

Genomic Insults and their Redressal in the Eutopic Endometrium of Women with Endometriosis

{kind=link}
Abstract
:1. Introduction
2. DNA Damage and Its Repair in the Endometrium
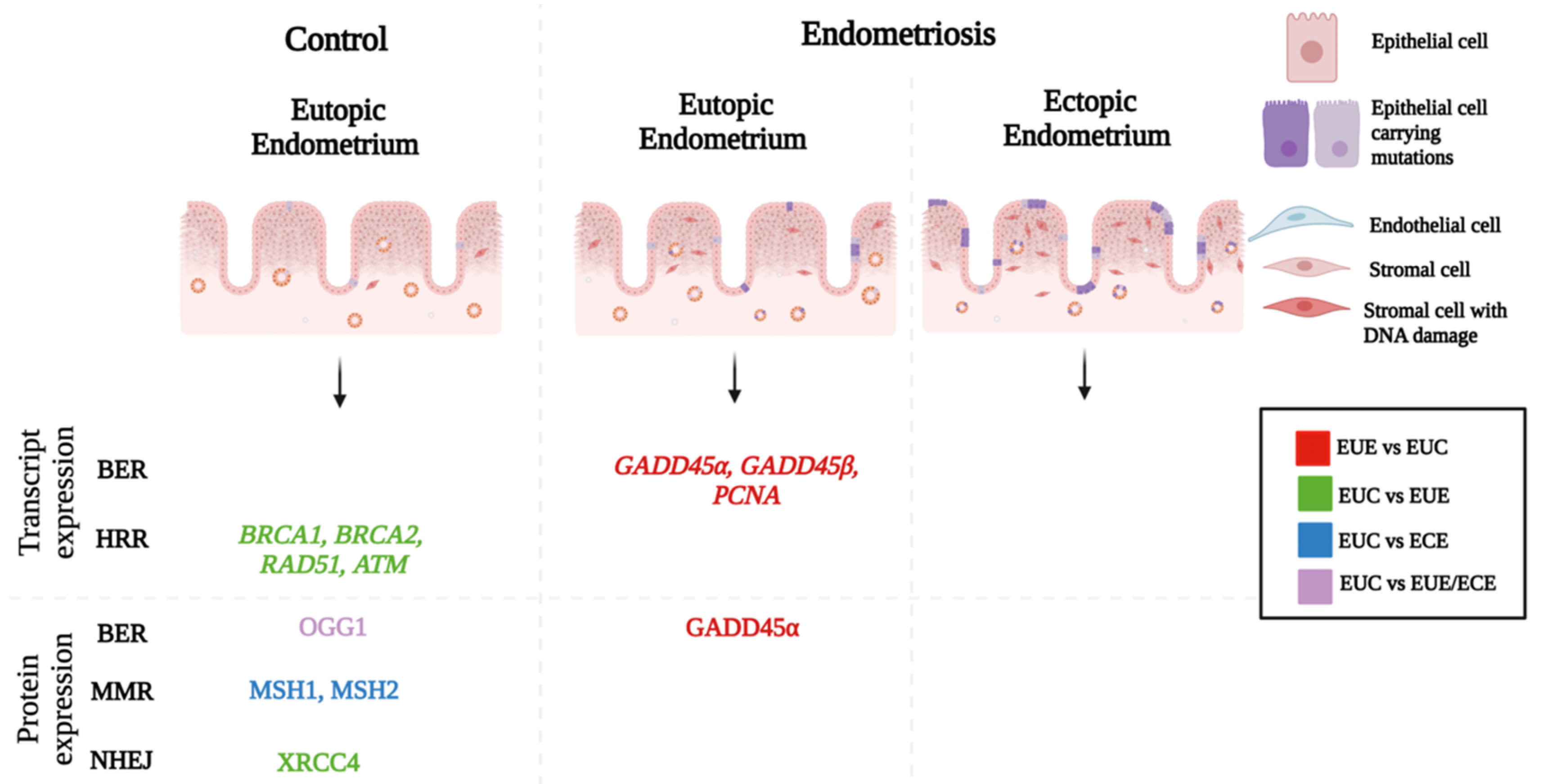
3. Endometrial DNA Damage in Endometriosis
4. Potential Causes of DNA Damage
4.1. Proliferative Stress
4.2. Oxidative Stress
4.3. Inflammation
4.4. Environmental Toxicants
4.5. Estrogenic Toxicity
5. DNA Repair in Women with Endometriosis
6. Conclusions
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Friedberg, E.C. DNA damage and repair. Nature 2003, 421, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Sancar, A.; Lindsey-Boltz, L.A.; Ünsal-Kaçmaz, K.; Linn, S. Molecular Mechanisms of Mammalian DNA Repair and the DNA Damage Checkpoints. Annu. Rev. Biochem. 2004, 73, 39–85. [Google Scholar] [CrossRef] [PubMed]
- Blanpain, C.; Mohrin, M.; Sotiropoulou, P.A.; Passegué, E. DNA-Damage Response in Tissue-Specific and Cancer Stem Cells. Cell Stem Cell 2011, 8, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.; Lee, S.; Kang, H.; Kim, J.; Kim, K.; Youn, H.S.; Jin, Y.W.; Seo, S.; Youn, B.H. Organ-Specific Effects of Low Dose Radiation Exposure: A Comprehensive Review. Front. Genet. 2020, 11, 1178. [Google Scholar] [CrossRef] [PubMed]
- Mohrin, M.; Bourke, E.; Alexander, D.; Warr, M.R.; Barry-Holson, K.; Le Beau, M.M.; Morrison, C.G.; Passegué, E. Hematopoietic stem cell quiescence promotes error-prone DNA repair and mutagenesis. Cell Stem Cell 2010, 7, 174–185. [Google Scholar] [CrossRef]
- Harding, S.M.; Benci, J.L.; Irianto, J.; Discher, D.E.; Minn, A.J.; Greenberg, R.A. Mitotic progression following DNA damage enables pattern recognition within micronuclei. Nature 2017, 548, 466–470. [Google Scholar] [CrossRef]
- Kiraly, O.; Gong, G.; Olipitz, W.; Muthupalani, S.; Engelward, B.P. Inflammation-Induced Cell Proliferation Potentiates DNA Damage-Induced Mutations In Vivo. PLoS Genet. 2015, 11, e1004901. [Google Scholar] [CrossRef]
- Stopper, H.; Schmitt, E.; Gregor, C.; Mueller, S.O.; Fischer, W.H. Increased cell proliferation is associated with genomic instability: Elevated micronuclei frequencies in estradiol-treated human ovarian cancer cells. Mutagenesis 2003, 18, 243–247. [Google Scholar] [CrossRef]
- de Pedro, I.; Alonso-Lecue, P.; Sanz-Gómez, N.; Freije, A.; Gandarillas, A. Sublethal UV irradiation induces squamous differentiation via a p53-independent, DNA damage-mitosis checkpoint. Cell Death Dis. 2018, 9, 1094. [Google Scholar] [CrossRef]
- Sanz-Gómez, N.; Freije, A.; Ceballos, L.; Obeso, S.; Sanz, J.R.; García-Reija, F.; Morales-Angulo, C.; Gandarillas, A. Response of head and neck epithelial cells to a DNA damage-differentiation checkpoint involving polyploidization. Head Neck 2018, 40, 2487–2497. [Google Scholar] [CrossRef]
- Bredemeyer, A.L.; Helmink, B.A.; Innes, C.L.; Calderon, B.; McGinnis, L.M.; Mahowald, G.K.; Gapud, E.J.; Walker, L.M.; Collins, J.B.; Weaver, B.K.; et al. DNA double-strand breaks activate a multi-functional genetic program in developing lymphocytes. Nature 2008, 456, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Inomata, K.; Aoto, T.; Binh, N.T.; Okamoto, N.; Tanimura, S.; Wakayama, T.; Iseki, S.; Hara, E.; Masunaga, T.; Shimizu, H.; et al. Genotoxic Stress Abrogates Renewal of Melanocyte Stem Cells by Triggering Their Differentiation. Cell 2009, 137, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Bane, K.; Desouza, J.; Shetty, D.; Choudhary, P.; Kadam, S.; Katkam, R.R.; Fernandes, G.; Sawant, R.; Dudhedia, U.; Warty, N.; et al. Endometrial DNA damage response is modulated in endometriosis. Hum. Reprod. 