
Developmental Pattern Formation: Spanish Contributions from a Biophysical Perspective

{kind=link}
{kind=link}
Abstract
:1. Introduction: Twenty Years of a Polemic Statement
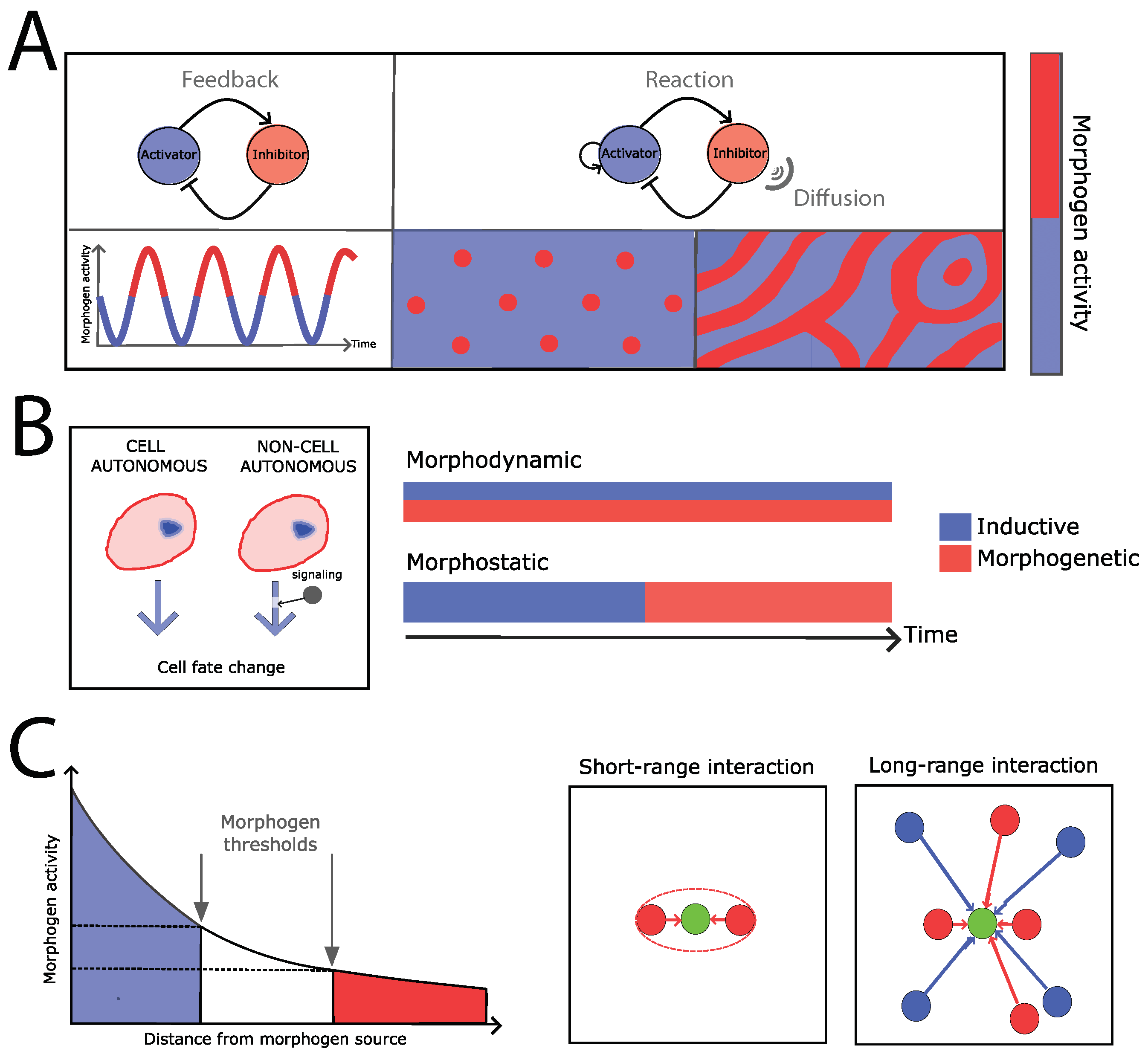
2. Developmental Pattern Formation: From Physics to Biology, from Models to Mechanisms
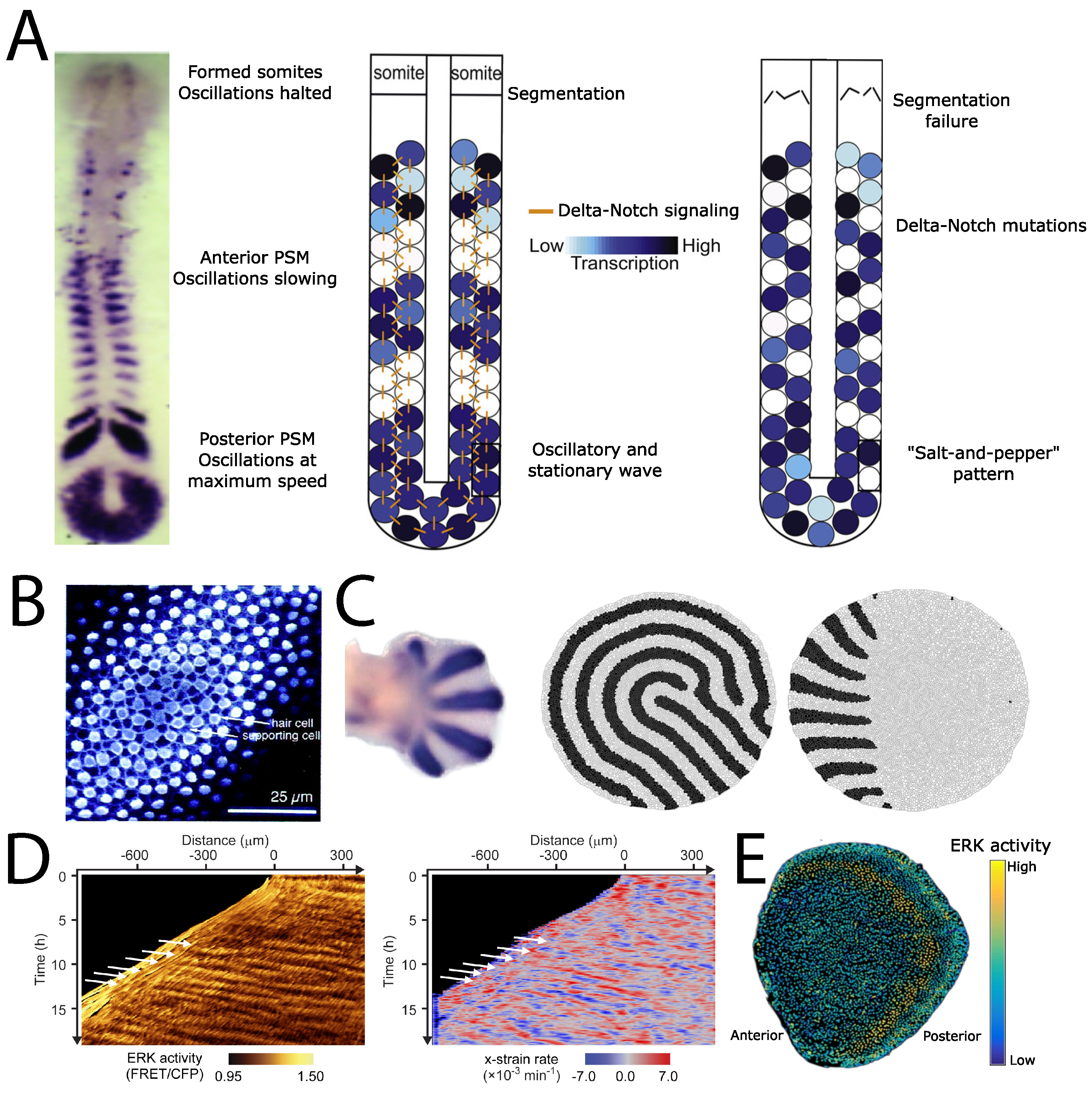
3. Examples of Developmental Patterning

4. Discussion
Funding
Acknowledgments
Conflicts of Interest
References
- Gilbert, S.F. Developmental Biology, 7th ed.; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Oates, A.C.; Gorfinkiel, N.; González-Gaitán, M.; Heisenberg, C.P. Quantitative approaches in developmental biology. Nat. Rev. Genet. 2009, 10, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Turing, A. The Chemical Basis of Morphogenesis. Philos. Trans. R. Soc. Lond. Ser. B 1990, 237, 37–72. [Google Scholar] [CrossRef]
- Wolpert, L. Positional information and pattern formation in development. Dev. Genet. 1994, 15, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.J.; Meinhardt, H. Biological pattern formation: From basic mechanisms to complex structures. Rev. Mod. Phys. 1994, 66, 1481–1507. [Google Scholar] [CrossRef]
- Murray, J.D. Mathematical Biology. 1: An Introduction, Softcover Reprint of the Hardcover 3rd edition 2002, corrected second printing ed.; Number 17 in Interdisciplinary applied mathematics; Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Forgacs, G.; Newman, S.A. Biological Physics of the Developing Embryo, 1st ed.; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar] [CrossRef]
- García-Ojalvo, J.; Sancho, J.M. Noise in Spatially Extended Systems; Institute for nonlinear science; Springer: New York, NY, USA, 1999. [Google Scholar]
- Sagués, F.; Sancho, J.; García-Ojalvo, J. Spatiotemporal order out of noise. Rev. Mod. Phys. 2007, 79, 829–882. [Google Scholar] [CrossRef]
- Lewis, J. From signals to patterns: Space, time, and mathematics in developmental biology. Science 2008, 322, 399–403. [Google Scholar] [CrossRef]
- Kicheva, A.; Cohen, M.; Briscoe, J. Developmental pattern formation: Insights from physics and biology. Science 2012, 338, 210–212. [Google Scholar] [CrossRef]
- Cross, M.; Hohenberg, P. Pattern formation outside of equilibrium. Rev. Mod. Phys. 1993, 65, 851–1112. [Google Scholar] [CrossRef]
- Salazar-Ciudad, I.; Jernvall, J.; Newman, S.A. Mechanisms of pattern formation in development and evolution. Development 2003, 130, 2027–2037. [Google Scholar] [CrossRef]
- Salazar-Ciudad, I.; Jernvall, J. How different types of pattern formation mechanisms affect the evolution of form and development. Evol. Dev. 2004, 6, 6–16. [Google Scholar] [CrossRef]
- Buceta, J.; Lindenberg, K. Patterns in reaction-diffusion systems generated by global alternation of dynamics. Phys. Stat. Mech. Its Appl. 2003, 325, 230–242. [Google Scholar] [CrossRef]
- Morelli, L.G.; Uriu, K.; Ares, S.; Oates, A.C. Computational approaches to developmental patterning. Science 2012, 336, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Wolpert, L. Positional information and the spatial pattern of cellular differentiation. J. Theor. Biol. 1969, 25, 1–47. [Google Scholar] [CrossRef]
- Diego, X.; Marcon, L.; Müller, P.; Sharpe, J. Key Features of Turing Systems are Determined Purely by Network Topology. Phys. Rev. X 2018, 8, 021071. [Google Scholar] [CrossRef]
- Kondo, S.; Miura, T. Reaction-diffusion model as a framework for understanding biological pattern formation. Science 2010, 329, 1616–1620. [Google Scholar] [CrossRef]
- Landge, A.N.; Jordan, B.M.; Diego, X.; Müller, P. Pattern formation mechanisms of self-organizing reaction-diffusion systems. Dev. Biol. 2020, 460, 2–11. [Google Scholar] [CrossRef]
