
Community Responses to Fire: A Global Meta-Analysis Unravels the Contrasting Responses of Fauna to Fire


Abstract
:1. Introduction
2. Methods
2.1. Article Search
2.2. Inclusion Criteria and Screening
2.3. Data Extraction
2.4. Effect Size Calculation and Analysis
2.5. Publication Bias

2.6. Distribution of Data in the Literature
3. Results
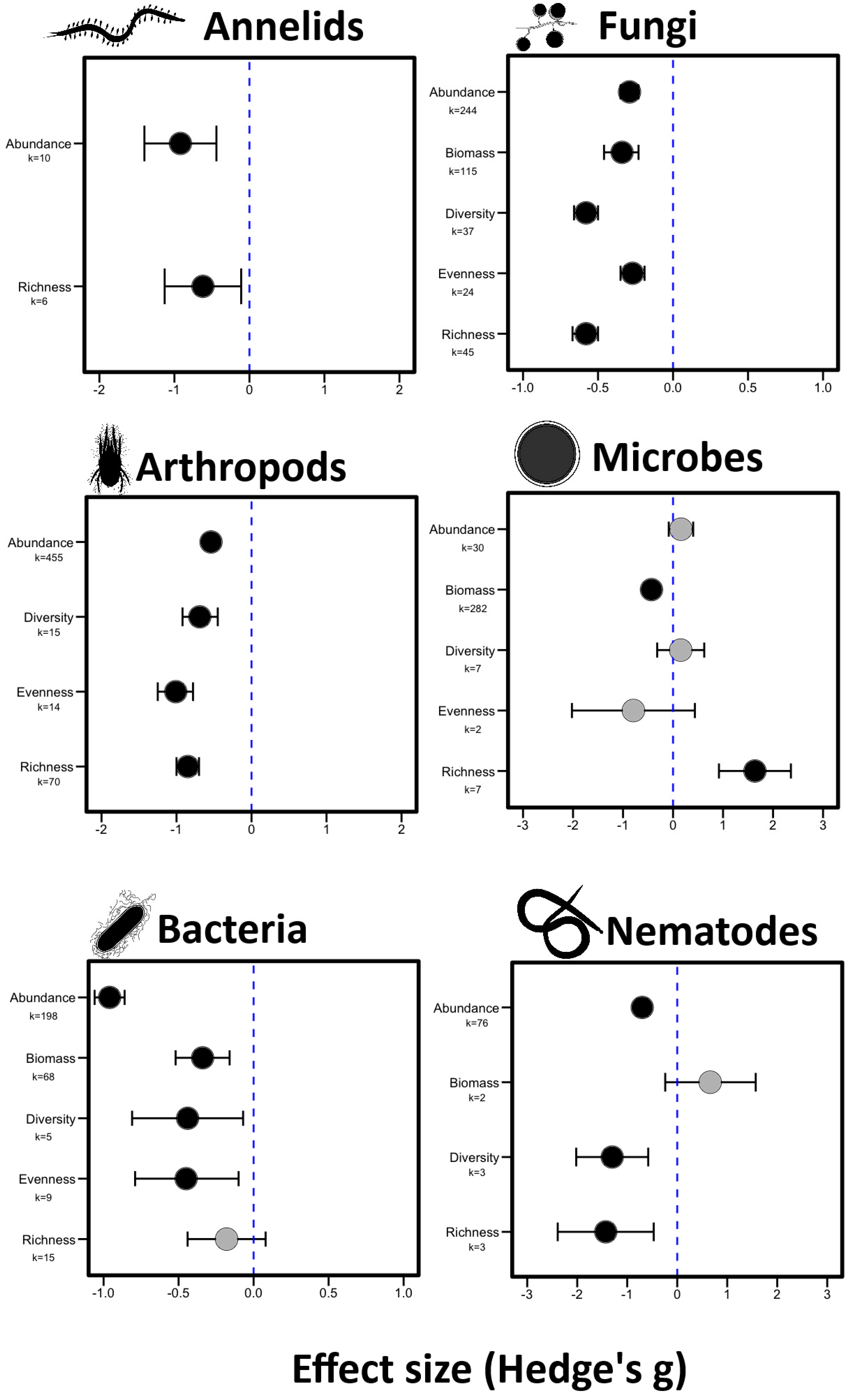
3.1. Effect of Fire on Aboveground and Belowground Fuana (Question 1)




3.2. Effect of Fire Types and Intensity on Organisms (Question 2)
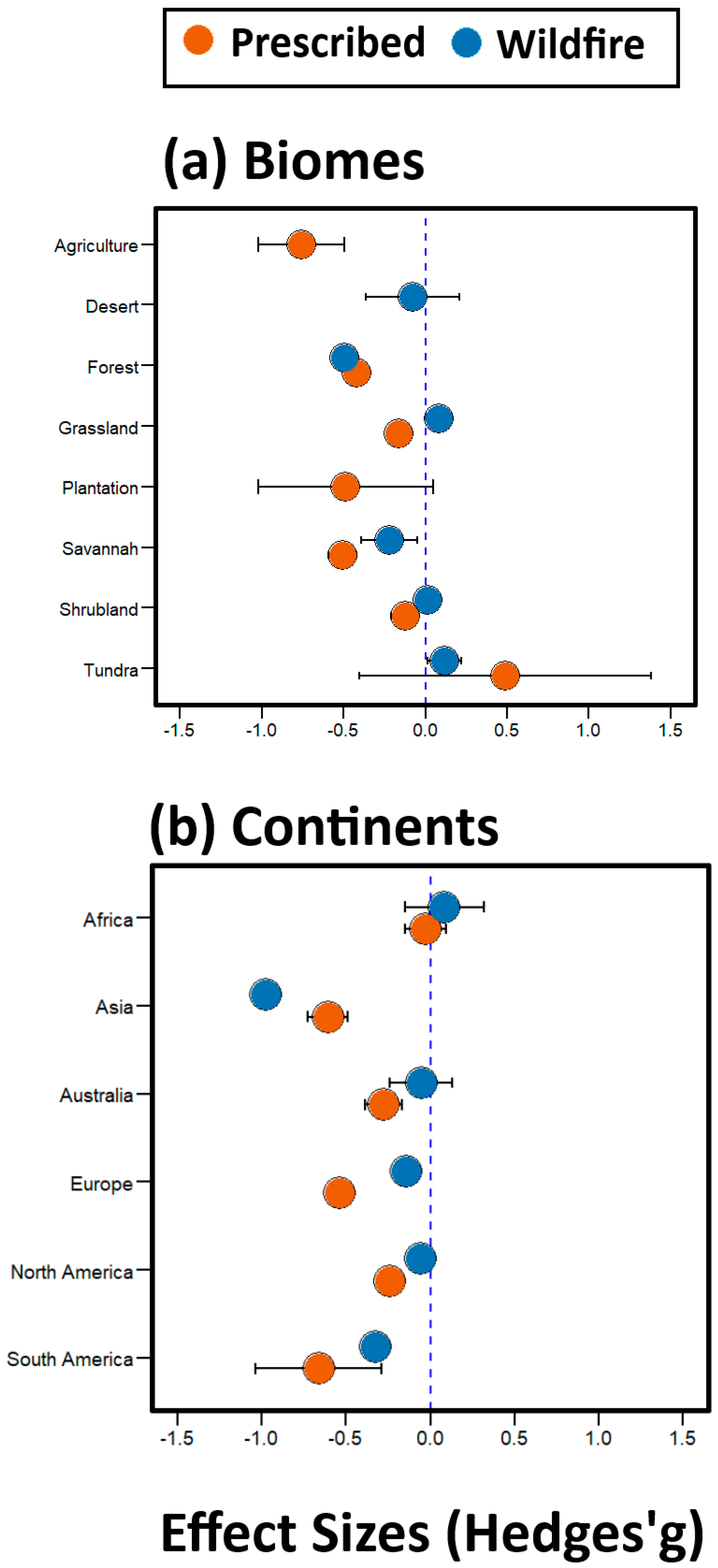
3.3. Global Effect of Fires across Biomes and Continents (Question 3)
3.4. Relationship between Abiotic Factors and Fire Effects (Question 4)
4. Discussion
4.1. Belowground Fauna Are More Susceptible to Fire Than Aboveground Fauna
4.2. Fire Types and Intensities Negatively Affect Organisms
4.3. Fire Effects Are More Pronounced in Certain Biomes and Continents
4.4. Abiotic Factors Do Not Predict Fire Effects Well
4.5. Future Research Needs in Fire Ecology
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Badejo, M.A. Effect of Accidental Fire on Soil Mite Density in a Forest Reserve in Nigeria. Exp. Appl. Acarol. 1994, 18, 703–710. [Google Scholar]
- Abom, R.; Schwarzkopf, L. Short-Term Responses of Reptile Assemblages to Fire in Native and Weedy Tropical Savannah. Glob. Ecol. Conserv. 2016, 6, 58–66. [Google Scholar] [CrossRef]
- Anjos, D.; Campos, R.; Campos, R.; Ribeiro, S. Monitoring Effect of Fire on Ant Assemblages in Brazilian Rupestrian Grasslands: Contrasting Effects on Ground and Arboreal Fauna. Insects 2017, 8, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, O.M.; Lewis, T.; Rezaei Rashti, M.; Chen, C. Long-Term Fire Regime Modifies Carbon and Nutrient Dynamics in Decomposing Eucalyptus Pilularis Leaf Litter. Front. For. Glob. Change 2020, 3, 22. [Google Scholar] [CrossRef] [Green Version]
- Butler, A.; Davis, C.A.; Fuhlendorf, S.D.; Wilder, S.M. Effects of Fire on Ground-dwelling Arthropods in a Shrub-dominated Grassland. Ecol. Evol. 2021, 11, 427–442. [Google Scholar] [CrossRef] [PubMed]
- Cadena-Zamudio, D.; Ruiz-Guerra, B.; Castillo, M.L.; Flores-Garnica, J.G.; Guevara, R. Prevalence of Stochastic Processes in the Fire-Mediated Reassemblage of the Soil Arthropod Community of a Pine Forest. Acta Oecologica 2022, 115, 103834. [Google Scholar] [CrossRef]
- Marlon, J.R.; Bartlein, P.J.; Carcaillet, C.; Gavin, D.G.; Harrison, S.P.; Higuera, P.E.; Joos, F.; Power, M.J.; Prentice, I.C. Climate and Human Influences on Global Biomass Burning over the Past Two Millennia. Nat. Geosci 2008, 1, 697–702. [Google Scholar] [CrossRef]
- Zennaro, P.; Kehrwald, N.; Marlon, J.; Ruddiman, W.F.; Brücher, T.; Agostinelli, C.; Dahl-Jensen, D.; Zangrando, R.; Gambaro, A.; Barbante, C. Europe on Fire Three Thousand Years Ago: Arson or Climate? Geophys. Res. Lett. 2015, 42, 2023–2033. [Google Scholar] [CrossRef]
- Turco, M.; von Hardenberg, J.; AghaKouchak, A.; Llasat, M.C.; Provenzale, A.; Trigo, R.M. On the Key Role of Droughts in the Dynamics of Summer Fires in Mediterranean Europe. Sci. Rep. 2017, 7, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessilt, T.D.; Abatzoglou, J.T.; Chen, Y.; Randerson, J.T.; Scholten, R.C.; van der Werf, G.; Veraverbeke, S. Future Increases in Lightning Ignition Efficiency and Wildfire Occurrence Expected from Drier Fuels in Boreal Forest Ecosystems of Western North America. Environ. Res. Lett. 2022, 17, 054008. [Google Scholar] [CrossRef]
- van Wagtendonk, J.W.; Cayan, D.R. Temporal and Spatial Distribution of Lightning Strikes in California in Relation to Large-Scale Weather Patterns. Fire Ecol. 2008, 4, 34–56. [Google Scholar] [CrossRef]
- Just, M.G.; Hohmann, M.G.; Hoffmann, W.A. Where Fire Stops: Vegetation Structure and Microclimate Influence Fire Spread along an Ecotonal Gradient. Plant Ecol. 2016, 217, 631–644. [Google Scholar] [CrossRef]
- Yuan, Z.; Wang, Y.; Xu, J.; Wu, Z. Effects of Climatic Factors on the Net Primary Productivity in the Source Region of Yangtze River, China. Sci. Rep. 2021, 11, 1376. [Google Scholar] [CrossRef] [PubMed]
- Jolly, W.M.; Cochrane, M.A.; Freeborn, P.H.; Holden, Z.A.; Brown, T.J.; Williamson, G.J.; Bowman, D.M.J.S. Climate-Induced Variations in Global Wildfire Danger from 1979 to 2013. Nat. Commun. 2015, 6, 7537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and Earlier Spring Increase Western U.S. Forest Wildfire Activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef] [Green Version]
- Fawzy, S.; Osman, A.I.; Doran, J.; Rooney, D.W. Strategies for Mitigation of Climate Change: A Review. Environ. Chem Lett 2020, 18, 2069–2094. [Google Scholar] [CrossRef]