2021, 36, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Grassi, T.; Calcagno, A.; Marzinotto, S.; Londero, A.P.; Orsaria, M.; Canciani, G.N.; Beltrami, C.A.; Marchesoni, D.; Mariuzzi, L. Mismatch repair system in endometriotic tissue and eutopic endometrium of unaffected women. Int. J. Clin. Exp. Pathol. 2015, 8, 1867–1877. [Google Scholar]
- Fuseya, C.; Horiuchi, A.; Hayashi, A.; Suzuki, A.; Miyamoto, T.; Hayashi, T.; Shiozawa, T. Involvement of pelvic inflammation-related mismatch repair abnormalities and microsatellite instability in the malignant transformation of ovarian endometriosis. Hum. Pathol. 2012, 43, 1964–1972. [Google Scholar] [CrossRef]
- Choi, Y.S.; Park, J.H.; Lee, J.H.; Yoon, J.K.; Yun, B.H.; Park, J.H.; Seo, S.K.; Sung, H.-J.; Kim, H.-S.; Cho, S.H.; et al. Association Between Impairment of DNA Double Strand Break Repair and Decreased Ovarian Reserve in Patients With Endometriosis. Front. Endocrinol. 2018, 9, 772. [Google Scholar] [CrossRef]
- Bane, K.; Desouza, J.; Rojewale, A.; Katkam, R.R.; Fernandes, G.; Sawant, R.; Dudhedia, U.; Warty, N.; Chauhan, A.; Chaudhari, U.; et al. Dysregulation of X-ray repair cross-complementing 4 expression in the eutopic endometrium of women with endometriosis. Reproduction 2022, 163, 95–105. [Google Scholar] [CrossRef]
- Carvalho, L.F.P.; Abrão, M.S.; Biscotti, C.; Sharma, R.; Nutter, B.; Falcone, T. Oxidative cell injury as a predictor of endometriosis progression. Reprod. Sci. 2013, 20, 688–698. [Google Scholar] [CrossRef]
- Suda, K.; Nakaoka, H.; Yoshihara, K.; Ishiguro, T.; Tamura, R.; Mori, Y.; Yamawaki, K.; Adachi, S.; Takahashi, T.; Kase, H.; et al. Clonal Expansion and Diversification of Cancer-Associated Mutations in Endometriosis and Normal Endometrium. Cell Rep. 2018, 24, 1777–1789. [Google Scholar] [CrossRef]
- Lac, V.; Verhoef, L.; Aguirre-Hernandez, R.; Nazeran, T.M.; Tessier-Cloutier, B.; Praetorius, T.; Orr, N.L.; Noga, H.; Lum, A.; Khattra, J.; et al. Iatrogenic endometriosis harbors somatic cancer-driver mutations. Hum. Reprod. 2019, 34, 69–78. [Google Scholar] [CrossRef]
- Wiegand, K.C.; Shah, S.P.; Al-Agha, O.M.; Zhao, Y.; Tse, K.; Zeng, T.; Senz, J.; McConechy, M.K.; Anglesio, M.S.; Kalloger, S.E.; et al. ARID1A mutations in endometriosis-associated ovarian carcinomas. N. Engl. J. Med. 2010, 363, 1532–1543. [Google Scholar] [CrossRef]
- Chandler, R.L.; Damrauer, J.S.; Raab, J.R.; Schisler, J.C.; Wilkerson, M.D.; Didion, J.P.; Starmer, J.; Serber, D.; Yee, D.; Xiong, J.; et al. Coexistent ARID1A-PIK3CA mutations promote ovarian clear-cell tumorigenesis through pro-tumorigenic inflammatory cytokine signalling. Nat. Commun. 2015, 6, 6118. [Google Scholar] [CrossRef] [PubMed]
- Obata, K.; Hoshiai, H. Common genetic changes between endometriosis and ovarian cancer. Gynecol. Obstet. Invest. 2000, 50 (Suppl. 1), 39–43. [Google Scholar] [CrossRef] [PubMed]
- Anglesio, M.S.; Papadopoulos, N.; Ayhan, A.; Nazeran, T.M.; Noë, M.; Horlings, H.M.; Lum, A.; Jones, S.; Senz, J.; Seckin, T.; et al. Cancer-Associated Mutations in Endometriosis without Cancer. N. Engl. J. Med. 2017, 376, 1835–1848. [Google Scholar] [CrossRef]
- ”Endometriosis|ASRM”. Available online: https://www.asrm.org/topics/topics-index/endometriosis/ (accessed on 30 December 2022).
