
The Counteraction of Cultivated Cistus creticus L. (Rock Rose) Plants to the Strain Imposed by a Long-Term Exposure to Non-Ionizing Radiation and the Role of DDC
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Exposure Setup
| CAGE | Average | Maximum |
| Control | 73.67 mV/m | 548 mV/m |
| Exposed | 2.072 V/m | 11.32 V/m |
2.2. Microscopy
2.3. Pigments Protocol
2.4. Protein Extraction and Determination of Protein Concentration
2.5. SDS-Polyacrylamide Gel Electrophoresis and Immunoblotting
2.6. Antibodies
2.7. Quantification
2.8. Total Reactive Oxygen Species (ROS) Estimation
2.9. Total Phenolic Content
2.10. Statistical Analysis
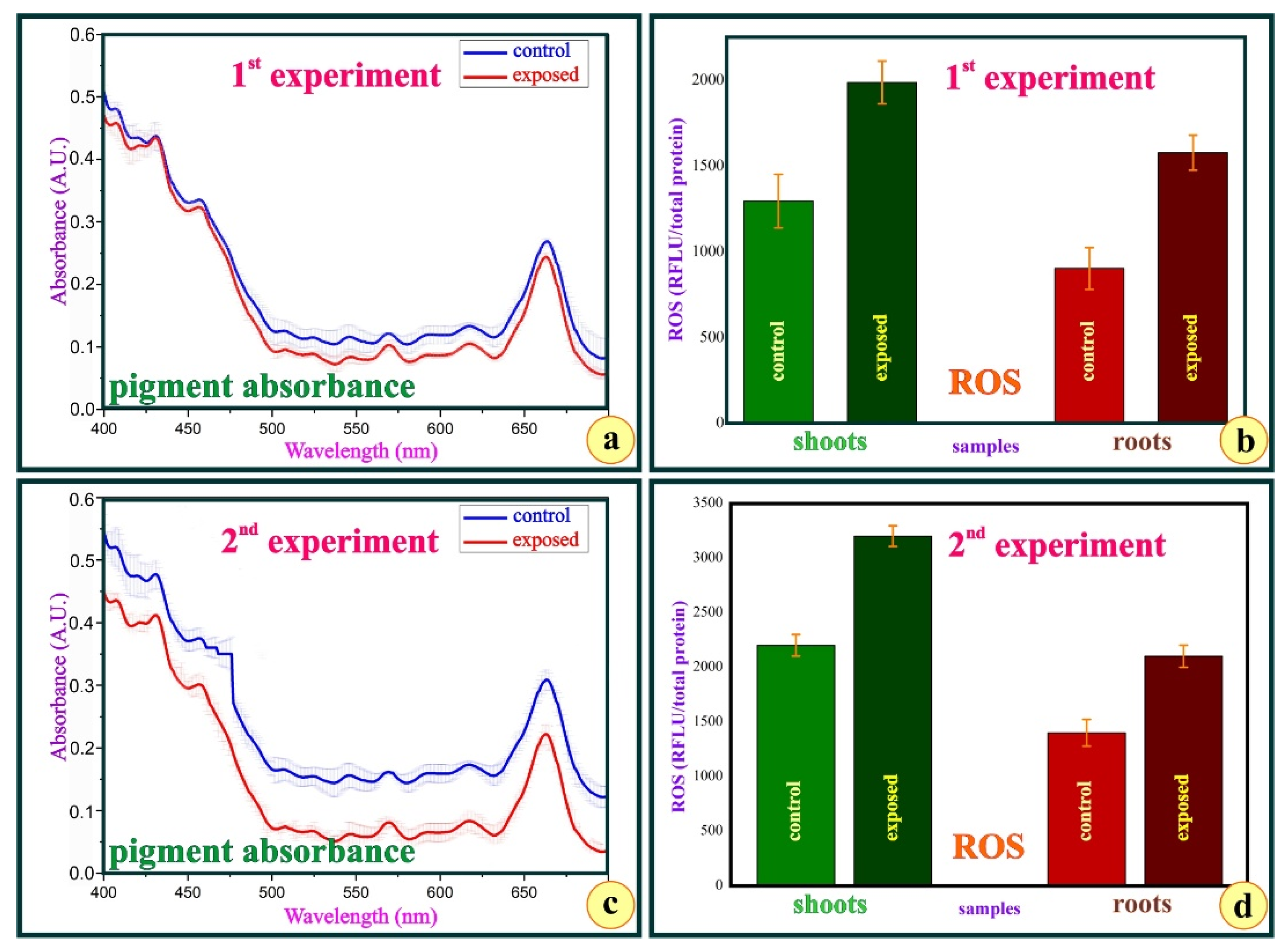
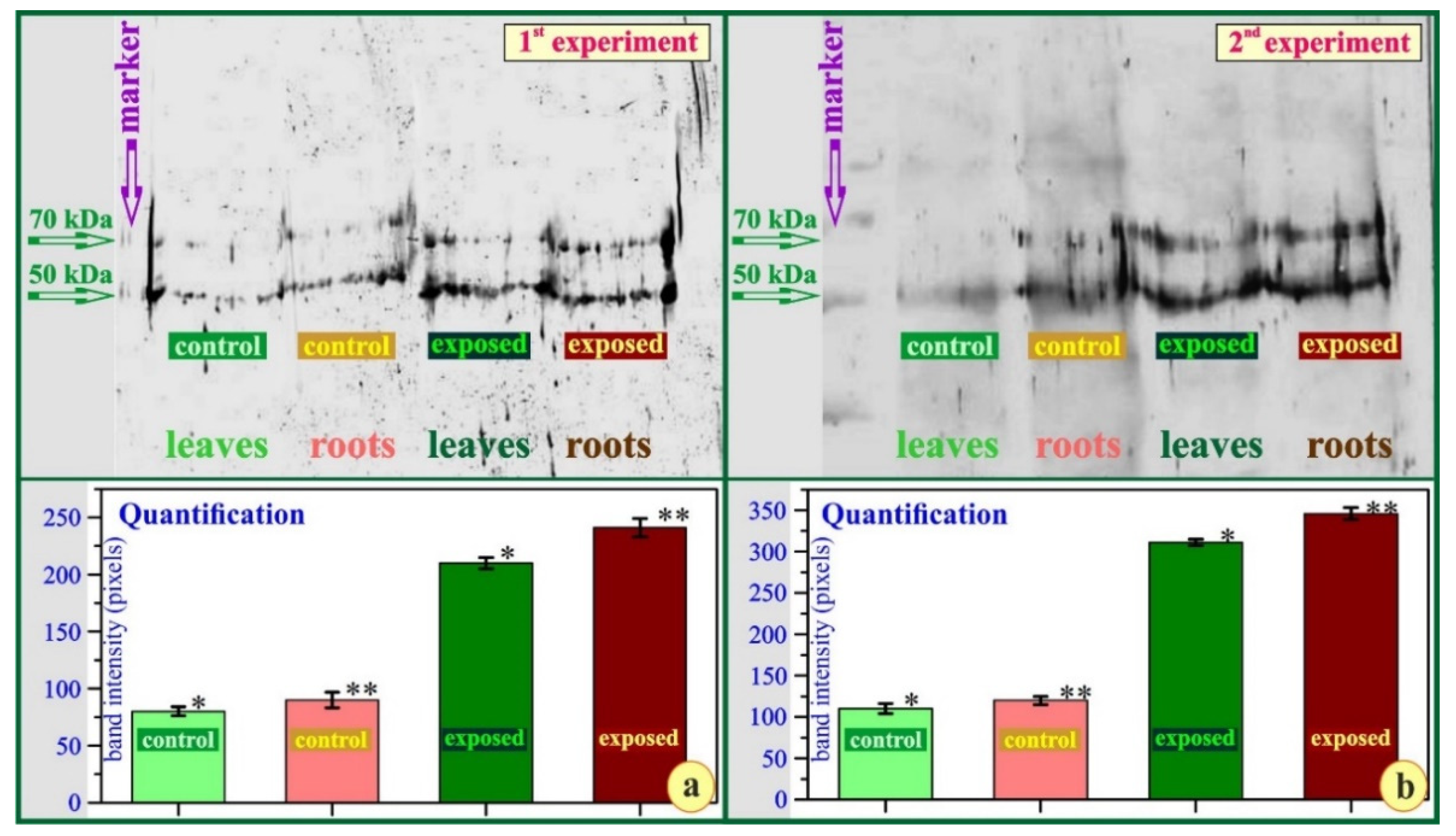
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitrakos, K. A theory of Mediterranean plant life. Oecologia Plant 1980, 1, 245–252. [Google Scholar]
- Mooney, H.A.; Parsons, D.J. Structure and Function in the California Chaparral—An Example from San Dimas. In Mediterranean Type Ecosystems. Origin and Structure; di Castri, F., Mooney, H.A.M., Eds.; Springer: Berlin, Germany, 1973; pp. 83–112. [Google Scholar]
- Christodoulakis, N.S.; Kogia, D.; Mavroeidi, D.; Fasseas, C. Anatomical and cytochemical investigation on the leaf of Teucrium polium L., a pharmaceutical shrub of the Greek phryganic formations. J. Biol. Res. 2010, 14, 199–209. [Google Scholar]
- Daget, P. Le bioclimat mediterraneen: Caracteres generaux, modes de caracterisation. Vegetatio 1977, 34, 1–20. [Google Scholar] [CrossRef]
- Nahal, I. The Mediterranean Climate from a Biological Viewpoint. In Ecosystems of the World, Mediterranean-Type Shrublands; de Castri, F., Goodall, D.W., Specht, R.L., Eds.; Elsevier Scientific Publishing Company: Amsterdam, The Netherlands, 1981; pp. 63–86. [Google Scholar]
- Hutchinson, J. The Families of Flowering Plants: Arranged According to a New System Based on Their Probable Phylogeny, 3rd ed.; Clarendon: Oxford, UK, 1973; ISBN 0-19-854377-8. [Google Scholar]
- Edlmann, M.L.A.; de Luca, L.; Lazzeri, S. Atlante Anatomico Degli Alberi ed Arbusti Della Macchia Mediterranea; (Relazioni e Monografie Agrarie Subtropicali e Tropicali, Nuova Serie 114.); Istituto Agronomico per l’ Oltremare: Firenze, Italy, 1994. [Google Scholar]
- De Micco, V.; Arena, C.; Vitale, L.; Aronne, G.; De Santo, A.V. Anatomy and photochemical behaviour of Mediterranean Cistus incanus winter leaves under natural outdoor and warmer indoor conditions. Botany 2011, 89, 677–688. [Google Scholar] [CrossRef]
