
On Deterministic and Stochastic Multiple Pathogen Epidemic Models


Abstract
:1. Introduction
2. Materials and Methods
2.1. Derivation of Stochastic Epidemic Model with Two Pathogen Strains
2.2. The Deterministic Model
2.3. Simulation Using the Euler–Maruyama Method
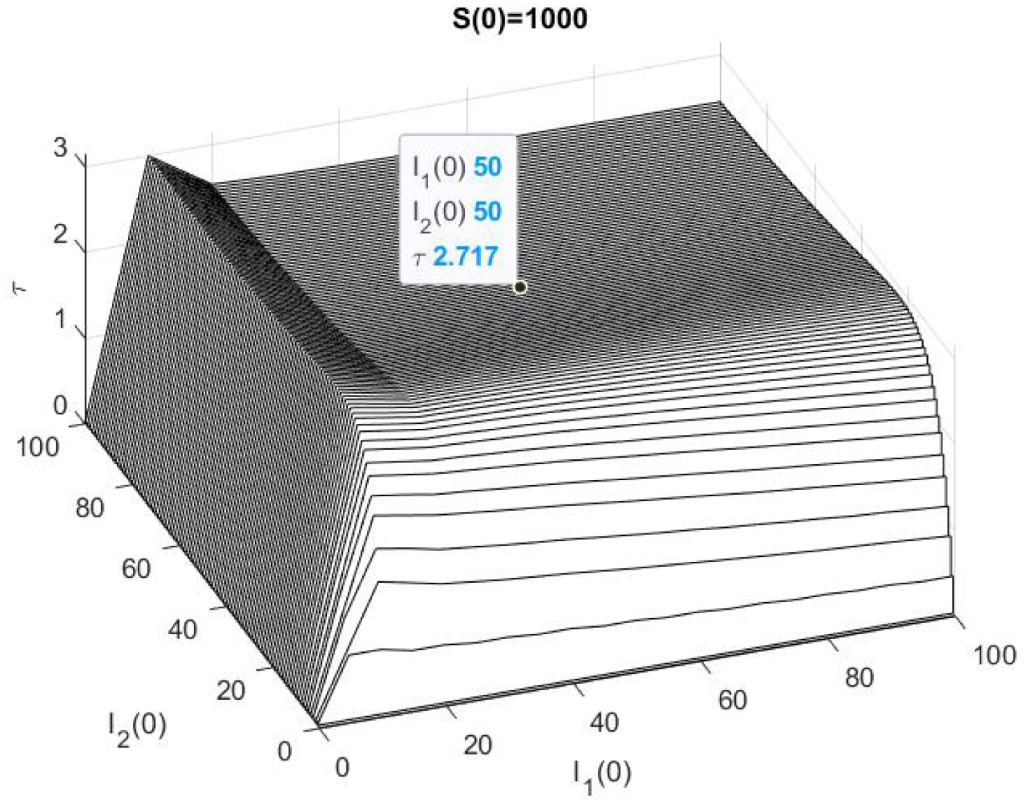
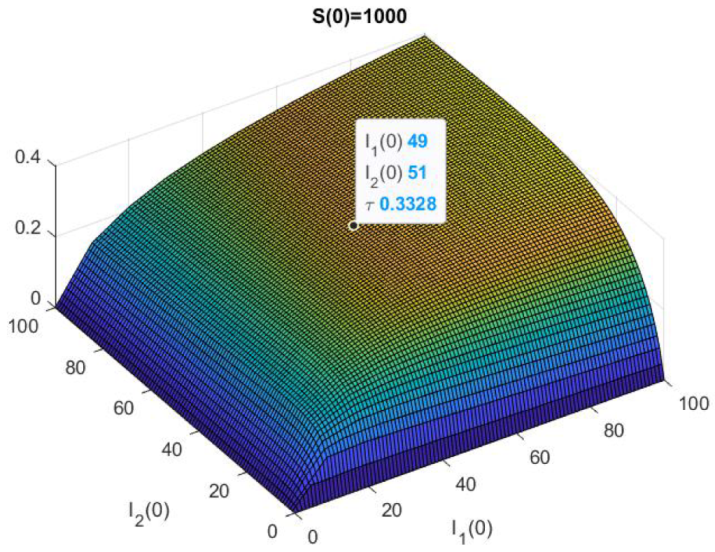
2.4. The Mean Persistence Time for the Model
2.5. Variational Formulation
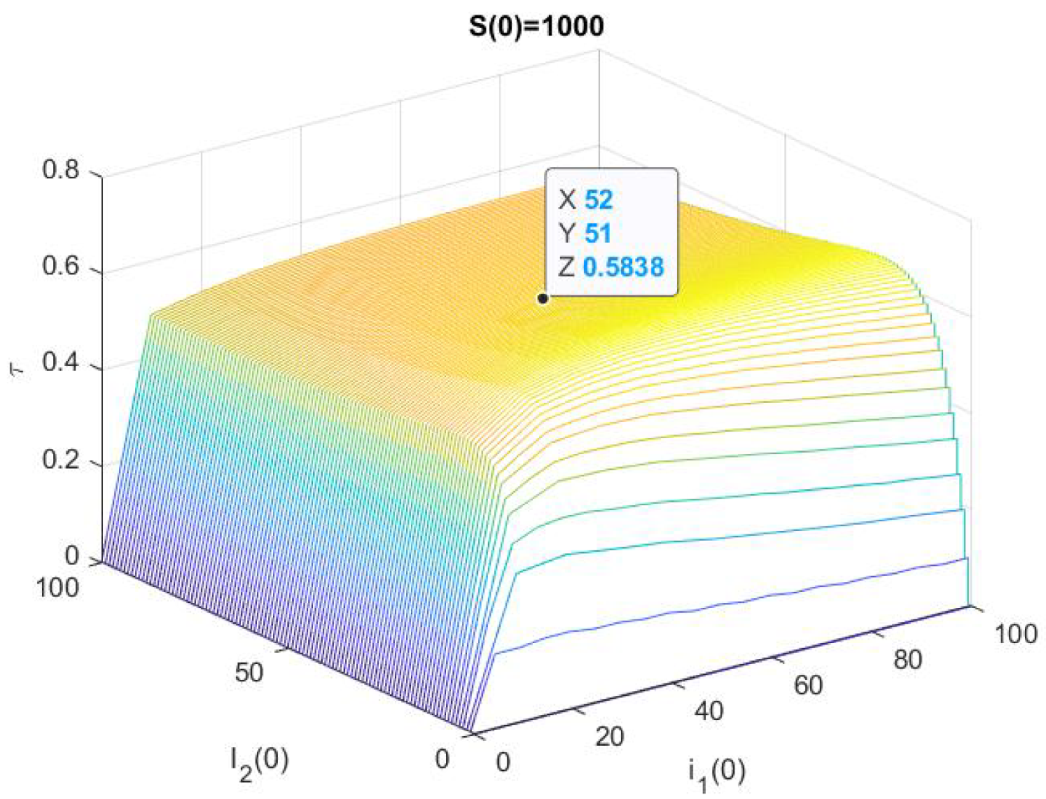
3. Numerical Results
4. Conclusions and Discution
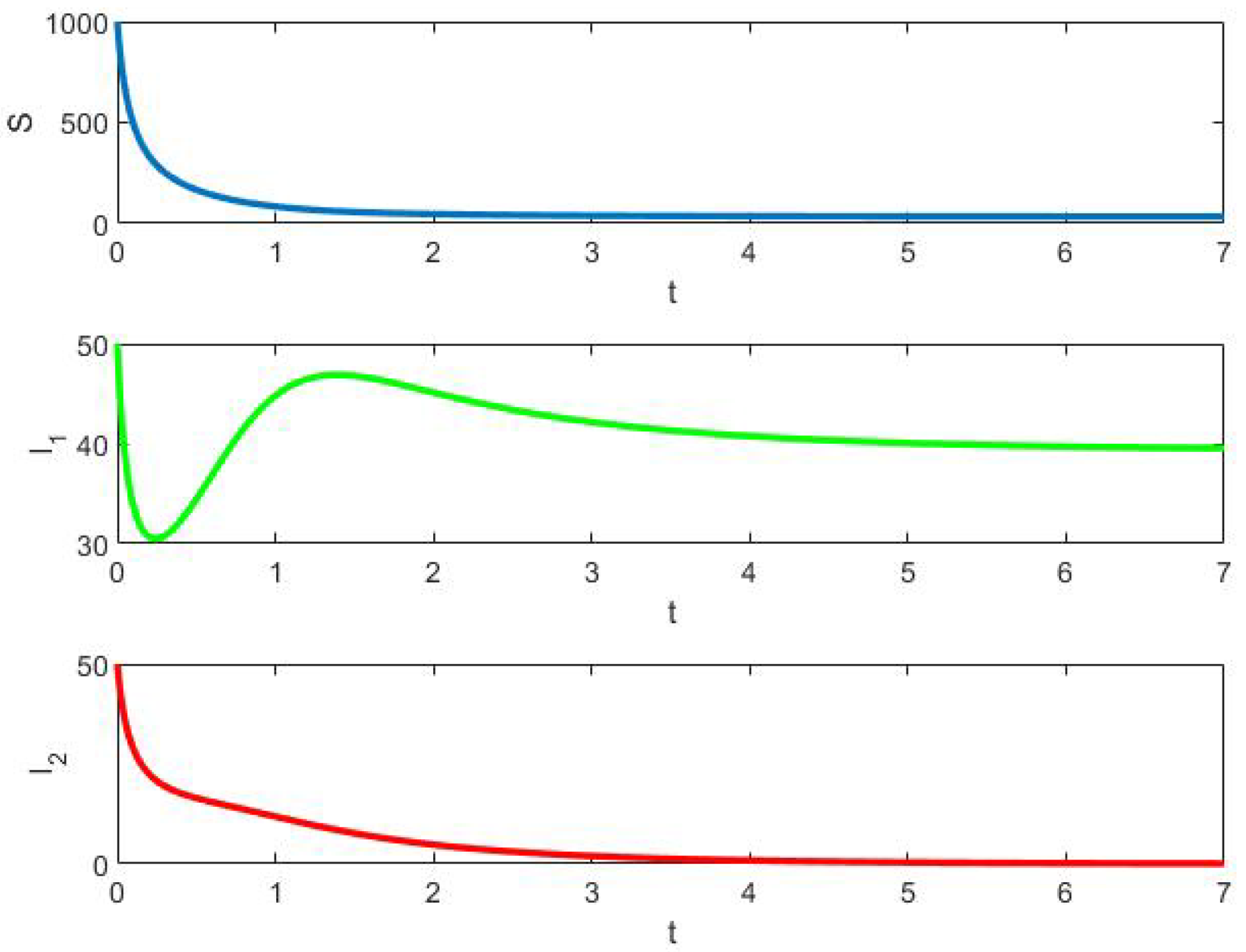
- Example 1: In this case, competitive exclusion occurs because in the deterministic model , while the stochastic model disappears somewhat more quickly.
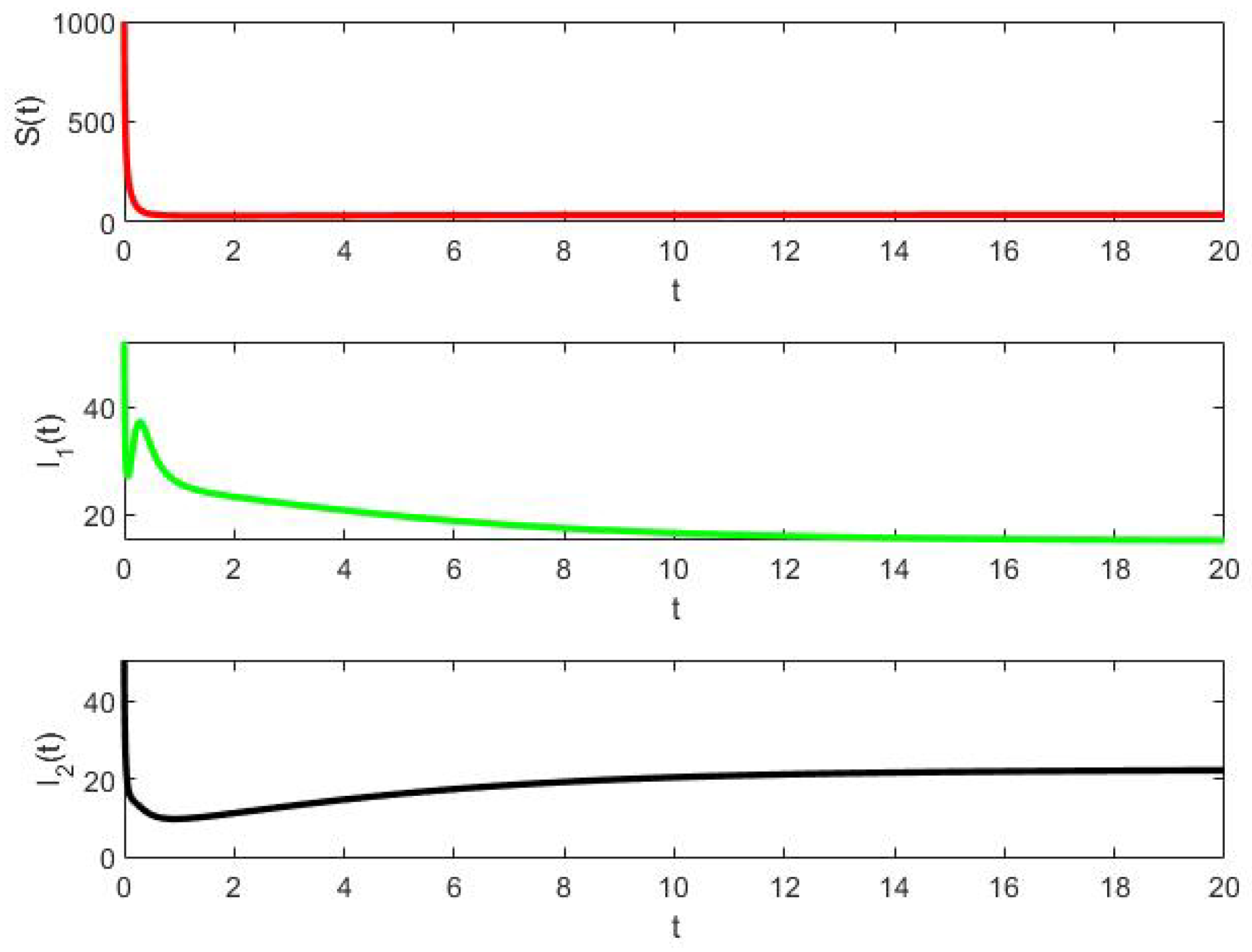
- Example 2: In this example, with vertical transmission of both strains, the deterministic solution cycles closer and closer while the stochastic solution is extinguished very quickly. The difference in the asymptotic behavior of deterministic and stochastic is very important.
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, J. Mathematical Biology I: An Introduction, 3rd ed.; Springer: Berlin, Germany, 2002. [Google Scholar]
- Daley, D.; Gani, J. Epidemid Modelling: An Introduction; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Hethcote, H. The Mathematics of Infection Diseases. SIAM Rev. 2000, 42, 599–653. [Google Scholar] [CrossRef] [Green Version]