- Gajbhiye, R.K.; Montgomery, G.; Pai, M.V.; Phukan, P.; Shekhar, S.; Padte, K.; Dasmahapatra, P.; John, B.M.; Shembekar, C.; Bhurke, A.V.; et al. Protocol for a case–control study investigating the clinical phenotypes and genetic regulation of endometriosis in Indian women: The ECGRI study. BMJ Open 2021, 11, e050844. [Google Scholar] [CrossRef] [PubMed]
- Sourial, S.; Tempest, N.; Hapangama, D.K. Theories on the Pathogenesis of Endometriosis. Int. J. Reprod. Med. 2014, 2014, 179515. [Google Scholar] [CrossRef] [PubMed]
- Pearce, C.L.; Templeman, C.; Rossing, M.A.; Lee, A.; Near, A.M.; Webb, P.M.; Nagle, C.M.; Doherty, J.A.; Cushing-Haugen, K.L.; Wicklund, K.G.; et al. Association between endometriosis and risk of histological subtypes of ovarian cancer: A pooled analysis of case-control studies. Lancet. Oncol. 2012, 13, 385–394. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, T.H.; Chung, H.H.; Song, Y.S. Risk and prognosis of ovarian cancer in women with endometriosis: A meta-analysis. Br. J. Cancer 2014, 110, 1878–1890. [Google Scholar] [CrossRef]
- Koppolu, A.; Maksym, R.B.; Paskal, W.; Machnicki, M.; Rak, B.; Pępek, M.; Garbicz, F.; Pełka, K.; Kuśmierczyk, Z.; Jacko, J.; et al. Epithelial cells of deep infiltrating endometriosis harbor mutations in cancer driver genes. Cells 2021, 10, 749. [Google Scholar] [CrossRef]
- McKinnon, B.D.; Lukowski, S.W.; Mortlock, S.; Crawford, J.; Atluri, S.; Subramaniam, S.; Johnston, R.L.; Nirgianakis, K.; Tanaka, K.; Amoako, A.; et al. Altered differentiation of endometrial mesenchymal stromal fibroblasts is associated with endometriosis susceptibility. Commun. Biol. 2022, 5, 600. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Yoshihara, K.; Suda, K.; Nakaoka, H.; Yachida, N.; Ueda, H.; Sugino, K.; Mori, Y.; Yamawaki, K.; Tamura, R.; et al. Three-dimensional understanding of the morphological complexity of the human uterine endometrium. iScience 2021, 24, 102258. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Nakaoka, H.; Suda, K.; Yoshihara, K.; Ishiguro, T.; Yachida, N.; Saito, K.; Ueda, H.; Sugino, K.; Mori, Y.; et al. Spatiotemporal dynamics of clonal selection and diversification in normal endometrial epithelium. Nat. Commun. 2022, 13, 943. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zhang, X.; Chen, Y. Mutagen sensitivity as a susceptibility marker for endometriosis. Hum. Reprod. 2003, 18, 2052–2057. [Google Scholar] [CrossRef]
- Hapangama, D.K.; Turner, M.A.; Drury, J.A.; Quenby, S.; Hart, A.; Maddick, M.; Martin-Ruiz, C.; Von Zglinicki, T. Sustained replication in endometrium of women with endometriosis occurs without evoking a DNA damage response. Hum. Reprod. 2009, 24, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Kao, S.H.; Huang, H.C.; Hsieh, R.H.; Chen, S.C.; Tsai, M.C.; Tzeng, C.R. Oxidative Damage and Mitochondrial DNA Mutations with Endometriosis. Ann. N. Y. Acad. Sci. 2005, 1042, 186–194. [Google Scholar] [CrossRef]
- Creed, J.; Maggrah, A.; Reguly, B.; Harbottle, A. Mitochondrial DNA deletions accurately detect endometriosis in symptomatic females of child-bearing age. Biomark. Med. 2019, 13, 291–306. [Google Scholar] [CrossRef]
- Govatati, S.; Tipirisetti, N.R.; Perugu, S.; Kodati, V.L.; Deenadayal, M.; Satti, V.; Bhanoori, M.; Shivaji, S. Mitochondrial genome variations in advanced stage endometriosis: A study in South Indian population. PLoS ONE 2012, 7, e40668. [Google Scholar] [CrossRef]
- Atkins, H.M.; Bharadwaj, M.S.; O’Brien Cox, A.; Furdui, C.M.; Appt, S.E.; Caudell, D.L. Endometrium and endometriosis tissue mitochondrial energy metabolism in a nonhuman primate model. Reprod. Biol. Endocrinol. 2019, 17, 70. [Google Scholar] [CrossRef]
- Kahyaoglu, I.; Kahyaoglu, S.; Moraloglu, O.; Zergeroglu, S.; Sut, N.; Batioglu, S. Comparison of Ki-67 proliferative index between eutopic and ectopic endometrium: A case control study. Taiwan. J. Obstet. Gynecol. 2012, 51, 393–396. [Google Scholar] [CrossRef]