- Dioscurides, A.C. De Materia Medica; Militos Publications: Athens, Greece, 2001. [Google Scholar]
- De Andrés, A.I.; Gómez-Serranillos, M.P.; Iglesias, I.; Villar, A.M. Effects of extract of Cistus populifolius L. on the central nervous system. Phytother. Res. 1999, 13, 575–579. [Google Scholar] [CrossRef]
- Küpeli, E.; Yesilada, E. Flavonoids with anti-inflammatory and antinociceptive activity from Cistus laurifolius L. leaves through bioassay-guided procedures. J. Ethnopharmacol. 2007, 112, 524–530. [Google Scholar] [CrossRef]
- Barrajõn-Catalán, E.; Fernández-Arroyo, S.; Roldán, C.; Guillén, E.; Saura, D.; Segura-Carretero, A.; Micol, V. A systematic study of the polyphenolic composition of aqueous extracts deriving from several Cistus genus species: Evolutionary relationship. Phytochem. Anal. 2011, 22, 303–312. [Google Scholar] [CrossRef]
- Barrajón-Catalán, E.; Fernández-Arroyo, S.; Saura, D.; Guillén, E.; Fernández-Gutiérrez, A.; Segura-Carretero, A.; Micol, V. Cistaceae aqueous extracts containing ellagitannins show antioxidant and antimicrobial capacity, and cytotoxic activity against human cancer cells. Food Chem. Toxicol. 2010, 48, 2273–2282. [Google Scholar] [CrossRef]
- Vitali, F.; Pennisi, G.; Attaguile, G.; Savoca, F.; Tita, B. Antiproliferative and cytotoxic activity of extracts from Cistus incanus L. and Cistus monspeliensis L. on human prostate cell lines. Nat. Prod. Res. 2011, 25, 188–202. [Google Scholar] [CrossRef]
- Droebner, K.; Ehrhardt, C.; Poetter, A.; Ludwig, S.; Planz, O. CYSTUS052, a polyphenol-rich plant extract, exerts anti-influenza virus activity in mice. Antivir. Res. 2007, 76, 1–10. [Google Scholar] [CrossRef]
- Chon, H. Medicinal herbs and plant extracts for influenza: Bioactivity, mechanism of anti-influenza effects, and modulation of immune responses. Stud. Nat. Prod. Chem. 2012, 38, 305–323. [Google Scholar]
- Aronne, G.; De Micco, V. Seasonal dimorphism in the Mediterranean Cistus incanus L. subsp. incanus. Ann. Bot. 2001, 87, 789–794. [Google Scholar] [CrossRef]
- Gülz, P.-G.; Herrmann, T.; Hangst, K. Leaf trichomes in the genus Cistus. Flora 1996, 191, 85–104. [Google Scholar] [CrossRef]
- Christodoulakis, N.S.; Georgoudi, M.; Fasseas, C. Leaf structure of Cistus creticus L. (rock rose), a medicinal plant widely used in folk remedies since ancient times. J. Herbs Spices Med. Plants 2014, 20, 103–114. [Google Scholar] [CrossRef]