- Brauer, F.; Castillo-Chaves, C. Mathematical Models in Population Biology and Epidemiology; Springer: Berlin, Germany, 2012. [Google Scholar]
- Martcheva, M. An Introductio to Mathematical Epidemiology; Springer: Berlin, Germany, 2015. [Google Scholar]
- Brauer, F.; Castillo-Chaves, C.; Feng, Z. Mathematical Models in Epidemiology; Springer: Berlin, Germany, 2019. [Google Scholar]
- Jiang, D.; Yu, J.; Ji, C.; Shi, N. Asymptotic behavior of global positive solution to a stochastic SIR model. Math. Comput. Model. 2011, 54, 221–232. [Google Scholar] [CrossRef]
- Zhao, Y.; Jiang, D. The threshold of a stochastic SIRS epidemic model with saturated incidence. Appl. Math. Latter 2014, 34, 90–93. [Google Scholar] [CrossRef]
- Rajasekar, S.; Pitchaimani, M. Ergodic stationary distribution and extinction of a stochastic SIRS epidemic model with logistic growth and nonlinear incidence. Appl. Math. Comput. 2020, 337, 125–143. [Google Scholar] [CrossRef]
- Liu, Q.; Jiang, D. Threshold behavior in a stochastic SIR epidemic model with Logistic birth. Phyisica A 2020, 540, 123–135. [Google Scholar] [CrossRef]
- Allen, E. Modeling with Itô Stochastic Differential Equations; Springer: Berlin, Germany, 2007. [Google Scholar]