- Collier, J.R.; Monk, N.A.; Maini, P.K.; Lewis, J.H. Pattern formation by lateral inhibition with feedback: A mathematical model of delta-notch intercellular signalling. J. Theor. Biol. 1996, 183, 429–446. [Google Scholar] [CrossRef] [PubMed]
- Talia, S.D.; Vergassola, M. Waves in Embryonic Development. Annu. Rev. Biophys. 2022, 51, 327–353. [Google Scholar] [CrossRef]
- Oates, A.C.; Morelli, L.G.; Ares, S. Patterning embryos with oscillations: Structure, function and dynamics of the vertebrate segmentation clock. Development 2012, 139, 625–639. [Google Scholar] [CrossRef]
- Kageyama, R. 25 years of the segmentation clock gene. Nature 2022, 611, 671–673. [Google Scholar] [CrossRef]
- Webb, A.B.; Lengyel, I.M.; Jörg, D.J.; Valentin, G.; Jülicher, F.; Morelli, L.G.; Oates, A.C. Persistence, period and precision of autonomous cellular oscillators from the zebrafish segmentation clock. eLife 2016, 5, e08438. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.; Hanisch, A.; Holder, M. Notch signaling, the segmentation clock, and the patterning of vertebrate somites. J. Biol. 2009, 8, 44. [Google Scholar] [CrossRef] [PubMed]
- Buceta, J. Multidisciplinary approaches towards compartmentalization in development: Dorsoventral boundary formation of the Drosophila wing disc as a case of study. Contrib. Sci. 2013, 9, 57–66. [Google Scholar] [CrossRef]
- Briscoe, J.; Small, S. Morphogen rules: Design principles of gradient-mediated embryo patterning. Development 2015, 142, 3996–4009. [Google Scholar] [CrossRef]
- Diaz-Cuadros, M.; Pourquié, O.; El-Sherif, E. Patterning with clocks and genetic cascades: Segmentation and regionalization of vertebrate versus insect body plans. PLoS Genet. 2021, 17, e1009812. [Google Scholar] [CrossRef]
- Jaeger, J.; Surkova, S.; Blagov, M.; Janssens, H.; Kosman, D.; Kozlov, K.N.; Manu; Myasnikova, E.; Vanario-Alonso, C.E.; Samsonova, M.; et al. Dynamic control of positional information in the early Drosophila embryo. Nature 2004, 430, 368–371. [Google Scholar] [CrossRef]
- Crombach, A.; Wotton, K.R.; Cicin-Sain, D.; Ashyraliyev, M.; Jaeger, J. Efficient reverse-engineering of a developmental gene regulatory network. PLoS Comput. Biol. 2012, 8, e1002589. [Google Scholar] [CrossRef]
- Jaeger, J.; Verd, B. Dynamic positional information: Patterning mechanism versus precision in gradient-driven systems. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2020; Volume 137, pp. 219–246. [Google Scholar] [CrossRef]
- Verd, B.; Clark, E.; Wotton, K.R.; Janssens, H.; Jiménez-Guri, E.; Crombach, A.; Jaeger, J. A damped oscillator imposes temporal order on posterior gap gene expression in Drosophila. PLoS Biol. 2018, 16, e2003174. [Google Scholar] [CrossRef]
- Verd, B.; Monk, N.A.; Jaeger, J. Modularity, criticality, and evolvability of a developmental gene regulatory network. eLife 2019, 8, e42832. [Google Scholar] [CrossRef]
- Balaskas, N.; Ribeiro, A.; Panovska, J.; Dessaud, E.; Sasai, N.; Page, K.M.; Briscoe, J.; Ribes, V. Gene regulatory logic for reading the Sonic Hedgehog signaling gradient in the vertebrate neural tube. Cell 2012, 148, 273–284. [Google Scholar] [CrossRef]
- Kicheva, A.; Bollenbach, T.; Ribeiro, A.; Valle, H.P.; Lovell-Badge, R.; Episkopou, V.; Briscoe, J. Coordination of progenitor specification and growth in mouse and chick spinal cord. Science 2014, 345, 1254927. [Google Scholar] [CrossRef] [PubMed]
- Saade, M.; Gutiérrez-Vallejo, I.; Le Dréau, G.; Rabadán, M.A.; Miguez, D.G.; Buceta, J.; Martí, E. Sonic Hedgehog Signaling Switches the Mode of Division in the Developing Nervous System. Cell Rep. 2013, 4, 492. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Hidalgo, D.; Domínguez-Cejudo, M.A.; Amore, G.; Brockmann, A.; Lemos, M.C.; Córdoba, A.; Casares, F. A Hh-driven gene network controls specification, pattern and size of the Drosophila simple eyes. Development 2013, 140, 82–92. [Google Scholar] [CrossRef] [PubMed]