- Demetzos, C.; Harvala, C.; Philianos, S.M.; Skaltsounis, A.L. A new labdane-type diterpene and other compounds from the leaves of Cistus incanus ssp. creticus. J. Nat. Prod. 1990, 53, 1365–1368. [Google Scholar] [CrossRef]
- Weyerstahl, P.; Marschall, H.; Weirauch, M.; Thefeld, K.; Surburg, H. Constituents of commercial Labdanum oil. Flavour Fragr. J. 1998, 13, 295–318. [Google Scholar] [CrossRef]
- Fahn, A.; Cutler, D.F. Xerophytes. In Encyclopedia of Plant Anatomy; Borntraeger: Berlin, Germany, 1992; Volume XIII. [Google Scholar]
- Catoni, R.L.; Gratani, L.; Varone, L. Physiological, morphological and anatomical trait variations between winter and summer leaves of Cistus species. Flora 2012, 207, 442–449. [Google Scholar] [CrossRef]
- Sharma, S.; Parihar, L. Effect of mobile phone radiation on nodule formation in the leguminous plants. Curr. World Environ. J. 2014, 9, 145–155. [Google Scholar] [CrossRef]
- Roux, D.; Vian, A.; Girard, S.; Bonnet, P.; Paladian, F.; Davies, E.; Ledoigt, G. Electromagnetic fields (900 MHz) evoke consistent molecular responses in tomato plants. Physiol. Plant. 2006, 128, 283–288. [Google Scholar] [CrossRef]
- Pietruszewski, S.; Muszyñski, S.; Dziwulska, A. Electromagnetic fields and electromagnetic radiation as non-invasive external stimulants for seeds (selected methods and responses). Int. Agrophysics 2007, 21, 95–100. [Google Scholar]
- Sheridan, S.; Kalkstein, L.; Kalkstein, A.; Greene, S. Heat-related Mortality and Heat Watch-warning Systems in the United States: Recent Developments. In Proceedings of the ISEE 22nd Annual Conference, Seoul, Korea, 28 August–1 September 2010. [Google Scholar]
- Vian, A.; Roux, D.; Girard, S.; Bonnet, P.; Paladian, F.; Davies, E.; Ledoigt, G. Microwave irradiation affects gene expression in plants. Plant Signal. Behav. 2006, 1, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Haggerty, K. Adverse influence of radio frequency background on trembling aspen seedlings: Preliminary observations. Int. J. For. Res. 2010, 2010, 836278. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, H.P.; Batish, D.R.; Kaur, S.; Kumar-Kohli, R. EMF radiations (1800 MHz)-inhibited early seedling growth of maize (Zea mays) involves alterations in starch and sucrose metabolism. Protoplasma 2015, 253, 1043–1099. [Google Scholar] [CrossRef]
- Hollósy, F. Effects of ultraviolet radiation on plant cells. Micron 2002, 33, 179–197. [Google Scholar] [CrossRef]