- Allen, L. An Introduction to Stochastic Proceses with Applications to Biology, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Vadillo, F. Comparing stochastic Lotka-Volterra predator-prey models. Appl. Math. Comput. 2019, 360, 181–189. [Google Scholar] [CrossRef]
- Skvortsov, A.; Ristic, B.; Kamenev, A. Predicting population extinction from early observations of the Lotka-Volterra system. Appl. Math. Comput. 2018, 320, 371–379. [Google Scholar] [CrossRef]
- Castillo-Chavez, C.; Huang, W.; Li, J. Competitive exclusion and coexistence of multiple strains in an SIS STD model. SIAM J. Appl. Math. 1999, 59, 1790–1811. [Google Scholar] [CrossRef]
- Kirupahara, N.; Allen, L. Coexistence of Multiple Pathogen Strains in Stochastic Epidemic Models with Density-Dependent Mortality. Bull. Math. Biol. 2004, 66, 841–864. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.; Kirupahara, N. Asymptotic Dynamincs of Determinicsand Stocahstic Epidemic Models witn Multiple Pathogens. Int. J. Numer. Anal. Model. 2005, 2, 329–344. [Google Scholar]
- De la Hoz, F.; Vadillo, F. A mean extinction-time estimate for a stochastic Lotka-Volterra predator-prey model. Appl. Math. Comput. 2012, 219, 170–179. [Google Scholar] [CrossRef]
- De la Hoz, F.; Doubova, A.; Vadillo, F. Persistence-time Estimation for some Stochastic SIS Epidemic Models. Discret. Countinous Dyn. Syst. Ser. B 2015, 20, 2933–2947. [Google Scholar] [CrossRef]