- García-Morales, D.; Navarro, T.; Iannini, A.; Míguez, D.G.; Casares, F. Dynamic Hh signaling can generate temporal information during tissue patterning. Development 2019, 146, dev.176933. [Google Scholar] [CrossRef]
- Míguez, D.G.; García-Morales, D.; Casares, F. Control of size, fate and time by the Hh morphogen in the eyes of flies. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2020; Volume 137, pp. 307–332. [Google Scholar] [CrossRef]
- Giudicelli, F.; Özbudak, E.M.; Wright, G.J.; Lewis, J. Setting the Tempo in Development: An Investigation of the Zebrafish Somite Clock Mechanism. PLoS Biol. 2007, 5, e150. [Google Scholar] [CrossRef]
- Keskin, S.; Devakanmalai, G.S.; Kwon, S.B.; Vu, H.T.; Hong, Q.; Lee, Y.Y.; Soltani, M.; Singh, A.; Ay, A.; Özbudak, E.M. Noise in the Vertebrate Segmentation Clock Is Boosted by Time Delays but Tamed by Notch Signaling. Cell Rep. 2018, 23, 2175–2185.e4. [Google Scholar] [CrossRef]
- Eddison, M.; Le Roux, I.; Lewis, J. Notch signaling in the development of the inner ear: Lessons from Drosophila. Proc. Natl. Acad. Sci. USA 2000, 97, 11692–11699. [Google Scholar] [CrossRef]
- Sheth, R.; Marcon, L.; Bastida, M.F.; Junco, M.; Quintana, L.; Dahn, R.; Kmita, M.; Sharpe, J.; Ros, M.A. Hox Genes Regulate Digit Patterning by Controlling the Wavelength of a Turing-Type Mechanism. Science 2012, 338, 1476–1480. [Google Scholar] [CrossRef]
- Anbari, S.; Buceta, J. Self-sustained planar intercalations due to mechanosignaling feedbacks lead to robust axis extension during morphogenesis. Sci. Rep. 2020, 10, 10973. [Google Scholar] [CrossRef]
- Hino, N.; Rossetti, L.; Marín-Llauradó, A.; Aoki, K.; Trepat, X.; Matsuda, M.; Hirashima, T. ERK-Mediated Mechanochemical Waves Direct Collective Cell Polarization. Dev. Cell 2020, 53, 646–660. [Google Scholar] [CrossRef]
- De Simone, A.; Evanitsky, M.N.; Hayden, L.; Cox, B.D.; Wang, J.; Tornini, V.A.; Ou, J.; Chao, A.; Poss, K.D.; Di Talia, S. Control of osteoblast regeneration by a train of Erk activity waves. Nature 2021, 590, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Dahmann, C.; Basler, K. Compartment boundaries: At the edge of development. Trends Genet. TIG 1999, 15, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Morata, G.; Lawrence, P.A. Anterior and posterior compartments in the head of Drosophila. Nature 1978, 274, 473–474. [Google Scholar] [CrossRef]
- Lawrence, P.A. Developmental biology. Straight and wiggly affinities. Nature 1997, 389, 546–547. [Google Scholar] [CrossRef]
- Landsberg, K.P.; Farhadifar, R.; Ranft, J.; Umetsu, D.; Widmann, T.J.; Bittig, T.; Said, A.; Jülicher, F.; Dahmann, C. Increased Cell Bond Tension Governs Cell Sorting at the Drosophila Anteroposterior Compartment Boundary. Curr. Biol. 2009, 19, 1950–1955. [Google Scholar] [CrossRef] [PubMed]
- Umetsu, D.; Dahmann, C. Compartment boundaries: Sorting cells with tension. Fly 2010, 4, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Canela-Xandri, O.; Sagués, F.; Casademunt, J.; Buceta, J. Dynamics and Mechanical Stability of the Developing Dorsoventral Organizer of the Wing Imaginal Disc. PLoS Comput. Biol. 2011, 7, e1002153. [Google Scholar] [CrossRef]
- Buceta, J.; Herranz, H.; Canela-Xandri, O.; Reigada, R.; Sagués, F.; Milán, M. Robustness and stability of the gene regulatory network involved in DV boundary formation in the Drosophila wing. PLoS ONE 2007, 2, e602. [Google Scholar] [CrossRef]
- Canela-Xandri, O.; Sagués, F.; Reigada, R.; Buceta, J. A spatial toggle switch drives boundary formation in development. Biophys. J. 2008, 95, 5111–5120. [Google Scholar] [CrossRef]