- Waldmann-Selsam, C.; Balmori-de la Puente, A.; Breunig, H.; Balmori, A. Radiofrequency radiation injures trees around mobile phone base stations. Sci. Total Environ. 2016, 572, 554–569. [Google Scholar] [CrossRef]
- Stefi, A.L.; Margaritis, L.H.; Christodoulakis, N.S. The effect of the non-ionizing radiation on exposed, laboratory cultivated maize (Zea mays L.) plants. Flora 2017, 233, 22–30. [Google Scholar] [CrossRef]
- Vian, A.; Davies, E.; Gendraud, M.; Bonnet, P. Plant responses to high frequency electromagnetic fields. Biomed. Res. Int. 2016, 2016, 1830262. [Google Scholar] [CrossRef]
- Stefi, A.L.; Margaritis, L.H.; Christodoulakis, N.S. The effect of the non-ionizing radiation on exposed, laboratory cultivated upland cotton (Gossypium hirsutum L.) plants. Flora 2017, 226, 55–64. [Google Scholar] [CrossRef]
- Stefi, A.L.; Margaritis, L.H.; Christodoulakis, N.S. The aftermath of long-term exposure to non-ionizing radiation on laboratory cultivated pine plants (Pinus halepensis M.). Flora 2017, 234, 173–186. [Google Scholar] [CrossRef]
- Tocher, R.D.; Tocher, C.S. Dopa decarboxylase in Cytisus scoparius. Phytochemistry 1972, 11, 1661–1667. [Google Scholar] [CrossRef]
- Iriti, M. Plant neurobiology, a fascinating perspective in the field of research on plant secondary metabolites. Int. J. Mol. Sci. 2013, 14, 10819–10821. [Google Scholar] [CrossRef] [PubMed]
- Stefi, A.L.; Vassilacopoulou, D.; Christodoulakis, N.S. Environmentally stressed summer leaves of the seasonally dimorphic Phlomis fruticosa and the relief through the L-Dopa decarboxylase (DDC). Flora 2019, 251, 11–19. [Google Scholar] [CrossRef]
- Jaffe, M.J. Evidence for the regulation of phytochrome-mediated processes in bean roots by the neurohumor, acetylcholine. Plant Physiol. 1970, 46, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Barlow, R.B.; Dixon, R.O.D. Choline acetyltransferase in the nettle Urtica dioica L. Biochem. J. 1973, 132, 15–18. [Google Scholar] [CrossRef] [Green Version]
- Odjakova, M.; Hadjiivanova, C. Animal neurotransmitter substances in plants. Bulg. J. Plant Physiol. 1997, 23, 94–102. [Google Scholar]
- Soares, A.R.; Marchiosi, R.; Siqueira-Soares, R.d.C.; Lima, R.B.d.; Santos, W.D.d.; Ferrarese-Filho, O. The role of L-DOPA in plants. Plant Signal. Behav. 2014, 9, e28275. [Google Scholar] [CrossRef]