- IPCC Summary for Policymakers. Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P.R., Skea, J., Calvo Buendia, E., Masson-Delmotte, V., Pörtner, H.-O., Roberts, D.C., Zhai, P., Slade, R., Connors, S., van Diemen, R., Eds.; IPCC: Geneva, Switzerland, 2022. [Google Scholar]
- Abram, N.J.; Henley, B.J.; Sen Gupta, A.; Lippmann, T.J.R.; Clarke, H.; Dowdy, A.J.; Sharples, J.J.; Nolan, R.H.; Zhang, T.; Wooster, M.J.; et al. Connections of Climate Change and Variability to Large and Extreme Forest Fires in Southeast Australia. Commun Earth Env. 2021, 2, 1–17. [Google Scholar] [CrossRef]
- Doamba, S.W.M.F.; Savadogo, P.; Nacro, H.B. Effects of Burning on Soil Macrofauna in a Savanna-Woodland under Different Experimental Fuel Load Treatments. Appl. Soil Ecol. 2014, 81, 37–44. [Google Scholar] [CrossRef]
- Gongalsky, K.B.; Persson, T. Recovery of Soil Macrofauna after Wildfires in Boreal Forests. Soil Biol. Biochem. 2013, 57, 182–191. [Google Scholar] [CrossRef]
- Underwood, E.C.; Quinn, J.F. Response of Ants and Spiders to Prescribed Fire in Oak Woodlands of California. J. Insect Conserv. 2010, 14, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Ross, P.M.; Harvey, K.; Vecchio, E.M.; Beckers, D. Impact of Fire and the Recovery of Molluscs in South-East Australian Salt Marsh. Ecol. Manag. Restor. 2019, 20, 126–135. [Google Scholar] [CrossRef]
- Furukori, N.; Kishimoto-Yamada, K.; Homma, K. Impacts of Burning and Herbicide Disturbances on Soil Animals and Organic Matter Decomposition in Terraced Paddy Field Levees in Japanese Satoyama. J. Soil Sci. Plant Nutr. 2022, 22, 270–280. [Google Scholar] [CrossRef]
- Roces-Díaz, J.V.; Santín, C.; Martínez-Vilalta, J.; Doerr, S.H. A Global Synthesis of Fire Effects on Ecosystem Services of Forests and Woodlands. Front. Ecol Environ. 2022, 20, 170–178. [Google Scholar] [CrossRef]
- Viljur, M.; Abella, S.R.; Adámek, M.; Alencar, J.B.R.; Barber, N.A.; Beudert, B.; Burkle, L.A.; Cagnolo, L.; Campos, B.R.; Chao, A.; et al. The Effect of Natural Disturbances on Forest Biodiversity: An Ecological Synthesis. Biol. Rev. 2022, 97, 1930–1947. [Google Scholar] [CrossRef] [PubMed]
- Pennington, R.T.; Lehmann, C.E.R.; Rowland, L.M. Tropical Savannas and Dry Forests. Curr. Biol. 2018, 28, R541–R545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solofondranohatra, C.L.; Vorontsova, M.S.; Hackel, J.; Besnard, G.; Cable, S.; Williams, J.; Jeannoda, V.; Lehmann, C.E.R. Grass Functional Traits Differentiate Forest and Savanna in the Madagascar Central Highlands. Front. Ecol. Evol. 2018, 6, 184. [Google Scholar] [CrossRef] [Green Version]
- Ratnam, J.; Bond, W.J.; Fensham, R.J.; Hoffmann, W.A.; Archibald, S.; Lehmann, C.E.R.; Anderson, M.T.; Higgins, S.I.; Sankaran, M. When Is a ‘Forest’ a Savanna, and Why Does It Matter? Glob. Ecol. Biogeogr. 2011, 20, 653–660. [Google Scholar] [CrossRef]
- Coyle, D.R.; Nagendra, U.J.; Taylor, M.K.; Campbell, J.H.; Cunard, C.E.; Joslin, A.H.; Mundepi, A.; Phillips, C.A.; Callaham, M.A. Soil Fauna Responses to Natural Disturbances, Invasive Species, and Global Climate Change: Current State of the Science and a Call to Action. Soil Biol. Biochem. 2017, 110, 116–133. [Google Scholar] [CrossRef]
- Pressler, Y.; Moore, J.C.; Cotrufo, M.F. Belowground Community Responses to Fire: Meta-Analysis Reveals Contrasting Responses of Soil Microorganisms and Mesofauna. Oikos 2019, 128, 309–327. [Google Scholar] [CrossRef]
- Santín, C.; Doerr, S.H. Fire Effects on Soils: The Human Dimension. Philos. Trans. R Soc. Lond. B Biol. Sci. 2016, 371, 20150171. [Google Scholar] [CrossRef] [Green Version]
- Haimi, J.; Fritze, H.; Moilanen, P. Responses of Soil Decomposer Animals to Wood-Ash Fertilisation and Burning in a Coniferous Forest Stand. For. Ecol. Manag. 2000, 129, 53–61. [Google Scholar] [CrossRef]
- Pellegrini, A.F.A.; Hobbie, S.E.; Reich, P.B.; Jumpponen, A.; Brookshire, E.N.J.; Caprio, A.C.; Coetsee, C.; Jackson, R.B. Repeated Fire Shifts Carbon and Nitrogen Cycling by Changing Plant Inputs and Soil Decomposition across Ecosystems. Ecol. Monogr. 2020, 90, e01409. [Google Scholar] [CrossRef]
- Cassagne, N.; Pimont, F.; Dupuy, J.-L.; Linn, R.R.; Mårell, A.; Oliveri, C.; Rigolot, E. Using a Fire Propagation Model to Assess the Efficiency of Prescribed Burning in Reducing the Fire Hazard. Ecol. Model. 2011, 222, 1502–1514. [Google Scholar] [CrossRef]