- Eris Yalcin, S.; Ocal, I.; Yalcin, Y.; Sen Selim, H.; Demir Caltekin, M.; Aydogmus, H.; Kelekci, S. Evaluation of the Ki-67 Proliferation Index and Urocortin Expression in Women with Ovarian Endometriomas. Eurasian J. Med. 2017, 49, 107. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.M.; Oh, Y.J.; Cho, S.H.; Chung, D.J.; Hwang, J.Y.; Park, K.H.; Cho, D.J.; Choi, Y.M.; Lee, B.S. Increased telomerase activity and human telomerase reverse transcriptase mRNA expression in the endometrium of patients with endometriosis. Hum. Reprod. 2007, 22, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Alnafakh, R.; Choi, F.; Bradfield, A.; Adishesh, M.; Saretzki, G.; Hapangama, D.K. Endometriosis Is Associated with a Significant Increase in hTERC and Altered Telomere/Telomerase Associated Genes in the Eutopic Endometrium, an Ex-Vivo and In Silico Study. Biomedicines 2020, 8, 588. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zeng, W.; Xu, S.; Nie, J.; Huang, L.; Lai, Y.; Yu, Y. Up-regulation of DNA2 results in cell proliferation and migration in endometriosis. J. Mol. Histol. 2021, 52, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Lee, J.H.; Kim, M.; Chang, H.J.; Hwang, K.J.; Chang, K.H. Endometrium from women with endometriosis shows increased proliferation activity. Fertil. Steril. 2009, 92, 1246–1249. [Google Scholar] [CrossRef] [PubMed]
- Ferda Verit, F.; Erel, O.; Celik, N. Serum paraoxonase-1 activity in women with endometriosis and its relationship with the stage of the disease. Hum. Reprod. 2008, 23, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Santulli, P.; Chouzenoux, S.; Fiorese, M.; Marcellin, L.; Lemarechal, H.; Millischer, A.E.; Batteux, F.; Borderie, D.; Chapron, C. Protein oxidative stress markers in peritoneal fluids of women with deep infiltrating endometriosis are increased. Hum. Reprod. 2015, 30, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Ngô, C.; Chéreau, C.; Nicco, C.; Weill, B.; Chapron, C.; Batteux, F. Reactive oxygen species controls endometriosis progression. Am. J. Pathol. 2009, 175, 225–234. [Google Scholar] [CrossRef]
- Szczepańska, M.; Koźlik, J.; Skrzypczak, J.; Mikołajczyk, M. Oxidative stress may be a piece in the endometriosis puzzle. Fertil. Steril. 2003, 79, 1288–1293. [Google Scholar] [CrossRef]
- Nasiri, N.; Moini, A.; Eftekhari-Yazdi, P.; Karimian, L.; Salman-Yazdi, R.; Arabipoor, A. Oxidative Stress Statues in Serum and Follicular Fluidof Women with Endometriosis. Cell J. 2017, 18, 582. [Google Scholar] [CrossRef]
- Prieto, L.; Quesada, J.F.; Cambero, O.; Pacheco, A.; Pellicer, A.; Codoceo, R.; Garcia-Velasco, J.A. Analysis of follicular fluid and serum markers of oxidative stress in women with infertility related to endometriosis. Fertil. Steril. 2012, 98, 126–130. [Google Scholar] [CrossRef]
- Goud, P.T.; Goud, A.P.; Joshi, N.; Puscheck, E.; Diamond, M.P.; Abu-Soud, H.M. Dynamics of nitric oxide, altered follicular microenvironment, and oocyte quality in women with endometriosis. Fertil. Steril. 2014, 102, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.W.; Schisterman, E.F.; Dey-Rao, R.; Browne, R.; Armstrong, D. Oxidative stress and endometriosis. Hum. Reprod. 2005, 20, 2014–2020. [Google Scholar] [CrossRef] [PubMed]
- McCord, J.M. Iron, Free Radicals, and Oxidative Injury. J. Nutr. 2004, 134, 3171S–3172S. [Google Scholar] [CrossRef] [PubMed]
- Defrère, S.; Lousse, J.C.; González-Ramos, R.; Colette, S.; Donnez, J.; Van Langendonckt, A. Potential involvement of iron in the pathogenesis of peritoneal endometriosis. Mol. Hum. Reprod. 2008, 14, 377–385. [Google Scholar] [CrossRef]
- Van Langendonckt, A.; Casanas-Roux, F.; Donnez, J. Iron overload in the peritoneal cavity of women with pelvic endometriosis. Fertil. Steril. 2002, 78, 712–718. [Google Scholar] [CrossRef]
- Lousse, J.C.; Defrère, S.; Van Langendonckt, A.; Gras, J.; González-Ramos, R.; Colette, S.; Donnez, J. Iron storage is significantly increased in peritoneal macrophages of endometriosis patients and correlates with iron overload in peritoneal fluid. Fertil. Steril. 2009, 91, 1668–1675. [Google Scholar] [CrossRef]
- Arumugam, K. Endometriosis: Endometriosis and infertility: Raised iron concentration in the peritoneal fluid and its effect on the acrosome reaction. Hum. Reprod. 1994, 9, 1153–1157. [Google Scholar] [CrossRef]
- Wölfler, M.M.; Meinhold-Heerlein, I.M.; Henkel, C.; Rath, W.; Neulen, J.; Maass, N.; Bräutigam, K. Reduced hemopexin levels in peritoneal fluid of patients with endometriosis. Fertil. Steril. 2013, 100, 777–781.e2. [Google Scholar] [CrossRef]
- Van Langendonckt, A.; Casanas-Roux, F.; Dolmans, M.M.; Donnez, J. Potential involvement of hemoglobin and heme in the pathogenesis of peritoneal endometriosis. Fertil. Steril. 2002, 77, 561–570. [Google Scholar] [CrossRef]
- Van Langendonckt, A.; Casanas-Roux, F.; Eggermont, J.; Donnez, J. Characterization of iron deposition in endometriotic lesions induced in the nude mouse model. Hum. Reprod. 2004, 19, 1265–1271. [Google Scholar] [CrossRef]
