.; Grove, T.; Morgan, T.K.; Clewell, W.; et al. Amniotic fluid infection, inflammation, and colonization in preterm labor with intact membranes. Am. J. Obstet. Gynecol. 2014, 210, 125.e1–125.e15. [Google Scholar] [CrossRef]
- Bachkangi, P.; Taylor, A.H.; Bari, M.; Maccarrone, M.; Konje, J.C. Prediction of preterm labour from a single blood test: The role of the endocannabinoid system in predicting preterm birth in high-risk women. Eur. J. Obstet. Gynecol. Reprod. Biol. 2019, 243, 1–6. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; The PRISMA Group. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Areia, A.L.; Mota-Pinto, A. Inflammation and Preterm Birth: A Systematic Review. Reprod. Med. 2022, 3, 101-111. https://doi.org/10.3390/reprodmed3020009
Areia AL, Mota-Pinto A. Inflammation and Preterm Birth: A Systematic Review. Reproductive Medicine. 2022; 3(2):101-111. https://doi.org/10.3390/reprodmed3020009
Chicago/Turabian StyleAreia, Ana Luísa, and Anabela Mota-Pinto. 2022. "Inflammation and Preterm Birth: A Systematic Review" Reproductive Medicine 3, no. 2: 101-111. https://doi.org/10.3390/reprodmed3020009
APA StyleAreia, A. L., & Mota-Pinto, A. (2022). Inflammation and Preterm Birth: A Systematic Review. Reproductive Medicine, 3(2), 101-111. https://doi.org/10.3390/reprodmed3020009