- Fernandes, P.M.; Botelho, H.S. A Review of Prescribed Burning Effectiveness in Fire Hazard Reduction. Int. J. Wildland Fire 2003, 12, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Ponisio, L.C.; Wilkin, K.; M’Gonigle, L.K.; Kulhanek, K.; Cook, L.; Thorp, R.; Griswold, T.; Kremen, C. Pyrodiversity Begets Plant–Pollinator Community Diversity. Glob. Change Biol. 2016, 22, 1794–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parr, C.L.; Brockett, B.H. Patch-Mosaic Burning: A New Paradigm for Savanna Fire Management in Protected Areas? Koedoe Afr. Prot. Area Conserv. Sci. 1999, 42, 117–130. [Google Scholar] [CrossRef] [Green Version]
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brockett, B.H.; Biggs, H.C.; Wilgen, B.W. van A Patch Mosaic Burning System for Conservation Areas in Southern African Savannas. Int. J. Wildland Fire 2001, 10, 169–183. [Google Scholar] [CrossRef]
- Jones, G.M.; Tingley, M.W. Pyrodiversity and Biodiversity: A History, Synthesis, and Outlook. Divers. Distrib. 2022, 28, 386–403. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Perry, G.L.W.; Higgins, S.I.; Johnson, C.N.; Fuhlendorf, S.D.; Murphy, B.P. Pyrodiversity Is the Coupling of Biodiversity and Fire Regimes in Food Webs. Philos. Trans. R Soc Lond. B Biol. Sci. 2016, 371, 20150169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwilk, D.W.; Keeley, J.E.; Bond, W.J. The Intermediate Disturbance Hypothesis Does Not Explain Fire and Diversity Pattern in Fynbos. Plant Ecol. 1997, 132, 77–84. [Google Scholar] [CrossRef]
- Pastro, L.A.; Dickman, C.R.; Letnic, M. Burning for Biodiversity or Burning Biodiversity? Prescribed Burn vs. Wildfire Impacts on Plants, Lizards, and Mammals. Ecol. Appl. 2011, 21, 3238–3253. [Google Scholar] [CrossRef]
- Mooney, S.D.; Harrison, S.P.; Bartlein, P.J.; Daniau, A.-L.; Stevenson, J.; Brownlie, K.C.; Buckman, S.; Cupper, M.; Luly, J.; Black, M.; et al. Late Quaternary Fire Regimes of Australasia. Quat. Sci. Rev. 2011, 30, 28–46. [Google Scholar] [CrossRef]
- Durigan, G.; Pilon, N.A.L.; Abreu, R.C.R.; Hoffmann, W.A.; Martins, M.; Fiorillo, B.F.; Antunes, A.Z.; Carmignotto, A.P.; Maravalhas, J.B.; Vieira, J.; et al. No Net Loss of Species Diversity After Prescribed Fires in the Brazilian Savanna. Front. For. Glob. Change 2020, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Giorgis, M.A.; Zeballos, S.R.; Carbone, L.; Zimmermann, H.; von Wehrden, H.; Aguilar, R.; Ferreras, A.E.; Tecco, P.A.; Kowaljow, E.; Barri, F.; et al. A Review of Fire Effects across South American Ecosystems: The Role of Climate and Time since Fire. Fire Ecol. 2021, 17, 11. [Google Scholar] [CrossRef]
- Robinson, N.M.; Leonard, S.W.J.; Ritchie, E.G.; Bassett, M.; Chia, E.K.; Buckingham, S.; Gibb, H.; Bennett, A.F.; Clarke, M.F. REVIEW: Refuges for Fauna in Fire-Prone Landscapes: Their Ecological Function and Importance. J. Appl. Ecol. 2013, 50, 1321–1329. [Google Scholar] [CrossRef]
- Najbar, A.; Konowalik, A.; Halupka, K.; Najbar, B.; Ogielska, M. Body Size and Life History Traits of the Fire Salamander Salamandra Salamandra from Poland. Amphib. -Reptil. 2020, 41, 63–74. [Google Scholar] [CrossRef]
- González, T.M.; González-Trujillo, J.D.; Muñoz, A.; Armenteras, D. Differential Effects of Fire on the Occupancy of Small Mammals in Neotropical Savanna-Gallery Forests. Perspect. Ecol. Conserv. 2021, 19, 179–188. [Google Scholar] [CrossRef]
- Malmström, A. Life-History Traits Predict Recovery Patterns in Collembola Species after Fire: A 10 Year Study. Appl. Soil Ecol. 2012, 56, 35–42. [Google Scholar] [CrossRef]
- Álvarez, G.; Ammagarahalli, B.; Hall, D.R.; Pajares, J.A.; Gemeno, C. Smoke, Pheromone and Kairomone Olfactory Receptor Neurons in Males and Females of the Pine Sawyer Monochamus Galloprovincialis (Olivier) (Coleoptera: Cerambycidae). J. Insect Physiol. 2015, 82, 46–55. [Google Scholar] [CrossRef]
- Grafe, T.U.; Döbler, S.; Linsenmair, K.E. Frogs Flee from the Sound of Fire. Proc. Biol. Sci. 2002, 269, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Klocke, D.; Schmitz, A.; Soltner, H.; Bousack, H.; Schmitz, H. Infrared Receptors in Pyrophilous (“Fire Loving”) Insects as Model for New Un-Cooled Infrared Sensors. Beilstein J. Nanotechnol. 2011, 2, 186–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, H.; Schmitz, A.; Schneider, E.S. Matched Filter Properties of Infrared Receptors Used for Fire and Heat Detection in Insects. In The Ecology of Animal Senses: Matched Filters for Economical Sensing; von der Emde, G., Warrant, E., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 207–234. ISBN 978-3-319-25492-0. [Google Scholar]
- Dell, J.; O’Brien, J.; Doan, L.; Richards, L.; Dyer, L. An Arthropod Survival Strategy in a Frequently Burned Forest. Ecology 2017, 98, 2972–2974. [Google Scholar] [CrossRef] [PubMed]