- Becam, I.; Milán, M. A permissive role of Notch in maintaining the DV affinity boundary of the Drosophila wing. Dev. Biol. 2008, 322, 190–198. [Google Scholar] [CrossRef]
- Bocci, F.; Onuchic, J.N.; Jolly, M.K. Understanding the Principles of Pattern Formation Driven by Notch Signaling by Integrating Experiments and Theoretical Models. Front. Physiol. 2020, 11, 929. [Google Scholar] [CrossRef] [PubMed]
- Sancho, J.M.; Ibañes, M. Landau theory for cellular patterns driven by lateral inhibition interaction. Phys. Rev. E 2020, 102, 032404. [Google Scholar] [CrossRef]
- Petrovic, J.; Formosa-Jordan, P.; Luna-Escalante, J.C.; Abelló, G.; Ibañes, M.; Neves, J.; Giraldez, F. Ligand-dependent Notch signaling strength orchestrates lateral induction and lateral inhibition in the developing inner ear. Development 2014, 141, 2313–2324. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.C.; Lyons, E.L.; Herman, T.G. cis-Inhibition of Notch by Endogenous Delta Biases the Outcome of Lateral Inhibition. Curr. Biol. 2009, 19, 1378–1383. [Google Scholar] [CrossRef] [PubMed]
- Palau-Ortin, D.; Formosa-Jordan, P.; Sancho, J.M.; Ibañes, M. Pattern selection by dynamical biochemical signals. Biophys. J. 2015, 108, 1555–1565. [Google Scholar] [CrossRef] [PubMed]
- Formosa-Jordan, P.; Ibañes, M. Diffusible ligand and lateral inhibition dynamics for pattern formation. J. Stat. Mech. Theory Exp. 2009, 2009, P03019. [Google Scholar] [CrossRef]
- Formosa-Jordan, P.; Ibañes, M. Competition in notch signaling with cis enriches cell fate decisions. PLoS ONE 2014, 9, e95744. [Google Scholar] [CrossRef]
- Fàbregas, N.; Formosa-Jordan, P.; Confraria, A.; Siligato, R.; Alonso, J.M.; Swarup, R.; Bennett, M.J.; Mähönen, A.P.; Caño-Delgado, A.I.; Ibañes, M. Auxin Influx Carriers Control Vascular Patterning and Xylem Differentiation in Arabidopsis thaliana. PLoS Genet. 2015, 11, e1005183. [Google Scholar] [CrossRef]
- Formosa-Jordan, P.; Ibañes, M.; Ares, S.; Frade, J.M. Regulation of neuronal differentiation at the neurogenic wavefront. Development 2012, 139, 2321–2329. [Google Scholar] [CrossRef]
- Formosa-Jordan, P.; Ibañes, M.; Ares, S.; Frade, J.M. Lateral inhibition and neurogenesis: Novel aspects in motion. Int. J. Dev. Biol. 2013, 57, 341–350. [Google Scholar] [CrossRef]
- Ouyang, Q.; Swinney, H.L. Transition from a uniform state to hexagonal and striped Turing patterns. Nature 1991, 352, 610–612. [Google Scholar] [CrossRef]
- Rüdiger, S.; Míguez, D.G.; Muñuzuri, A.P.; Sagués, F.; Casademunt, J. Dynamics of Turing Patterns under Spatiotemporal Forcing. Phys. Rev. Lett. 2003, 90, 128301. [Google Scholar] [CrossRef] [PubMed]
- Míguez, D.G.; Nicola, E.M.; Muñuzuri, A.P.; Casademunt, J.; Sagués, F.; Kramer, L. Traveling-Stripe Forcing Generates Hexagonal Patterns. Phys. Rev. Lett. 2004, 93, 048303. [Google Scholar] [CrossRef] [PubMed]
- Kærn, M.; Míguez, D.G.; Muñuzuri, A.P.; Menzinger, M. Control of chemical pattern formation by a clock-and-wavefront type mechanism. Biophys. Chem. 2004, 110, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Konow, C.; Dolnik, M.; Epstein, I.R. Insights from chemical systems into Turing-type morphogenesis. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2021, 379, 20200269. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Yoshimoto, E.; Kondo, S. Pattern regulation in the stripe of zebrafish suggests an underlying dynamic and autonomous mechanism. Proc. Natl. Acad. Sci. USA 2007, 104, 4790–4793. [Google Scholar] [CrossRef]
- Nakamasu, A.; Takahashi, G.; Kanbe, A.; Kondo, S. Interactions between zebrafish pigment cells responsible for the generation of Turing patterns. Proc. Natl. Acad. Sci. USA 2009, 106, 8429–8434. [Google Scholar] [CrossRef]