- Defrère, S.; Van Langendonckt, A.; Vaesen, S.; Jouret, M.; Ramos, R.G.; Gonzalez, D.; Donnez, J. Iron overload enhances epithelial cell proliferation in endometriotic lesions induced in a murine model. Hum. Reprod. 2006, 21, 2810–2816. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, H.; Halliwell, B. Damage to DNA by reactive oxygen and nitrogen species: Role in inflammatory disease and progression to cancer. Biochem. J. 1996, 313 Pt 1, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Yermilov, V.; Rubio, J.; Ohshima, H. Formation of 8-nitroguanine in DNA treated with peroxynitrite in vitro and its rapid removal from DNA by depurination. FEBS Lett. 1995, 376, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Canton, M.; Sánchez-Rodríguez, R.; Spera, I.; Venegas, F.C.; Favia, M.; Viola, A.; Castegna, A. Reactive Oxygen Species in Macrophages: Sources and Targets. Front. Immunol. 2021, 12, 4077. [Google Scholar] [CrossRef]
- Ragab, D.; Abbas, A.; Salem, R. Increased expression of IL-37 correlates with TNF-α levels and disease stage in endometriosis patients. Egypt. J. Med. Hum. Genet. 2022, 23, 72. [Google Scholar] [CrossRef]
- Senapati, S.; Clauw, D.; As-Sanie, S. The relationship between serum tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6) with pelvic pain symptoms in women with endometriosis. Fertil. Steril. 2009, 92, S112. [Google Scholar] [CrossRef]
- Milewski, Ł.; Dziunycz, P.; Barcz, E.; Radomski, D.; Roszkowski, P.I.; Korczak-Kowalska, G.; Kamiński, P.; Malejczyk, J. Increased levels of human neutrophil peptides 1, 2, and 3 in peritoneal fluid of patients with endometriosis: Association with neutrophils, T cells and IL-8. J. Reprod. Immunol. 2011, 91, 64–70. [Google Scholar] [CrossRef]
- Tariverdian, N.; Siedentopf, F.; Rücke, M.; Blois, S.M.; Klapp, B.F.; Kentenich, H.; Arck, P.C. Intraperitoneal immune cell status in infertile women with and without endometriosis. J. Reprod. Immunol. 2009, 80, 80–90. [Google Scholar] [CrossRef]
- Kwak, J.Y.; Park, S.W.; Kim, K.H.; Na, Y.J.; Lee, K.S. Modulation of neutrophil apoptosis by plasma and peritoneal fluid from patients with advanced endometriosis. Hum. Reprod. 2002, 17, 595–600. [Google Scholar] [CrossRef]
- Arici, A.; Seli, E.; Zeyneloglu, H.B.; Senturk, L.M.; Oral, E.; Olive, D.L. Interleukin-8 induces proliferation of endometrial stromal cells: A potential autocrine growth factor. J. Clin. Endocrinol. Metab. 1998, 83, 1201–1205. [Google Scholar] [CrossRef]
- Khan, K.N.; Masuzaki, H.; Fujishita, A.; Kitajima, M.; Sekine, I.; Ishimaru, T. Differential macrophage infiltration in early and advanced endometriosis and adjacent peritoneum. Fertil. Steril. 2004, 81, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Berbic, M.; Schulke, L.; Markham, R.; Tokushige, N.; Russell, P.; Fraser, I.S. Macrophage expression in endometrium of women with and without endometriosis. Hum. Reprod. 2009, 24, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Braun, P.D.; Ding, J.; Shen, J.; Rana, N.; Fernandez, B.B.; Dmowski, W.P. Relationship between apoptosis and the number of macrophages in eutopic endometrium from women with and without endometriosis. Fertil. Steril. 2002, 78, 830–835. [Google Scholar] [CrossRef] [PubMed]
- Takebayashi, A.; Kimura, F.; Kishi, Y.; Ishida, M.; Takahashi, A.; Yamanaka, A.; Wu, D.; Zheng, L.; Takahashi, K.; Suginami, H.; et al. Subpopulations of macrophages within eutopic endometrium of endometriosis patients. Am. J. Reprod. Immunol. 2015, 73, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Jolicoeur, C.; Boutouil, M.; Drouin, R.; Paradis, I.; Lemay, A.; Akoum, A. Increased expression of monocyte chemotactic protein-1 in the endometrium of women with endometriosis. Am. J. Pathol. 1998, 152, 125. [Google Scholar] [CrossRef]
- Na, Y.J.; Jin, J.O.; Lee, M.S.; Song, M.G.; Lee, K.S.; Kwak, J.Y. Peritoneal fluid from endometriosis patients switches differentiation of monocytes from dendritic cells to macrophages. J. Reprod. Immunol. 2008, 77, 63–74. [Google Scholar] [CrossRef]
- Chuang, P.C.; Wu, M.H.; Shoji, Y.; Tsai, S.J. Downregulation of CD36 results in reduced phagocytic ability of peritoneal macrophages of women with endometriosis. J. Pathol. 2009, 219, 232–241. [Google Scholar] [CrossRef]