- Korobushkin, D.I.; Gorbunova, A.Yu.; Zaitsev, A.S.; Gongalsky, K.B. Trait-Specific Response of Soil Macrofauna to Forest Burning along a Macrogeographic Gradient. Appl. Soil Ecol. 2017, 112, 97–100. [Google Scholar] [CrossRef]
- Carbone, L.M.; Tavella, J.; Pausas, J.G.; Aguilar, R. A Global Synthesis of Fire Effects on Pollinators. Glob. Ecol. Biogeogr. 2019, 28, 1487–1498. [Google Scholar] [CrossRef]
- Peralta, G.; Stevani, E.L.; Chacoff, N.P.; Dorado, J.; Vázquez, D.P. Fire Influences the Structure of Plant–Bee Networks. J. Anim. Ecol. 2017, 86, 1372–1379. [Google Scholar] [CrossRef] [Green Version]
- García, Y.; Clara Castellanos, M.; Pausas, J.G. Differential Pollinator Response Underlies Plant Reproductive Resilience after Fires. Ann. Bot. 2018, 122, 961–971. [Google Scholar] [CrossRef]
- Mola, J.M.; Miller, M.R.; O’Rourke, S.M.; Williams, N.M. Wildfire Reveals Transient Changes to Individual Traits and Population Responses of a Native Bumble Bee Bombus Vosnesenskii. J. Anim. Ecol. 2020, 89, 1799–1810. [Google Scholar] [CrossRef]
- Karpestam, E.; Merilaita, S.; Forsman, A. Reduced Predation Risk for Melanistic Pygmy Grasshoppers in Post-Fire Environments. Ecol. Evol. 2012, 2, 2204–2212. [Google Scholar] [CrossRef]
- Kelly, L.T.; Brotons, L.; Giljohann, K.M.; McCarthy, M.A.; Pausas, J.G.; Smith, A.L. Bridging the Divide: Integrating Animal and Plant Paradigms to Secure the Future of Biodiversity in Fire-Prone Ecosystems. Fire 2018, 1, 29. [Google Scholar] [CrossRef] [Green Version]
- Dove, N.C.; Hart, S.C. Fire Reduces Fungal Species Richness and In Situ Mycorrhizal Colonization: A Meta-Analysis. Fire Ecol. 2017, 13, 37–65. [Google Scholar] [CrossRef]
- Knight, T.M.; Holt, R.D. Fire Generates Spatial Gradients in Herbivory: An Example from a Florida Sandhill Ecosystem. Ecology 2005, 86, 587–593. [Google Scholar] [CrossRef] [Green Version]
- Geary, W.L.; Doherty, T.S.; Nimmo, D.G.; Tulloch, A.I.T.; Ritchie, E.G. Predator Responses to Fire: A Global Systematic Review and Meta-Analysis. J. Anim. Ecol. 2020, 89, 955–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doherty, T.S.; Geary, W.L.; Jolly, C.J.; Macdonald, K.J.; Miritis, V.; Watchorn, D.J.; Cherry, M.J.; Conner, L.M.; González, T.M.; Legge, S.M.; et al. Fire as a Driver and Mediator of Predator-Prey Interactions. Biol. Rev. Camb. Philos. Soc. 2022, 97, 1539–1558. [Google Scholar] [CrossRef] [PubMed]
- Letnic, M.; Tischler, M.; Gordon, C. Desert Small Mammal Responses to Wildfire and Predation in the Aftermath of a La Nińa Driven Resource Pulse. Austral Ecol. 2013, 38, 841–849. [Google Scholar] [CrossRef]
- McLauchlan, K.K.; Higuera, P.E.; Miesel, J.; Rogers, B.M.; Schweitzer, J.; Shuman, J.K.; Tepley, A.J.; Varner, J.M.; Veblen, T.T.; Adalsteinsson, S.A.; et al. Fire as a Fundamental Ecological Process: Research Advances and Frontiers. J. Ecol. 2020, 108, 2047–2069. [Google Scholar] [CrossRef]
- Burrows, N.D.; Friend, G. Biological Indicators of Appropriate Fire Regimes in Southwest Australian Ecosystems. In Fire in Ecosystem Management: Shifting the Paradigm from Suppression to Prescription; Tall Timbers Fire Ecology Conference Proceedings, No. 20; Tall Timbers Research Station: Tallahassee, Florida, 1998; pp. 413–421. [Google Scholar]
- Solascasas, P.; Azcárate, F.M.; Hevia, V. Edaphic Arthropods as Indicators of the Ecological Condition of Temperate Grassland Ecosystems: A Systematic Review. Ecol. Indic. 2022, 142, 109277. [Google Scholar] [CrossRef]
- Mantoni, C.; Di Musciano, M.; Fattorini, S. Use of Microarthropods to Evaluate the Impact of Fire on Soil Biological Quality. J. Environ. Manag. 2020, 266, 110624. [Google Scholar] [CrossRef]
- Lacki, M.J.; Cox, D.R.; Dodd, L.E.; Dickinson, M.B. Response of Northern Bats (Myotis Septentrionalis) to Prescribed Fires in Eastern Kentucky Forests. J. Mammal. 2009, 90, 1165–1175. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.G.; Tangney, R.; Enright, N.J.; Fontaine, J.B.; Merritt, D.J.; Ooi, M.K.J.; Ruthrof, K.X.; Miller, B.P. Mechanisms of Fire Seasonality Effects on Plant Populations. Trends Ecol. Evol. 2019, 34, 1104–1117. [Google Scholar] [CrossRef]
- Keith, D.A.; Dunker, B.; Driscoll, D.A. Dispersal: The Eighth Fire Seasonality Effect on Plants. Trends Ecol. Evol. 2020, 35, 305–307. [Google Scholar] [CrossRef] [PubMed]
- Harzing, A.W. Publish or Perish on MacOS 2007. Available online: https://harzing.com/resources/publish-or-perish (accessed on 1 October 2021).
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. BMJ 2009, 339, b2535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pullin, A.S.; Stewart, G.B. Guidelines for Systematic Review in Conservation and Environmental Management. Conserv. Biol. 2006, 20, 1647–1656. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Lin, J.; Demner-Fushman, D. Evaluation of PICO as a Knowledge Representation for Clinical Questions. AMIA Annu. Symp. Proc. 2006, 2006, 359–363. [Google Scholar]