- Kondo, S. Turing Pattern Formation without Diffusion. In How the World Computes; Springer: Berlin/Heidelberg, Germany, 2012; pp. 416–421. [Google Scholar] [CrossRef]
- Economou, A.D.; Ohazama, A.; Porntaveetus, T.; Sharpe, P.T.; Kondo, S.; Basson, M.A.; Gritli-Linde, A.; Cobourne, M.T.; Green, J.B.A. Periodic stripe formation by a Turing mechanism operating at growth zones in the mammalian palate. Nat. Genet. 2012, 44, 348–351. [Google Scholar] [CrossRef]
- Raspopovic, J.; Marcon, L.; Russo, L.; Sharpe, J. Modeling digits. Digit patterning is controlled by a Bmp-Sox9-Wnt Turing network modulated by morphogen gradients. Science 2014, 345, 566–570. [Google Scholar] [CrossRef]
- Cooper, K.L. Self-organization in the limb: A Turing mechanism for digit development. Curr. Opin. Genet. Dev. 2015, 32, 92–97. [Google Scholar] [CrossRef]
- Marcon, L.; Diego, X.; Sharpe, J.; Müller, P. High-throughput mathematical analysis identifies Turing networks for patterning with equally diffusing signals. eLife 2016, 5, e14022. [Google Scholar] [CrossRef] [PubMed]
- Buceta, J. Finite cell-size effects on protein variability in Turing patterned tissues. J. R. Soc. Interface 2017, 14, 20170316. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Buceta, J. A non-linear analysis of Turing pattern formation. PLoS ONE 2019, 14, e0220994. [Google Scholar] [CrossRef]
- Jacobs, C.R.; Huang, H.; Kwon, R.Y. Introduction to Cell Mechanics and Mechanobiology; OCLC: 1019679867; Garland Science: New York, NY, USA, 2013. [Google Scholar]
- Tambe, D.; Hardin, C.; Angelini, T.E.; Rajendran, K.; Park, C.Y.; Serra-Picamal, X.; Zhou, E.H.; Zaman, M.H.; Butler, J.P.; Weitz, D.A.; et al. Collective cell guidance by cooperative intercellular forces. Nat. Mat. 2011, 10, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Gonzalez, R.; Zallen, J.A. Epithelial organization: May the force be with you. Curr. Biol. 2008, 18, R163–R165. [Google Scholar] [CrossRef]
- Gómez-Gálvez, P.; Anbari, S.; Escudero, L.M.; Buceta, J. Mechanics and self-organization in tissue development. Semin. Cell Dev. Biol. 2021, 120, 147. [Google Scholar] [CrossRef]
- Serra-Picamal, X.; Conte, V.; Vincent, R.; Anon, E.; Tambe, D.T.; Bazellieres, E.; Butler, J.P.; Fredberg, J.J.; Trepat, X. Mechanical waves during tissue expansion. Nat. Phys. 2012, 8, 628–634. [Google Scholar] [CrossRef]
- Pérez-González, C.; Alert, R.; Blanch-Mercader, C.; Gómez-González, M.; Kolodziej, T.; Bazellieres, E.; Casademunt, J.; Trepat, X. Active wetting of epithelial tissues. Nat. Phys. 2019, 15, 79–88. [Google Scholar] [CrossRef]
- Prat-Rojo, C.; Pouille, P.A.; Buceta, J.; Martin-Blanco, E. Mechanical coordination is sufficient to promote tissue replacement during metamorphosis in Drosophila. EMBO J. 2020, 39, e103594. [Google Scholar] [CrossRef]
- Alert, R.; Trepat, X. Physical Models of Collective Cell Migration. Annu. Rev. Condens. Matter Phys. 2020, 11, 77–101. [Google Scholar] [CrossRef]
- Latorre, E.; Kale, S.; Casares, L.; Gómez-González, M.; Uroz, M.; Valon, L.; Nair, R.V.; Garreta, E.; Montserrat, N.; del Campo, A.; et al. Active superelasticity in three-dimensional epithelia of controlled shape. Nature 2018, 563, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Pérez-González, C.; Ceada, G.; Greco, F.; Matejčić, M.; Gómez-González, M.; Castro, N.; Menendez, A.; Kale, S.; Krndija, D.; Clark, A.G.; et al. Mechanical compartmentalization of the intestinal organoid enables crypt folding and collective cell migration. Nat. Cell Biol. 2021, 23, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gálvez, P.; Vicente-Munuera, P.; Anbari, S.; Buceta, J.; Escudero, L.M. The Complex Three-Dimensional Organization of Epithelial Tissues. Development 2021, 148, dev195669. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gálvez, P.; Vicente-Munuera, P.; Anbari, S.; Tagua, A.; Gordillo-Vázquez, C.; Andrés-San Román, J.A.; Franco-Barranco, D.; Palacios, A.M.; Velasco, A.; Capitán-Agudo, C.; et al. A quantitative biophysical principle to explain the 3D cellular connectivity in curved epithelia. Cell Syst. 2022, 13, 631–643.e8. [Google Scholar] [CrossRef] [PubMed]