- Montagna, P.; Capellino, S.; Villaggio, B.; Remorgida, V.; Ragni, N.; Cutolo, M.; Ferrero, S. Peritoneal fluid macrophages in endometriosis: Correlation between the expression of estrogen receptors and inflammation. Fertil. Steril. 2008, 90, 156–164. [Google Scholar] [CrossRef]
- Shao, J.; Zhang, B.; Yu, J.J.; Wei, C.Y.; Zhou, W.J.; Chang, K.K.; Yang, H.L.; Jin, L.P.; Zhu, X.Y.; Li, M.Q. Macrophages promote the growth and invasion of endometrial stromal cells by downregulating IL-24 in endometriosis. Reproduction 2016, 152, 673–682. [Google Scholar] [CrossRef]
- Chan, R.W.S.; Lee, C.L.; Ng, E.H.Y.; Yeung, W.S.B. Co-culture with macrophages enhances the clonogenic and invasion activity of endometriotic stromal cells. Cell Prolif. 2017, 50, e12330. [Google Scholar] [CrossRef]
- Shimizu, K.; Kamada, Y.; Sakamoto, A.; Matsuda, M.; Nakatsuka, M.; Hiramatsu, Y. High Expression of High-Mobility Group Box 1 in Menstrual Blood: Implications for Endometriosis. Reprod. Sci. 2017, 24, 1532–1537. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.H.; Chon, S.J.; Choi, Y.S.; Cho, S.; Lee, B.S.; Seo, S.K. Pathophysiology of Endometriosis: Role of High Mobility Group Box-1 and Toll-Like Receptor 4 Developing Inflammation in Endometrium. PLoS ONE 2016, 11, e0148165. [Google Scholar] [CrossRef] [PubMed]
- Santulli, P.; Borghese, B.; Chouzenoux, S.; Vaiman, D.; Borderie, D.; Streuli, I.; Goffinet, F.; De Ziegler, D.; Weill, B.; Batteux, F.; et al. Serum and peritoneal interleukin-33 levels are elevated in deeply infiltrating endometriosis. Hum. Reprod. 2012, 27, 2001–2009. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.E.; Monsanto, S.P.; Ahn, S.H.; Khalaj, K.; Fazleabas, A.T.; Young, S.L.; Lessey, B.A.; Koti, M.; Tayade, C. Interleukin-33 modulates inflammation in endometriosis. Sci. Rep. 2017, 7, 17903. [Google Scholar] [CrossRef]
- Heilier, J.F.; Nackers, F.; Verougstraete, V.; Tonglet, R.; Lison, D.; Donnez, J. Increased dioxin-like compounds in the serum of women with peritoneal endometriosis and deep endometriotic (adenomyotic) nodules. Fertil. Steril. 2005, 84, 305–312. [Google Scholar] [CrossRef]
- Simsa, P.; Mihalyi, A.; Schoeters, G.; Koppen, G.; Kyama, C.M.; Den Hond, E.M.; Fülöp, V.; D’Hooghe, T.M. Increased exposure to dioxin-like compounds is associated with endometriosis in a case-control study in women. Reprod. Biomed. Online 2010, 20, 681–688. [Google Scholar] [CrossRef]
- Hoon Kim, S.; Chun, S.; Yeon Jang, J.M.; Dong Chae, H.; Kim, C.-H.; Moon Kang, B. Increased plasma levels of phthalate esters in women with advanced-stage endometriosis: A prospective case-control study. Fertil. Steril. 2011, 95, 357–359. [Google Scholar] [CrossRef]
- Rier, S.E.; Turner, W.E.; Martin, D.C.; Morris, R.; Lucier, G.W.; Clark, G.C. Serum Levels of TCDD and Dioxin-like Chemicals in Rhesus Monkeys Chronically Exposed to Dioxin: Correlation of Increased Serum PCB Levels with Endometriosis. Toxicol. Sci. 2001, 59, 147–159. [Google Scholar] [CrossRef]
- Porpora, M.G.; Medda, E.; Abballe, A.; Bolli, S.; De Angelis, I.; di Domenico, A.; Ferro, A.; Ingelido, A.M.; Maggi, A.; Panici, P.B.; et al. Endometriosis and organochlorinated environmental pollutants: A case-control study on Italian women of reproductive age. Environ. Health Perspect. 2009, 117, 1070–1075. [Google Scholar] [CrossRef]
- Huang, P.-C.; Tsai, E.-M.; Li, W.-F.; Liao, P.-C.; Chung, M.-C.; Wang, Y.-H.; Wang, S.-L. Association between phthalate exposure and glutathione S-transferase M1 polymorphism in adenomyosis, leiomyoma and endometriosis. Hum. Reprod. 2010, 25, 986–994. [Google Scholar] [CrossRef]
- Buck Louis, G.M.; Sundaram, R.; Sweeney, A.M.; Schisterman, E.F.; Maisog, J.; Kannan, K. Urinary bisphenol A, phthalates, and couple fecundity: The Longitudinal Investigation of Fertility and the Environment (LIFE) Study. Fertil. Steril. 2014, 101, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Eskenazi, B.; Mocarelli, P.; Warner, M.; Samuels, S.; Vercellini, P.; Olive, D.; Needham, L.L.; Patterson, D.G.; Brambilla, P.; Gavoni, N.; et al. Serum dioxin concentrations and endometriosis: A cohort study in Seveso, Italy. Environ. Health Perspect. 2002, 110, 629–634. [Google Scholar] [CrossRef]