- Rohatgi, A. Plotdigitizer: Version 3 2021. Available online: http://plotdigitizer.sourceforge.net (accessed on 14 December 2021).
- Hedges, L.V. Distribution Theory for Glass’s Estimator of Effect Size and Related Estimators. J. Educ. Stat. 1981, 6, 107–128. [Google Scholar] [CrossRef]
- Buck, R.J.; Fieberg, J.; Larkin, D.J. The Use of Weighted Averages of Hedges’ d in Meta-Analysis: Is It Worth It? Methods Ecol. Evol. 2022, 13, 1093–1105. [Google Scholar] [CrossRef]
- Rosenberg, M.S.; Rothstein, H.R.; Gurevitch, J. Effect Sizes: Conventional Choices and Calculations. In Handbook of Meta-analysis in Ecology and Evolution; Princeton University Press: Princeton, NJ, USA, 2013; pp. 61–71. ISBN 978-0-691-13728-5. [Google Scholar]
- Borenstein, M.; Hedges, L.V.; Higgins, J.P.T.; Rothstein, H.R. A Basic Introduction to Fixed-Effect and Random-Effects Models for Meta-Analysis. Res. Synth. Methods 2010, 1, 97–111. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Viechtbauer, W. Conducting Meta-Analyses in R with the Metafor Package. J. Stat. Softw. 2010, 36, 1–48. [Google Scholar] [CrossRef] [Green Version]
- Salerno, M.; Berlino, M.; Mangano, M.C.; Sarà, G. Microplastics and the Functional Traits of Fishes: A Global Meta-analysis. Glob. Change Biol. 2021, 27, 2645–2655. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M. Lme4: Linear Mixed-Effects Models Using S4 Classes. CRAN. 2019. Available online: https://cran.r-project.org/package=lme4 (accessed on 25 August 2022).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Statis-Tical Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Egger, M.; Smith, G.D.; Schneider, M.; Minder, C. Bias in Meta-Analysis Detected by a Simple, Graphical Test. BMJ 1997, 315, 629–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duval, S. The Trim and Fill Method. In Publication Bias in Meta-Analysis: Prevention, Assessment and Adjustments; Rothstein, H.R., Sutton, A.J., Borenstein, M., Eds.; John Wiley & Sons, Ltd: West Sussex, UK, 2005; pp. 127–144. ISBN 978-0-470-87016-7. [Google Scholar]
- Duval, S.; Tweedie, R. Trim and Fill: A Simple Funnel-Plot-Based Method of Testing and Adjusting for Publication Bias in Meta-Analysis. Biometrics 2000, 56, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Marks-Anglin, A.; Duan, R.; Lin, L.; Hong, C.; Chu, H.; Chen, Y. Accounting for Publication Bias Using a Bivariate Trim and Fill Meta-Analysis Procedure. Stat. Med. 2022, 41, 3466–3478. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Castilla, B.; Declercq, L.; Jamshidi, L.; Beretvas, S.N.; Onghena, P.; Van den Noortgate, W. Detecting Selection Bias in Meta-Analyses with Multiple Outcomes: A Simulation Study. J. Exp. Educ. 2021, 89, 125–144. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth: A New Global Map of Terrestrial Ecoregions Provides an Innovative Tool for Conserving Biodiversity. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- dos Anjos, A.G.; Solé, M.; Benchimol, M. Fire Effects on Anurans: What We Know so Far? For. Ecol. Manag. 2021, 495, 119338. [Google Scholar] [CrossRef]
- Cairney, J.W.G.; Bastias, B.A. Influences of Fire on Forest Soil Fungal Communities This Article Is One of a Selection of Papers Published in the Special Forum on Towards Sustainable Forestry—The Living Soil: Soil Biodiversity and Ecosystem Function. Can. J. For. Res. 2007, 37, 207–215. [Google Scholar] [CrossRef]
- Grady, K.C.; Hart, S.C. Influences of Thinning, Prescribed Burning, and Wildfire on Soil Processes and Properties in Southwestern Ponderosa Pine Forests: A Retrospective Study. For. Ecol. Manag. 2006, 234, 123–135. [Google Scholar] [CrossRef]
- Waldrop, M.P.; Harden, J.W. Interactive Effects of Wildfire and Permafrost on Microbial Communities and Soil Processes in an Alaskan Black Spruce Forest. Glob. Change Biol. 2008, 14, 2591–2602. [Google Scholar] [CrossRef]
- Whitman, T.; Whitman, E.; Woolet, J.; Flannigan, M.D.; Thompson, D.K.; Parisien, M.-A. Soil Bacterial and Fungal Response to Wildfires in the Canadian Boreal Forest across a Burn Severity Gradient. Soil Biol. Biochem. 2019, 138, 107571. [Google Scholar] [CrossRef]
- Day, N.J.; Cumming, S.G.; Dunfield, K.E.; Johnstone, J.F.; Mack, M.C.; Reid, K.A.; Turetsky, M.R.; Walker, X.J.; Baltzer, J.L. Identifying Functional Impacts of Heat-Resistant Fungi on Boreal Forest Recovery After Wildfire. Front. For. Glob. Change 2020, 3, 68. [Google Scholar] [CrossRef]