- Fulton, T.; Verd, B.; Steventon, B. The unappreciated generative role of cell movements in pattern formation. R. Soc. Open Sci. 2022, 9, 211293. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, M.A.; Alert, R.; LaChance, J.M.; Zajdel, T.J.; Košmrlj, A.; Cohen, D.J. Size-dependent patterns of cell proliferation and migration in freely-expanding epithelia. eLife 2020, 9, e58945. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Ciudad, I. Tooth morphogenesis in vivo, in vitro, and in silico. Curr. Top. Dev. Biol. 2008, 81, 341–371. [Google Scholar] [CrossRef]
- Gritti, N.; Oriola, D.; Trivedi, V. Rethinking embryology in vitro: A synergy between engineering, data science and theory. Dev. Biol. 2021, 474, 48–61. [Google Scholar] [CrossRef]
- Matejčić, M.; Trepat, X. Mechanobiological approaches to synthetic morphogenesis: Learning by building. Trends Cell Biol. 2022, 33, 95–111. [Google Scholar] [CrossRef]
- Mongera, A.; Rowghanian, P.; Gustafson, H.J.; Shelton, E.; Kealhofer, D.A.; Carn, E.K.; Serwane, F.; Lucio, A.A.; Giammona, J.; Campàs, O. A fluid-to-solid jamming transition underlies vertebrate body axis elongation. Nature 2018, 561, 401–405. [Google Scholar] [CrossRef]
- Kim, S.; Pochitaloff, M.; Stooke-Vaughan, G.A.; Campàs, O. Embryonic tissues as active foams. Nat. Phys. 2021, 17, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Banavar, S.P.; Carn, E.K.; Rowghanian, P.; Stooke-Vaughan, G.; Kim, S.; Campàs, O. Mechanical control of tissue shape and morphogenetic flows during vertebrate body axis elongation. Sci. Rep. 2021, 11, 8591. [Google Scholar] [CrossRef] [PubMed]
- Samson, S.C.; Khan, A.M.; Mendoza, M.C. ERK signaling for cell migration and invasion. Front. Mol. Biosci. 2022, 9, 998475. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.; Kondo, Y.; Naoki, H.; Hiratsuka, T.; Itoh, R.E.; Matsuda, M. Propagating Wave of ERK Activation Orients Collective Cell Migration. Dev. Cell 2017, 43, 305–317.e5. [Google Scholar] [CrossRef] [PubMed]
- Hino, N.; Matsuda, K.; Jikko, Y.; Maryu, G.; Sakai, K.; Imamura, R.; Tsukiji, S.; Aoki, K.; Terai, K.; Hirashima, T.; et al. A feedback loop between lamellipodial extension and HGF-ERK signaling specifies leader cells during collective cell migration. Dev. Cell 2022, 57, 2290–2304.e7. [Google Scholar] [CrossRef]
- Boocock, D.; Hino, N.; Ruzickova, N.; Hirashima, T.; Hannezo, E. Theory of mechanochemical patterning and optimal migration in cell monolayers. Nat. Phys. 2020, 17, 267–274. [Google Scholar] [CrossRef]
- Fletcher, A.G.; Osborne, J.M. Seven challenges in the multiscale modeling of multicellular tissues. WIREs Mech. Dis. 2022, 14. [Google Scholar] [CrossRef]
- Graner, F.; Glazier, J. Simulation of biological cell sorting using a two-dimensional extended Potts model. Phys. Rev. Lett. 1992, 69, 2013–2016. [Google Scholar] [CrossRef]
- Izaguirre, J.A.; Chaturvedi, R.; Huang, C.; Cickovski, T.; Coffland, J.; Thomas, G.; Forgacs, G.; Alber, M.; Hentschel, G.; Newman, S.A.; et al. CompuCell, a multi-model framework for simulation of morphogenesis. Bioinformatics 2004, 20, 1129–1137. [Google Scholar] [CrossRef]
- Fletcher, A.G.; Osterfield, M.; Baker, R.E.; Shvartsman, S.Y. Vertex models of epithelial morphogenesis. Biophys. J. 2014, 106, 2291–2304. [Google Scholar] [CrossRef]