- Stefi, A.L.; Vassilacopoulou, D.; Routsi, E.; Stathopoulos, P.; Argyropoulou, A.; Skaltsounis, A.-L.; Christodoulakis, N.S. The Combined Environmental Stress on the Leaves of Olea europaea L. and the Relief Mechanism Through Biosynthesis of Certain Secondary Metabolites. J. Plant Growth Regul. 2020, 40, 1044–1059. [Google Scholar] [CrossRef]
- Kulma, A.; Szopa, J. Catecholamines are active compounds in plants. Plant Sci. 2007, 172, 433–440. [Google Scholar] [CrossRef]
- Liu, Q.; Gao, T.; Liu, Q.; Liu, Y.; Zhao, Y.; Liu, Y.; Li, W.; Ding, K.; Ma, F.; Li, C. Functions of dopamine in plants: A review. Plant Signal. Behav. 2020, 15, e1827782. [Google Scholar] [CrossRef]
- Stefi, A.L.; Margaritis, L.H.; Christodoulakis, N.S. The effect of the non-ionizing radiation on cultivated plants of Arabidopsis thaliana (Col.). Flora 2016, 223, 114–120. [Google Scholar] [CrossRef]
- Stefi, A.L.; Vassilacopoulou, D.; Margaritis, L.H.; Christodoulakis, N.S. Oxidative stress and an animal neurotransmitter synthesizing enzyme in the leaves of wild growing myrtle after exposure to GSM radiation. Flora 2018, 243, 67–76. [Google Scholar] [CrossRef]
- Aničić, N.; Patelou, E.; Papanikolaou, A.; Kanioura, A.; Valdesturli, C.; Arapitsas, P.; Skorić, M.; Dragićević, M.; Gašić, U.; Koukounaras, A.; et al. Comparative Metabolite and Gene Expression Analyses in Combination with Gene Characterization Revealed the Patterns of Flavonoid Accumulation During Cistus criticus subsp. creticus Fruit Development. Front. Plant Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Atsalakis, E.; Chinou, I.; Makropoulou, M.; Karabournioti, S.; Graikou, K. Evaluation of Phenolic Compounds in Cistus creticus Bee Pollen from Greece. Antioxidant and Antimicrobial Properties. Nat. Prod. Commun. 2017. [Google Scholar] [CrossRef]
- Stefi, A.L.; Mitsigiorgi, K.; Vassilacopoulou, D.; Christodoulakis, N.S. Response of young Nerium oleander plants to long-term non-ionizing radiation. Planta 2020, 251, 108–125. [Google Scholar] [CrossRef] [PubMed]
- Doussi, M.A.; Thanos, C.A. Post-fire regeneration of hard seeded plants: Ecophysiology of seed germination. In Proceedings of the 2nd International Conference Forest Fire Research, Coimbra, Portugal, 21–24 November 1994. [Google Scholar]
- Margaritis, L.; Manta, A.K.; Kokkaliaris, K.D.; Schiza, D.; Alimisis, K.; Barkas, G.; Georgiou, E.; Giannakopoulou, O.; Kollia, I.; Kontogianni, G.; et al. Drosophila oogenesis as a bio-marker responding to EMF sources. Electromagn. Biol. Med. 2014, 33, 165–189. [Google Scholar] [CrossRef] [PubMed]