- Neary, D.G.; Klopatek, C.C.; DeBano, L.F.; Ffolliott, P.F. Fire Effects on Belowground Sustainability: A Review and Synthesis. For. Ecol. Manag. 1999, 122, 51–71. [Google Scholar] [CrossRef]
- Smith, G.R.; Edy, L.C.; Peay, K.G. Contrasting Fungal Responses to Wildfire across Different Ecosystem Types. Mol. Ecol. 2021, 30, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Pulido-Chavez, M.F.; Alvarado, E.C.; DeLuca, T.H.; Edmonds, R.L.; Glassman, S.I. High-Severity Wildfire Reduces Richness and Alters Composition of Ectomycorrhizal Fungi in Low-Severity Adapted Ponderosa Pine Forests. For. Ecol. Manag. 2021, 485, 118923. [Google Scholar] [CrossRef]
- Wang, Q.; Zhong, M.; Wang, S. A Meta-Analysis on the Response of Microbial Biomass, Dissolved Organic Matter, Respiration, and N Mineralization in Mineral Soil to Fire in Forest Ecosystems. For. Ecol. Manag. 2012, 271, 91–97. [Google Scholar] [CrossRef]
- Singh, D.; Sharma, P.; Kumar, U.; Daverey, A.; Arunachalam, K. Effect of Forest Fire on Soil Microbial Biomass and Enzymatic Activity in Oak and Pine Forests of Uttarakhand Himalaya, India. Ecol. Process. 2021, 10, 29. [Google Scholar] [CrossRef]
- Mabuhay, J.A.; Nakagoshi, N.; Isagi, Y. Soil Microbial Biomass, Abundance, and Diversity in a Japanese Red Pine Forest: First Year after Fire. J. For. Res. 2006, 11, 165–173. [Google Scholar] [CrossRef]
- Pechenik, J. Biology of the Invertebrates, 7th ed.; McGraw Hill: New York, NY, USA, 2014; ISBN 978-0-07-352418-4. [Google Scholar]
- Santos, X.; Mateos, E.; Bros, V.; Brotons, L.; De Mas, E.; Herraiz, J.A.; Herrando, S.; Miño, À.; Olmo-Vidal, J.M.; Quesada, J.; et al. Is Response to Fire Influenced by Dietary Specialization and Mobility? A Comparative Study with Multiple Animal Assemblages. PLoS ONE 2014, 9, e88224. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, A.D.; Brook, B.W.; Griffiths, A.D.; Brook, B.W. Effect of Fire on Small Mammals: A Systematic Review. Int. J. Wildland Fire 2014, 23, 1034–1043. [Google Scholar] [CrossRef] [Green Version]
- Renken, R.B. Does Fire Affect Amphibians and Reptiles in Eastern U.S. Oak Forests? In Fire in Eastern Oak Forests: Delivering Science to Land Managers. U.S. Forest Service General Technical Report NRS-P-1; Dickinson, M.B., Ed.; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newton Square, PA, USA, 2006; pp. 158–166. [Google Scholar]
- Hoffmann, B.D.; Andersen, A.N. Responses of Ants to Disturbance in Australia, with Particular Reference to Functional Groups. Austral Ecol. 2003, 28, 444–464. [Google Scholar] [CrossRef]
- Gongalsky, K.B.; Cividanes, F.J. Distribution of Carabid Beetles in Agroecosystems across Spatial Scales—A Review. Balt. J. Coleopterol. 2008, 8, 15–30. [Google Scholar]
- Gongalsky, K.B.; Zaitsev, A.S.; Korobushkin, D.I.; Saifutdinov, R.A.; Butenko, K.O.; de Vries, F.T.; Ekschmitt, K.; Degtyarev, M.I.; Gorbunova, A.Yu.; Kostina, N.V.; et al. Forest Fire Induces Short-Term Shifts in Soil Food Webs with Consequences for Carbon Cycling. Ecol. Lett. 2021, 24, 438–450. [Google Scholar] [CrossRef] [PubMed]
- Floyd, T.M.; Russell, K.R.; Moorman, C.E.; Van Lear, D.H.; Guynn, D.C.J.; Lanham, J.D. Effects of Prescribed Fire on Herpetofauna within Hardwood Forests of the Upper Piedmont of South Carolina: A Preliminary Analysis. In Proceedings of the eleventh biennial southern silvicultural research conference. Gen. Tech. Rep. SRS-48; Outcalt, K.W., Ed.; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2002; pp. 123–127. [Google Scholar]
- Greenberg, C.H.; Waldrop, T.A. Short-Term Response of Reptiles and Amphibians to Prescribed Fire and Mechanical Fuel Reduction in a Southern Appalachian Upland Hardwood Forest. For. Ecol. Manag. 2008, 255, 2883–2893. [Google Scholar] [CrossRef]
- Greenberg, C.H. Fire, Habitat Structure and Herpetofauna in the Southeast. In Proceedings of a Special Workshop on the Role of Fire for Nongame Wildlife Management and Community Restoration: Traditional Uses and New Directions; Ford, W.M., Ford, K.R., Moorman, C.E., Eds.; USDA Forest Service General Technical Report NE-288; Northern Research Station: Newtown Square, PA, USA, 2000; pp. 91–99. [Google Scholar]
- Hankins, D.L. The Effects of Indigenous Prescribed Fire on Herpetofauna and Small Mammals in Two Central Valley California Riparian Ecosystems. Calif. Geogr. 2009, 49, 31–50. [Google Scholar]
- Gaines, W.L.; Lyons, A.L.; Weaver, K.; Sprague, A. Monitoring the Short-Term Effects of Prescribed Fire on an Endemic Mollusk in the Dry Forests of the Eastern Cascades, Washington, USA. For. Ecol. Manag. 2011, 261, 1460–1465. [Google Scholar] [CrossRef]