- Gilbarg, D.; Trudinger, N. Elliptic Partial Differential Equations of Second Order, 2nd ed.; Springer: Berlin, Germany, 1983. [Google Scholar]
- Smoller, J. Shock Waves and Reaction-Diffusion Equations; Springer: Berlin, Germany, 1983. [Google Scholar]
- Stroock, D. Lectures on Stochastic Analysssi: Diffusion Theory; Cambridge University Press: Cambridge, UK, 1987. [Google Scholar]
- Hecht, F. New development in freefem++. J. Numer. Math. 2012, 20, 251–265. [Google Scholar] [CrossRef]
- Kloeden, P.; Platen, P.E. Numerical Solution of Stochastic Differential Equations; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Zhang, Z.; Karniadakis, G. Numerical Methods for Stochastic Partial Differential Equations with White Noise; Springer: Berlin, Germany, 2017. [Google Scholar]
- Higham, D.J.; Kloeden, P.E. An Introduction to the Numerical Simulation of Stochastic Differential Equations; Society for Industial and Applied Mathematics: Philadelphia, PA, USA, 2021. [Google Scholar]
- Higham, D.J. An Algorithmic Introduction to Numerical Simulation of Stochastic Differential Equations. SIAM Rev. 2001, 43, 525–546. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Changes | Probabilities |
|---|---|