- ICNIRP. Guidelines for limiting exposure to time-varying electric, magnetic and electromagnetic fields (up to 300 GHz). Health Phys. 1998, 74, 494–522. [Google Scholar]
- Sabatini, D.; Bensch, K.; Barrnett, B. Cytochemistry and electron microscopy. The preservation of cellular ultrastructure and enzymatic activity by aldehyde fixation. J. Cell Biol. 1963, 17, 208–212. [Google Scholar]
- Christodoulakis, N.S.; Tsiarta, M.; Fasseas, C. Leaf structure and histochemical investigation in Papaver rhoeas L. (corn poppy). J. Herbs Spices Med. Plants 2013, 19, 119–131. [Google Scholar] [CrossRef]
- Gechev, T.; Mehterov, N.; Denev, I.; Hille, J. A Simple and Powerful Approach for Isolation of Arabidopsis Mutants with Increased Tolerance to H2O2 Induced Cell Death. Methods Enzymol. 2013, 527, 203–220. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Ann. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed]
- Batteiger, B.; Newhall, W.J.; Jones, R.B. The use of Tween 20 as a blocking agent in the immunological detection of proteins transferred to nitrocellulose membranes. J. Immunol. Methods 1982, 55, 297–307. [Google Scholar] [CrossRef]
- Sunkar, R. Review: MicroRNAs with macro-effects on plant stress responses. Semin. Cell Dev. Biol. 2010, 21, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Athanasakis, G.; Aligiannis, N.; Gonou-Zagou, Z.; Skaltsounis, A.-L.; Fokialakis, N. Antioxidant Properties of the Wild Edible Mushroom Lactarius salmonicolor. Full Communications. J. Med. Food 2013, 16, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Kukula-Koch, W.; Aligiannis, N.; Halabalaki, M.; Skaltsounis, A.-L.; Glowniak, K.; Kalpoutzakis, E. Influence of extraction procedures on phenolic content and antioxidant activity of Cretan barberry herb. Food Chem. 2013, 138, 406–413. [Google Scholar] [CrossRef]
- Kouka, P.; Priftis, A.; Stagos, D.; Angelis, A.; Stathopoulos, P.; Xinos, N.; Skaltsounis, A.-L.; Mamoulakis, C.; Tsatsakis, A.M.; Spandidos, D.; et al. Assessment of the antioxidant activity of an olive oil total polyphenolic fraction and hydroxytyrosol from a Greek Olea europaea variety in endothelial cells and myoblasts. Int. J. Mol. Med. 2017, 40, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetry of total phenolics with phosphomolybdic-phosphotungtic acid reagent. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Salluca, T.G.; Penarrieta, J.M.; Alvarado, J.A.; Bergenstahl, B. Determination of total phenolic compounds content and the antioxidant capacity of Andean tubers and roots (isaño, oca, ulluco and arracacha). Rev. Bol. Quim. 2008, 25, 59–62. [Google Scholar]