- Fierens, S.; Mairesse, H.; Heilier, J.F.; de Burbure, C.; Focant, J.F.; Eppe, G.; de Pauw, E.; Bernard, A. Dioxin/polychlorinated biphenyl body burden, diabetes and endometriosis: Findings in a population-based study in Belgium. Biomarkers 2003, 8, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Zeitoun, K.M.; Bulun, S.E. Aromatase: A key molecule in the pathophysiology of endometriosis and a therapeutic target. Fertil. Steril. 1999, 72, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Noble, L.S.; Takayama, K.; Zeitoun, K.M.; Putman, J.M.; Johns, D.A.; Hinshelwood, M.M.; Agarwal, V.R.; Zhao, Y.; Carr, B.R.; Bulun, S.E. Prostaglandin E2 stimulates aromatase expression in endometriosis-derived stromal cells. J. Clin. Endocrinol. Metab. 1997, 82, 600–606. [Google Scholar] [CrossRef]
- Mori, T.; Ito, F.; Koshiba, A.; Kataoka, H.; Takaoka, O.; Okimura, H.; Khan, K.N.; Kitawaki, J. Local estrogen formation and its regulation in endometriosis. Reprod. Med. Biol. 2019, 18, 305–311. [Google Scholar] [CrossRef]
- Monsivais, D.; Dyson, M.T.; Yin, P.; Coon, J.S.; Navarro, A.; Feng, G.; Malpani, S.S.; Ono, M.; Ercan, C.M.; Wei, J.J.; et al. ERβ- and prostaglandin E2-regulated pathways integrate cell proliferation via Ras-like and estrogen-regulated growth inhibitor in endometriosis. Mol. Endocrinol. 2014, 28, 1304–1315. [Google Scholar] [CrossRef]
- Burns, K.A.; Thomas, S.Y.; Hamilton, K.J.; Young, S.L.; Cook, D.N.; Korach, K.S. Early Endometriosis in Females Is Directed by Immune-Mediated Estrogen Receptor α and IL-6 Cross-Talk. Endocrinology 2018, 159, 103–118. [Google Scholar] [CrossRef]
- Han, S.J.; Jung, S.Y.; Wu, S.P.; Hawkins, S.M.; Park, M.J.; Kyo, S.; Qin, J.; Lydon, J.P.; Tsai, S.Y.; Tsai, M.J.; et al. Estrogen Receptor β Modulates Apoptosis Complexes and the Inflammasome to Drive the Pathogenesis of Endometriosis. Cell 2015, 163, 960–974. [Google Scholar] [CrossRef]
- Huhtinen, K.; Desai, R.; Ståhle, M.; Salminen, A.; Handelsman, D.J.; Perheentupa, A.; Poutanen, M. Endometrial and Endometriotic Concentrations of Estrone and Estradiol Are Determined by Local Metabolism Rather than Circulating Levels. J. Clin. Endocrinol. Metab. 2012, 97, 4228. [Google Scholar] [CrossRef]
- Othman, E.R.; Markeb, A.A.; Khashbah, M.Y.; Abdelaal, I.I.; ElMelegy, T.T.; Fetih, A.N.; Van der Houwen, L.E.; Lambalk, C.B.; Mijatovic, V. Markers of Local and Systemic Estrogen Metabolism in Endometriosis. Reprod. Sci. 2021, 28, 1001–1011. [Google Scholar] [CrossRef]
- Saliminejad, K.; Saket, M.; Kamali, K.; Memariani, T.; Khorram Khorshid, H.R. DNA Repair Gene XRCC1 and XRCC4 Variations and Risk of Endometriosis: An Association Study. Gynecol. Obstet. Invest. 2015, 80, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.Y.; Bau, D.T.; Chang, C.C.; Tsai, C.H.; Chen, C.P.; Tsai, F.J. XRCC4 codon 247*A and XRCC4 promoter-1394*T related genotypes but not XRCC4 intron 3 gene polymorphism are associated with higher susceptibility for endometriosis. Mol. Reprod. Dev. 2008, 75, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Attar, R.; Cacina, C.; Sozen, S.; Attar, E.; Agachan, B. DNA repair genes in endometriosis. Genet. Mol. Res. 2010, 9, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Kiyomizu, M.; Kitawaki, J.; Obayashi, H.; Ohta, M.; Koshiba, H.; Ishihara, H.; Honjo, H. Association of Two Polymorphisms in the Peroxisome Proliferator-Acativated Receptor-γ Gene With Adenomyosis, Endometriosis, and Leiomyomata in Japanese Women. J. Soc. Gynecol. Investig. 2006, 13, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Dogan, S.; Machicao, F.; Wallwiener, D.; Haering, H.U.; Diedrich, K.; Hornung, D. Association of peroxisome proliferator-activated receptor gamma 2 Pro-12-Ala polymorphism with endometriosis. Fertil. Steril. 2004, 81, 1411–1413. [Google Scholar] [CrossRef]
- Vietri, M.T.; Molinari, A.M.; Iannella, I.; Cioffi, M.; Bontempo, P.; Ardovino, M.; Scaffa, C.; Colacurci, N.; Cobellis, L. Arg72Pro p53 polymorphism in Italian women: No association with endometriosis. Fertil. Steril. 2007, 88, 1468–1469. [Google Scholar] [CrossRef]
- Lattuada, D.; Viganò, P.; Somigliana, E.; Abbiati, A.; Candiani, M.; Di Blasio, A.M. Analysis of the codon 72 polymorphism of the TP53 gene in patients with endometriosis. Mol. Hum. Reprod. 2004, 10, 651–654. [Google Scholar] [CrossRef]
- Gallegos-Arreola, M.P.; Figuera-Villanueva, L.E.; Puebla-Pérez, A.M.; Montoya-Fuentes, H.; Suarez-Rincon, A.E.; Zúñiga-González, G.M. Association of TP53 gene codon 72 polymorphism with endometriosis in Mexican women. Genet. Mol. Res. 2012, 11, 1401–1408. [Google Scholar] [CrossRef]
- Chang, C.C.; Hsieh, Y.Y.; Tsai, F.J.; Tsai, C.H.; Tsai, H.D.; Lin, C.C. The proline form of p53 codon 72 polymorphism is associated with endometriosis. Fertil. Steril. 2002, 77, 43–45. [Google Scholar] [CrossRef]