- Ishimoto, Y.; Morishita, Y. Bubbly vertex dynamics: A dynamical and geometrical model for epithelial tissues with curved cell shapes. Phys. Rev. E 2014, 90, 052711. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Gutiérrez, D.; Tozluoglu, M.; Barry, J.D.; Pascual, A.; Mao, Y.; Escudero, L.M. Fundamental physical cellular constraints drive self-organization of tissues. EMBO J. 2016, 35, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gálvez, P.; Vicente-Munuera, P.; Tagua, A.; Forja, C.; Castro, A.M.; Letrán, M.; Valencia-Expósito, A.; Grima, C.; Bermúdez-Gallardo, M.; Serrano-Pérez-Higueras, Ó.; et al. Scutoids are a geometrical solution to three-dimensional packing of epithelia. Nat. Commun. 2018, 9, 2960. [Google Scholar] [CrossRef] [PubMed]
- Canela-Xandri, O.; Anbari, S.; Buceta, J. TiFoSi: An efficient tool for mechanobiology simulations of epithelia. Bioinformatics 2020, 36, 4525–4526. [Google Scholar] [CrossRef] [PubMed]
- Durney, C.H.; Feng, J.J. A three-dimensional vertex model for Drosophila salivary gland invagination. Phys. Biol. 2021, 18, 046005. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez Cerro, Á.; Sancho, S.; Rodríguez, M.; Gamón, M.A.; Guitou, L.; Martínez, R.J.; Buceta, J. ANISE: An application to design mechanobiology simulations of planar epithelia. Bioinformatics 2022, 38, 4246–4247. [Google Scholar] [CrossRef]
- Andrés-San Román, J.A.; Gordillo-Vázquez, C.; Franco-Barranco, D.; Morato, L.; Tagua, A.; Vicente-Munuera, P.; Palacios, A.M.; Gavilán, M.P.; Annese, V.; Gómez-Gálvez, P.; et al. CartoCell, a high-throughput pipeline for accurate 3D image analysis, unveils cell morphology patterns in epithelial cysts. bioRxiv 2023. [Google Scholar] [CrossRef]
- Schamberger, B.; Roschger, A.; Ziege, R.; Anselme, K.; Amar, M.B.; Bykowski, M.; Castro, A.P.G.; Cipitria, A.; Coles, R.; Dimova, R.; et al. Curvature in Biological Systems: Its quantification, Emergence and Implications Across the Scales. Adv. Mater. 2023, 35, 2206110. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Koo, B.K.; Knoblich, J.A. Human organoids: Model systems for human biology and medicine. Nat. Rev. Mol. Cell Biol. 2020, 21, 571–584. [Google Scholar] [CrossRef]
- Vives, J.; Batlle-Morera, L. The challenge of developing human 3D organoids into medicines. Stem Cell Res. Ther. 2020, 11, 72. [Google Scholar] [CrossRef]
- Huch, M.; Knoblich, J.A.; Lutolf, M.P.; Martinez-Arias, A. The hope and the hype of organoid research. Development 2017, 144, 938–941. [Google Scholar] [CrossRef]
- Moris, N.; Anlas, K.; van den Brink, S.C.; Alemany, A.; Schröder, J.; Ghimire, S.; Balayo, T.; van Oudenaarden, A.; Martinez Arias, A. An in vitro model of early anteroposterior organization during human development. Nature 2020, 582, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Gjorevski, N.; Nikolaev, M.; Brown, T.E.; Mitrofanova, O.; Brandenberg, N.; DelRio, F.W.; Yavitt, F.M.; Liberali, P.; Anseth, K.S.; Lutolf, M.P. Tissue geometry drives deterministic organoid patterning. Science 2022, 375, eaaw9021. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, Y.; Hamidi, S.; Yoshioka-Kobayashi, K.; Munira, S.; Sunadome, K.; Zhang, Y.; Kurokawa, Y.; Ericsson, R.; Mieda, A.; Thompson, J.L.; et al. Reconstituting human somitogenesis in vitro. Nature 2023, 614, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Parrondo, J.M.R.; van den Broeck, C.; Buceta, J.; de la Rubia, F.J. Noise-induced spatial patterns. Phys. A Stat. Mech. Its Appl. 1996, 224, 153–161. [Google Scholar] [CrossRef]
- Buceta, J.; Lindenberg, K.; Parrondo, J.M.R. Stationary and Oscillatory Spatial Patterns Induced by Global Periodic Switching. Phys. Rev. Lett. 2001, 88, 024103. [Google Scholar] [CrossRef]