- Smirnoff, N. Antioxidant Systems and Plant Response to the Environment. In Environment and Plant Metabolism: Flexibility and Acclimation; Smirnoff, V., Ed.; BIOS Scientific Publishers: Oxford, UK, 1995; pp. 217–243. [Google Scholar]
- Manivannan, P.; Jaleel, C.A.; Sankar, B.; Kishorekumar, A.; Somasundaram, R.; Lakshmanan, G.M.A.; Panneerselvam, R.N. Growth, biochemical modifications and proline metabolism in Helianthus annuus L. as induced by drought stress. Colloids Surf. B Biointerfaces 2007, 59, 141–149. [Google Scholar] [CrossRef]
- Kratsch, H.A.; Wise, R.R. The ultrastructure of chilling stress. Plant Cell Environ. 2000, 23, 337–350. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Aminaka, R.; Yoshioka, M.; Khatoon, M.; Komayama, K.; Takenaka, D.; Yamashita, A.; Nijo, N.; Inagawa, K.; Morita, N.; et al. Quality control of photosystem II: Impact of light and heat stresses. Photosynth. Res. 2008, 98, 589–608. [Google Scholar] [CrossRef]
- Stephanou, M.; Manetas, Y. The effects of seasons, exposure, enhanced UV-B radiation, and water stress on leaf epicuticular and internal UV-B absorbing capacity of Cistus creticus: A Mediterranean field study. J. Exp. Bot. 1997, 48, 1977–1985. [Google Scholar] [CrossRef]
- Nikolakaki, A.; Christodoulakis, N.S. Secreting formations and cytochemical investigation of the leaf of Phlomis fruticosa, a seasonally dimorphic subshrub. Secreting activity of the leaf-originating calluses. Flora 2007, 202, 429–436. [Google Scholar] [CrossRef]
- Weidner, S.; Karolac, M.; Karamać, M.; Kosińska, A.; Amarowitcz, R. Phenolic compounds and properties of antioxidants in grapevine roots (Vitis vinifera L.) under drought stress followed by recovery. Acta Soc. Bot. Pol. 2009, 78, 97–103. [Google Scholar] [CrossRef]
- Kanazawa, K.; Sakakibara, H. High content of dopamine, a strong antioxidant, in Cavendish banana. J. Agric. Food Chem. 2000, 48, 844–848. [Google Scholar] [CrossRef]
- Pedroso, A.N.V.; Bussotti, F.; Tani, C.; Papini, A.; Domingos, M. Pollution emissions from a petrochemical complex and other environmental stressors induce structural and ultrastructural damage in leaves of a biosensor tree species from the Atlantic Rain Forest. Ecol. Indic. 2016, 67, 215–226. [Google Scholar] [CrossRef]
- Kaur, A.; Thurkral, A.K. Effect of animal hormones on the growth, protein and sugar contents of Vigna unguiculate L. seedlings. Indian J. Plant Physiol. 1990, 33, 259–261. [Google Scholar]