| Zeros | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| S | ∗ | ∗ | ∗ | 0 | ∗ | 0 |
| ∗ | 0 | 0 | ∗ | ∗ | ∗ | |
| 0 | ∗ | ∗ | ∗ | ∗ | ∗ |
| Initial Point | Number of Stops | Mean | Std | |
|---|---|---|---|---|
| (1000, 50, 50) | 0 | |||
| 120 | 1.6929 | 0.8579 | ||
| 9880 | 2.5880 | 1.0005 |
| Initial Point | Number of Stops | Mean | Std | |
|---|---|---|---|---|
| (1000, 49, 51) | 4109 | 0.5716 | 0.0955 | |
| 14 | ||||
| 5877 | 0.3276 | 0.1237 |
| Initial Point | Number of Stops | Mean | Std | |
|---|---|---|---|---|
| (1000, 52, 51) | 0 | |||
| 9543 | 0.9348 | 0.3894 | ||
| 457 | 0.5024 | 0.3119 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vadillo, F. On Deterministic and Stochastic Multiple Pathogen Epidemic Models. Epidemiologia 2021, 2, 325-337. https://doi.org/10.3390/epidemiologia2030025
Vadillo F. On Deterministic and Stochastic Multiple Pathogen Epidemic Models. Epidemiologia. 2021; 2(3):325-337. https://doi.org/10.3390/epidemiologia2030025
Chicago/Turabian StyleVadillo, Fernando. 2021. "On Deterministic and Stochastic Multiple Pathogen Epidemic Models" Epidemiologia 2, no. 3: 325-337. https://doi.org/10.3390/epidemiologia2030025
APA StyleVadillo, F. (2021). On Deterministic and Stochastic Multiple Pathogen Epidemic Models. Epidemiologia, 2(3), 325-337. https://doi.org/10.3390/epidemiologia2030025