- Hsieh, Y.Y.; Lin, C.S. P53 codon 11, 72, and 248 gene polymorphisms in endometriosis. Int. J. Biol. Sci. 2006, 2, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Nikbakht Dastjerdi, M.; Aboutorabi, R.; Eslami Farsani, B. Association of TP53 gene codon 72 polymorphism with endometriosis risk in Isfahan. Iran. J. Reprod. Med. 2013, 11, 473–478. [Google Scholar] [PubMed]
- Hussain, R.; Khaliq, S.; Raza, S.M.; Khaliq, S.; Lone, K.P. Association of TP53 codon 72 polymorphism in women suffering from endometriosis from Lahore, Pakistan. J. Pak. Med. Assoc. 2018, 68, 224–230. [Google Scholar] [PubMed]
- Monteiro, M.; Vilas Boas, D.; Gigliotti, C.; Salvadori, D. Association among XRCC1, XRCC3, and BLHX gene polymorphisms and chromosome instability in lymphocytes from patients with endometriosis and ovarian cancer. Genet. Mol. Res. 2014, 13, 636–648. [Google Scholar] [CrossRef]
- Hsieh, Y.Y.; Chang, C.C.; Chen, S.Y.; Chen, C.P.; Lin, W.H.; Tsai, F.J. XRCC1 399*Arg-related genotype and allele, but not XRCC1 His107Arg, XRCC1 Trp194Arg, KCNQ2, AT1R, and hOGG1 polymorphisms, are associated with higher susceptibility of endometriosis. Gynecol. Endocrinol. 2012, 28, 305–309. [Google Scholar] [CrossRef]
- Shen, T.C.; Tsai, C.W.; Chang, W.S.; Wang, Y.C.; Hsu, H.M.; Li, H.T.; Gu, J.; Bau, D.T. Genetic variants in the nucleotide excision repair genes are associated with the risk of developing endometriosis. Biol. Reprod. 2019, 101, 928–937. [Google Scholar] [CrossRef]
- Martini, M.; Ciccarone, M.; Garganese, G.; Maggiore, C.; Evangelista, A.; Rahimi, S.; Zannoni, G.; Vittori, G.; Larocca, L.M. Possible involvement of hMLH1, p16INK4a and PTEN in the malignant transformation of endometriosis. Int. J. Cancer 2002, 102, 398–406. [Google Scholar] [CrossRef]
- Matta, J.L.; Flores, I.; Morales, L.M.; Monteiro, J.; Alvarez-Garriga, C.; Bayona, M. Women with endometriosis have a higher DNA repair capacity and diminished breast cancer risk. Mol. Cancer Biol. 2013, 1. [Google Scholar] [CrossRef]
- Talbi, S.; Hamilton, A.E.; Vo, K.C.; Tulac, S.; Overgaard, M.T.; Dosiou, C.; Le Shay, N.; Nezhat, C.N.; Kempson, R.; Lessey, B.A.; et al. Molecular phenotyping of human endometrium distinguishes menstrual cycle phases and underlying biological processes in normo-ovulatory women. Endocrinology 2006, 147, 1097–1121. [Google Scholar] [CrossRef]
- Burney, R.O.; Talbi, S.; Hamilton, A.E.; Kim, C.V.; Nyegaard, M.; Nezhat, C.R.; Lessey, B.A.; Giudice, L.C. Gene expression analysis of endometrium reveals progesterone resistance and candidate susceptibility genes in women with endometriosis. Endocrinology 2007, 148, 3814–3826. [Google Scholar] [CrossRef]
- Hever, A.; Roth, R.B.; Hevezi, P.; Marin, M.E.; Acosta, J.A.; Acosta, H.; Rojas, J.; Herrera, R.; Grigoriadis, D.; White, E.; et al. Human endometriosis is associated with plasma cells and overexpression of B lymphocyte stimulator. Proc. Natl. Acad. Sci. USA 2007, 104, 12451–12456. [Google Scholar] [CrossRef] [PubMed]
- Tamaresis, J.S.; Irwin, J.C.; Goldfien, G.A.; Rabban, J.T.; Burney, R.O.; Nezhat, C.; DePaolo, L.V.; Giudice, L.C. Molecular classification of endometriosis and disease stage using high-dimensional genomic data. Endocrinology 2014, 155, 4986–4999. [Google Scholar] [CrossRef] [PubMed]
- Poli-Neto, O.B.; Carlos, D.; Favaretto, A.; Rosa-e-Silva, J.C.; Meola, J.; Tiezzi, D. Eutopic endometrium from women with endometriosis and chlamydial endometritis share immunological cell types and DNA repair imbalance: A transcriptome meta-analytical perspective. J. Reprod. Immunol. 2021, 145, 103307. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munshi, I.; Sachdeva, G. Genomic Insults and their Redressal in the Eutopic Endometrium of Women with Endometriosis. Reprod. Med. 2023, 4, 74-88. https://doi.org/10.3390/reprodmed4020009
Munshi I, Sachdeva G. Genomic Insults and their Redressal in the Eutopic Endometrium of Women with Endometriosis. Reproductive Medicine. 2023; 4(2):74-88. https://doi.org/10.3390/reprodmed4020009
Chicago/Turabian StyleMunshi, Itti, and Geetanjali Sachdeva. 2023. "Genomic Insults and their Redressal in the Eutopic Endometrium of Women with Endometriosis" Reproductive Medicine 4, no. 2: 74-88. https://doi.org/10.3390/reprodmed4020009
APA StyleMunshi, I., & Sachdeva, G. (2023). Genomic Insults and their Redressal in the Eutopic Endometrium of Women with Endometriosis. Reproductive Medicine, 4(2), 74-88. https://doi.org/10.3390/reprodmed4020009