- Ray, E.J.; Bergey, E.A. After the Burn: Factors Affecting Land Snail Survival in Post-Prescribed-Burn Woodlands. J. Molluscan Stud. 2015, 81, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Nowacki, G.J.; Abrams, M.D. The Demise of Fire and “Mesophication” of Forests in the Eastern United States. BioScience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Adams, D.E.; Anderson, R.C.; Collins, S.L. Differential Response of Woody and Herbaceous Species to Summer and Winter Burning in an Oklahoma Grassland. Southwest. Nat. 1982, 27, 55–61. [Google Scholar] [CrossRef]
- Holmes, P. A Comparison of the Impacts of Winter versus Summer Burning of Slash Fuel in Alien-Invaded Fynbos Areas in the Western Cape. South. Afr. For. J. 2001, 192, 41–50. [Google Scholar] [CrossRef]
- Gabrey, S.W.; Afton, A.D.; Wilson, B.C. Effects of Structural Marsh Management and Winter Burning on Plant and Bird Communities during Summer in the Gulf Coast Chenier Plain. Wildl. Soc. Bull. 2001, 29, 218–231. [Google Scholar]
- Bond, W.J.; Keeley, J.E. Fire as a Global ‘Herbivore’: The Ecology and Evolution of Flammable Ecosystems. Trends Ecol. Evol. 2005, 20, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, R.E.; Bent, E.; Hollingsworth, T.N.; Chapin, F.S.; Taylor, D.L. Resilience of Arctic Mycorrhizal Fungal Communities after Wildfire Facilitated by Resprouting Shrubs. Écoscience 2013, 20, 296–310. [Google Scholar] [CrossRef]
- Heim, R.J.; Heim, W.; Bültmann, H.; Kamp, J.; Rieker, D.; Yurtaev, A.; Hölzel, N. Fire Disturbance Promotes Biodiversity of Plants, Lichens and Birds in the Siberian Subarctic Tundra. Glob. Change Biol. 2022, 28, 1048–1062. [Google Scholar] [CrossRef] [PubMed]
- Ramo, R.; Roteta, E.; Bistinas, I.; van Wees, D.; Bastarrika, A.; Chuvieco, E.; van der Werf, G.R. African Burned Area and Fire Carbon Emissions Are Strongly Impacted by Small Fires Undetected by Coarse Resolution Satellite Data. Proc. Natl. Acad. Sci. USA 2021, 118, e2011160118. [Google Scholar] [CrossRef]
- Tosca, M. The Impact of Savanna Fires on Africa’s Rainfall Patterns. Available online: http://theconversation.com/the-impact-of-savanna-fires-on-africas-rainfall-patterns-46942 (accessed on 24 August 2022).
- Probert, J.R.; Parr, C.L.; Holdo, R.M.; Anderson, T.M.; Archibald, S.; Courtney Mustaphi, C.J.; Dobson, A.P.; Donaldson, J.E.; Hopcraft, G.C.; Hempson, G.P.; et al. Anthropogenic Modifications to Fire Regimes in the Wider Serengeti-Mara Ecosystem. Glob. Change Biol. 2019, 25, 3406–3423. [Google Scholar] [CrossRef]
- Regos, A.; Tapia, L.; Arenas-Castro, S.; Gil-Carrera, A.; Domínguez, J. Ecosystem Functioning Influences Species Fitness at Upper Trophic Levels. Ecosystems 2022, 25, 1037–1051. [Google Scholar] [CrossRef]
- Calizza, E.; Costantini, M.L.; Rossi, L. Effect of Multiple Disturbances on Food Web Vulnerability to Biodiversity Loss in Detritus-Based Systems. Ecosphere 2015, 6, art124. [Google Scholar] [CrossRef] [Green Version]
- Vallejo, V.R.; Arianoutsou, M.; Moreira, F. Fire Ecology and Post-Fire Restoration Approaches in Southern European Forest Types. In Post-Fire Management and Restoration of Southern European Forests; Moreira, F., Arianoutsou, M., Corona, P., De las Heras, J., Eds.; Managing Forest Ecosystems; Springer: Dordrecht, The Netherlands, 2012; pp. 93–119. ISBN 978-94-007-2208-8. [Google Scholar]
- de las Heras, J.; Moya, D.; Vega, J.A.; Daskalakou, E.; Vallejo, V.R.; Grigoriadis, N.; Tsitsoni, T.; Baeza, J.; Valdecantos, A.; Fernández, C.; et al. Post-Fire Management of Serotinous Pine Forests. In Post-Fire Management and Restoration of Southern European Forests; Moreira, F., Arianoutsou, M., Corona, P., De las Heras, J., Eds.; Managing Forest Ecosystems; Springer: Dordrecht, The Netherlands, 2012; pp. 121–150. ISBN 978-94-007-2208-8. [Google Scholar]
- Glassman, S.I.; Levine, C.R.; DiRocco, A.M.; Battles, J.J.; Bruns, T.D. Ectomycorrhizal Fungal Spore Bank Recovery after a Severe Forest Fire: Some like It Hot. ISME J. 2016, 10, 1228–1239. [Google Scholar] [CrossRef]
- Burkle, L.A.; Simanonok, M.P.; Durney, J.S.; Myers, J.A.; Belote, R.T. Wildfires Influence Abundance, Diversity, and Intraspecific and Interspecific Trait Variation of Native Bees and Flowering Plants Across Burned and Unburned Landscapes. Front. Ecol. Evol. 2019, 7, 252. [Google Scholar] [CrossRef] [Green Version]
- Moritz, M.A.; Parisien, M.-A.; Batllori, E.; Krawchuk, M.A.; Van Dorn, J.; Ganz, D.J.; Hayhoe, K. Climate Change and Disruptions to Global Fire Activity. Ecosphere 2012, 3, art49. [Google Scholar] [CrossRef]