- Christodoulakis, N.S.; Fasseas, C. Air pollution effects on the leaf structure of Laurus nobilis, an injury resistant species. Bull. Environ. Contam. Toxicol. 1990, 44, 276–281. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, Y.; Jia, B.; Zhou, G. Elevated-CO2 Response of Stomata and Its Dependence on Environmental Factors. Front. Plant Sci. 2016, 7, 657. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Haworth, M.; Elliott-Kingston, C.; McElwain, J.C. Co-ordination of physiological and morphological responses of stomata to elevated [CO2] in vascular plants. Oecologia 2013, 171, 71–82. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, F.M.; Godoy, A.G.; Menezes-Silva, P.E.; Martins, S.C.V.; Sanglard, L.M.V.P.; Morais, L.E.; Torre-Neto, A.; Ghini, R. Sustained enhancement of photosynthesis in coffee trees grown under free-air CO2 enrichment conditions: Disentangling the contributions of stomatal, mesophyll, and biochemical limitations. J. Exp. Bot. 2016, 67, 341–352. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefi, A.L.; Kalouda, G.; Skouroliakou, A.S.; Vassilacopoulou, D.; Christodoulakis, N.S. The Counteraction of Cultivated Cistus creticus L. (Rock Rose) Plants to the Strain Imposed by a Long-Term Exposure to Non-Ionizing Radiation and the Role of DDC. Biophysica 2022, 2, 248-265. https://doi.org/10.3390/biophysica2030024
Stefi AL, Kalouda G, Skouroliakou AS, Vassilacopoulou D, Christodoulakis NS. The Counteraction of Cultivated Cistus creticus L. (Rock Rose) Plants to the Strain Imposed by a Long-Term Exposure to Non-Ionizing Radiation and the Role of DDC. Biophysica. 2022; 2(3):248-265. https://doi.org/10.3390/biophysica2030024
Chicago/Turabian StyleStefi, Aikaterina L., Georgia Kalouda, Aikaterini S. Skouroliakou, Dido Vassilacopoulou, and Nikolaos S. Christodoulakis. 2022. "The Counteraction of Cultivated Cistus creticus L. (Rock Rose) Plants to the Strain Imposed by a Long-Term Exposure to Non-Ionizing Radiation and the Role of DDC" Biophysica 2, no. 3: 248-265. https://doi.org/10.3390/biophysica2030024
APA StyleStefi, A. L., Kalouda, G., Skouroliakou, A. S., Vassilacopoulou, D., & Christodoulakis, N. S. (2022). The Counteraction of Cultivated Cistus creticus L. (Rock Rose) Plants to the Strain Imposed by a Long-Term Exposure to Non-Ionizing Radiation and the Role of DDC. Biophysica, 2(3), 248-265. https://doi.org/10.3390/biophysica2030024