- Buceta, J.; Lindenberg, K.; Parrondo, J.M.R. Pattern formation induced by nonequilibrium global alternation of dynamics. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2002, 66, 036216. [Google Scholar] [CrossRef]
- Buceta, J.; Ibañes, M.; Sancho, J.M.; Lindenberg, K. Noise-Driven Mechanism for Pattern Formation. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2002, 307. [Google Scholar] [CrossRef]
- Buceta, J.; Lindenberg, K. Switching-induced Turing instability. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2002, 66, 046202. [Google Scholar] [CrossRef]
- Buceta, J.; Lindenberg, K. Spatial Patterns Induced Purely by Dichotomous Disorder. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2003, 68, 011103. [Google Scholar] [CrossRef]
- Wood, K.; Buceta, J.; Lindenberg, K. Comprehensive study of pattern formation in relaxational systems. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2006, 73, 022101. [Google Scholar] [CrossRef] [PubMed]
- Buceta, J.; Lindenberg, K.; Parrondo, J.M.R. Spatial patterns induced by random switching. In The Random and Fluctuating World; World Scientific: Singapore, 2022; pp. 203–211. [Google Scholar] [CrossRef]
- Bollenbach, T.; Kruse, K.; Pantazis, P.; González-Gaitán, M.; Jülicher, F. Robust formation of morphogen gradients. Phys. Rev. Lett. 2005, 94, 018103. [Google Scholar] [CrossRef] [PubMed]
- Gregor, T.; Tank, D.W.; Wieschaus, E.F.; Bialek, W. Probing the limits to positional information. Cell 2007, 130, 153–164. [Google Scholar] [CrossRef]
- Bollenbach, T.; Pantazis, P.; Kicheva, A.; Bökel, C.; González-Gaitán, M.; Jülicher, F. Precision of the Dpp gradient. Development 2008, 135, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Rudge, T.; Burrage, K. Effects of intrinsic and extrinsic noise can accelerate juxtacrine pattern formation. Philos. Trans. R. Soc. Lond. Ser. B 2008, 70, 971–991. [Google Scholar] [CrossRef] [PubMed]
- Saunders, T.E.; Howard, M. Morphogen profiles can be optimized to buffer against noise. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2009, 80, 041902. [Google Scholar] [CrossRef] [PubMed]
- Tkačik, G.; Dubuis, J.O.; Petkova, M.D.; Gregor, T. Positional Information, Positional Error, and Readout Precision in Morphogenesis: A Mathematical Framework. Genetics 2015, 199, 39–59. [Google Scholar] [CrossRef]
- Belousov, R.; Jacobo, A.; Hudspeth, A.J. Fluctuation theory in space and time: White noise in reaction-diffusion models of morphogenesis. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2018, 98, 052125. [Google Scholar] [CrossRef]
- Exelby, K.; Herrera-Delgado, E.; Perez, L.G.; Perez-Carrasco, R.; Sagner, A.; Metzis, V.; Sollich, P.; Briscoe, J. Precision of tissue patterning is controlled by dynamical properties of gene regulatory networks. Development 2021, 148, dev197566. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buceta, J.; Guitou, L. Developmental Pattern Formation: Spanish Contributions from a Biophysical Perspective. Biophysica 2023, 3, 335-347. https://doi.org/10.3390/biophysica3020022
Buceta J, Guitou L. Developmental Pattern Formation: Spanish Contributions from a Biophysical Perspective. Biophysica. 2023; 3(2):335-347. https://doi.org/10.3390/biophysica3020022
Chicago/Turabian StyleBuceta, Javier, and Léna Guitou. 2023. "Developmental Pattern Formation: Spanish Contributions from a Biophysical Perspective" Biophysica 3, no. 2: 335-347. https://doi.org/10.3390/biophysica3020022
APA StyleBuceta, J., & Guitou, L. (2023). Developmental Pattern Formation: Spanish Contributions from a Biophysical Perspective. Biophysica, 3(2), 335-347. https://doi.org/10.3390/biophysica3020022