- Hong, J.; Zhang, J.; Ma, Y.; Gu, B.; Lee, R. The Fates of Nitrogen in an Experimental Wetland Food Web: A Stable Isotope Study. Wetlands 2019, 39, 303–310. [Google Scholar] [CrossRef]
- Chasar, L.C.; Chanton, J.P.; Koenig, C.C.; Coleman, F.C. Evaluating the Effect of Environmental Disturbance on the Trophic Structure of Florida Bay, U.S.A.: Multiple Stable Isotope Analyses of Contemporary and Historical Specimens. Limnol. Oceanogr. 2005, 50, 1059–1072. [Google Scholar] [CrossRef]
- Blonder, B.; Lamanna, C.; Violle, C.; Enquist, B.J. The N-Dimensional Hypervolume. Glob. Ecol. Biogeogr. 2014, 23, 595–609. [Google Scholar] [CrossRef]
- Blonder, B.; Morrow, C.B.; Maitner, B.; Harris, D.J.; Lamanna, C.; Violle, C.; Enquist, B.J.; Kerkhoff, A.J. New Approaches for Delineating N-Dimensional Hypervolumes. Methods Ecol. Evol. 2018, 9, 305–319. [Google Scholar] [CrossRef] [Green Version]
- Whyte, K.P. On the Role of Traditional Ecological Knowledge as a Collaborative Concept: A Philosophical Study. Ecol. Process. 2013, 2, 7. [Google Scholar] [CrossRef]
- Ganey, J.L.; Iniguez, J.M.; Sanderlin, J.S.; Block, W.M. Developing a Monitoring Program for Bird Populations in the Chiricahua Mountains, Arizona, Using Citizen Observers: Initial Stages; General Technical Report (GTR) RMRS-GTR-368; U.S, Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2017. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Expectations | |
|---|---|
| Question 1: to what extent does fire disturbance affect aboveground and belowground organisms? Hypothesis A: aboveground organisms will be negatively affected to a greater extent than belowground organism as the latter tend to have the ability to move and to other habitats compared to their underground counterparts |
|
| Question 2: how do contrasting fire types of intensities affect organisms? |
|
| Question 3: how do fire effects vary across biomes and continental scales? |
|
| Questions 4: how do key abiotic factors such as moisture, pH, and sampling depth influence below-ground organism responses to fire? Hypothesis B: Abiotic factors should have differential effects on organisms |
|
| General Response Variable | Specific Response Variable |
|---|---|
| Abundance | representation of number of organisms Number of Individual per unit area/volume (e.g., ind.m−2, individual.core−1, individual. gram of soil−1, nematodes. cm−3) Colony Forming Units (CFU) Spores per unit of soil |
| Biomass | total mass of organisms in an area, habitat, or region dry weight (mg.g−1, ug ergosterol/g, PLFA/Kg, ug/g dry soil, mg) |
| Richness | number of species in a community (number of species or genera, families, OTUs, morphospecies, groups) |
| Diversity | Shannon-Wiener diversity index Simpson Diversity Index McIntosh index |
| Evenness | Pielou’s evenness, Simpson’s equitability, Shannon evenness |
| Response | Independent Variable | β | SE | Lower-95 | Upper-95 | R2 | p |
|---|---|---|---|---|---|---|---|
| Abundance | |||||||
| Moisture | 322.00 | 276.73 | −187.04 | 833.74 | 0.03 | 0.26 | |
| pH | 1846.00 | 3360.23 | −4341.37 | 8842.58 | 0.03 | 0.58 | |
| Depth | 1717.50 | 17,212.01 | −1437.74 | 4868.22 | 0.03 | 0.34 | |
| Biomass | |||||||
| Moisture | 0.05 | 0.03 | 0.00 | 0.11 | 0.12 | 0.06 | |
| pH | −0.20 | 0.22 | −0.67 | 0.21 | 0.12 | 0.36 | |
| Depth | 0.18 | 0.09 | 0.00 | 0.36 | 0.12 | 0.05 | |
| Diversity | |||||||
| Moisture | 0.00 | 0.04 | −0.06 | 0.06 | 0.13 | 0.95 | |
| pH | 0.48 | 0.49 | −0.37 | 1.03 | 0.13 | 0.41 | |
| Depth | −0.16 | 0.22 | −0.39 | 0.01 | 0.13 | 0.62 | |
| Evenness | |||||||
| Moisture | −0.08 | 0.02 | −0.11 | −0.05 | 0.72 | 0.03 | |
| pH | −0.51 | 0.23 | −0.85 | −0.16 | 0.72 | 0.12 | |
| Depth | 0.63 | 0.36 | 0.10 | 1.15 | 0.72 | 1.00 | |
| Richness | |||||||
| Moisture | −0.03 | 0.06 | −0.15 | 0.07 | 0.26 | 0.62 | |
| pH | 0.63 | 0.80 | −1.52 | 1.88 | 0.26 | 0.44 | |
| Depth | −0.27 | 0.16 | −0.52 | −0.02 | 0.26 | 0.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moyo, S. Community Responses to Fire: A Global Meta-Analysis Unravels the Contrasting Responses of Fauna to Fire. Earth 2022, 3, 1087-1111. https://doi.org/10.3390/earth3040063
Moyo S. Community Responses to Fire: A Global Meta-Analysis Unravels the Contrasting Responses of Fauna to Fire. Earth. 2022; 3(4):1087-1111. https://doi.org/10.3390/earth3040063
Chicago/Turabian StyleMoyo, Sydney. 2022. "Community Responses to Fire: A Global Meta-Analysis Unravels the Contrasting Responses of Fauna to Fire" Earth 3, no. 4: 1087-1111. https://doi.org/10.3390/earth3040063
APA StyleMoyo, S. (2022). Community Responses to Fire: A Global Meta-Analysis Unravels the Contrasting Responses of Fauna to Fire. Earth, 3(4), 1087-1111. https://doi.org/10.3390/earth3040063