
Uptake and Advanced Therapy of Butyrate in Inflammatory Bowel Disease

Abstract
:1. Introduction
2. SCFAs and Intestinal Homeostasis in IBD
2.1. The Role of SCFAs in Intestinal Immune Response
2.2. SCFAs in IBD
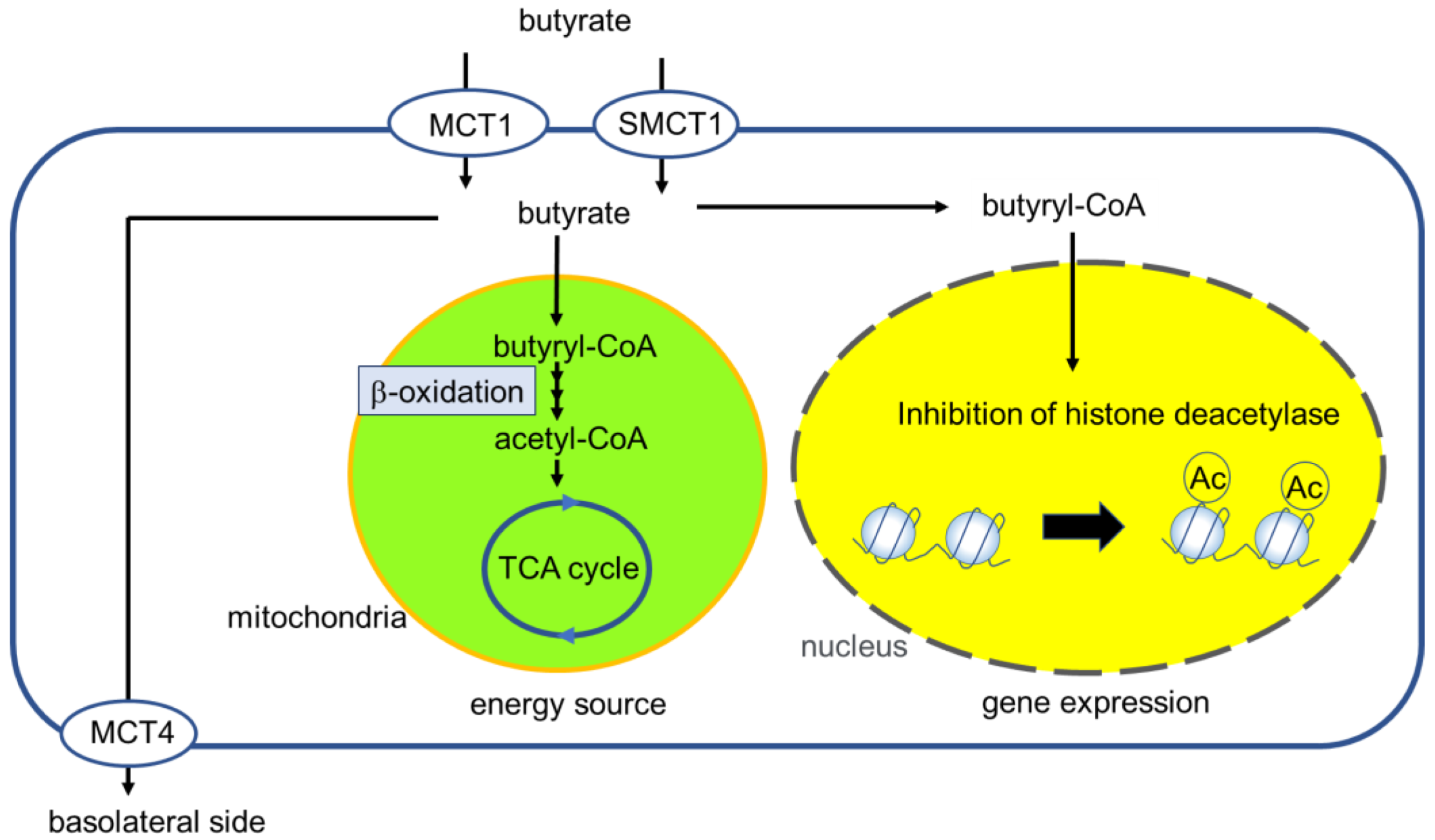
3. Butyrate Transport to IECs
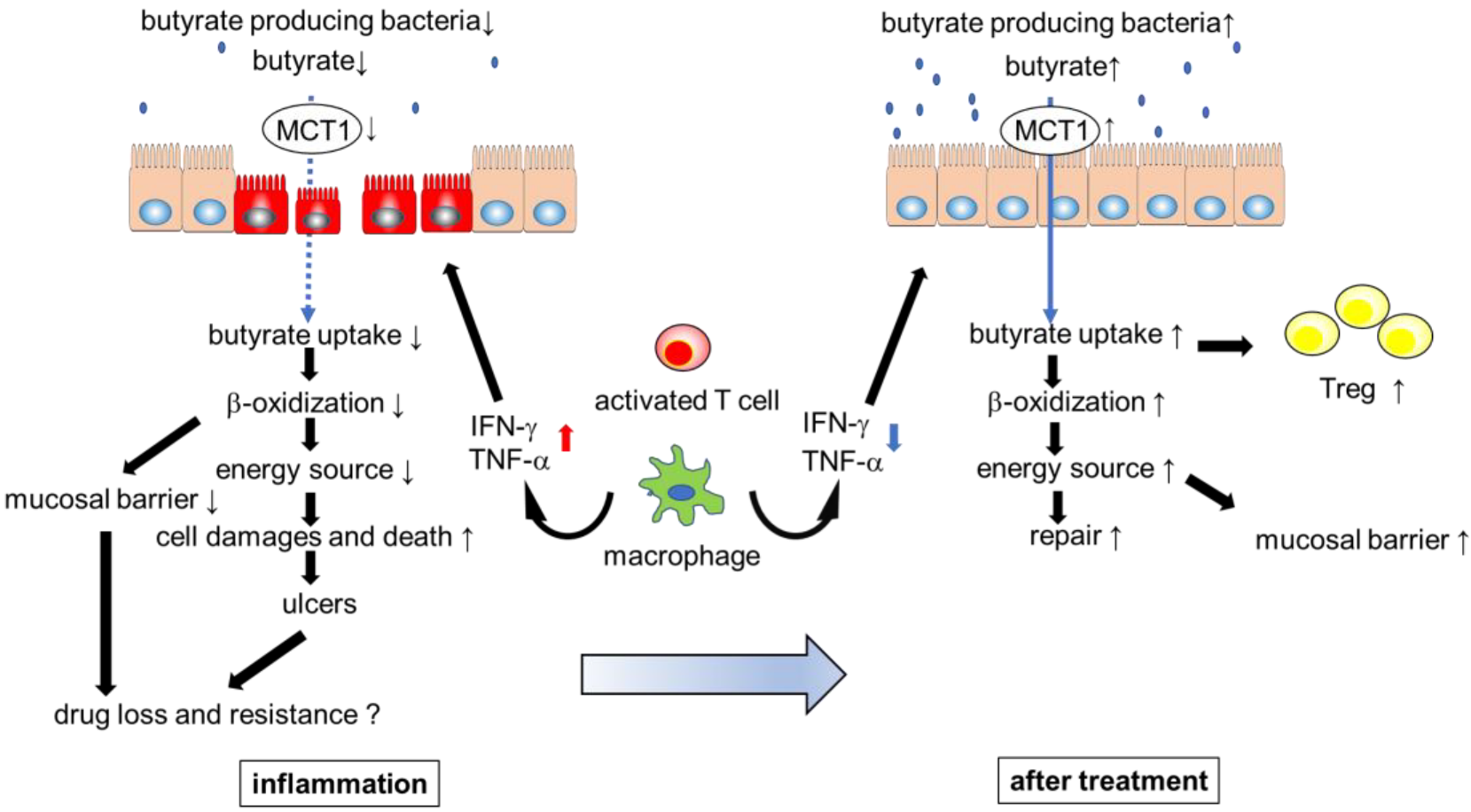
4. Regulation and Role of MCT1 in IECs
5. Gut Microbiota and Systemic Immune Response
6. Influence of Gut Microbiota on Response to Advanced Therapies
7. Advanced Therapy with Butyrate
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Molodecky, N.A.; Soon, I.S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W.; et al. Increasing Incidence and Prevalence of the Inflammatory Bowel Diseases with Time, Based on Systematic Review. Gastroenterology 2012, 142, 46–54.e42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, W.K.; Wong, S.H.; Ng, S.C. Changing Epidemiological Trends of Inflammatory Bowel Disease in Asia. Intest. Res. 2016, 14, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ananthakrishnan, A.N. Epidemiology and Risk Factors for IBD. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Glassner, K.L.; Abraham, B.P.; Quigley, E.M.M. The Microbiome and Inflammatory Bowel Disease. J. Allergy Clin. Immunol. 2020, 145, 16–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchesi, J.R.; Holmes, E.; Khan, F.; Kochhar, S.; Scanlan, P.; Shanahan, F.; Wilson, I.D.; Wang, Y. Rapid and Noninvasive Metabonomic Characterization of Inflammatory Bowel Disease. J. Proteome Res. 2007, 6, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Imhann, F.; Vich Vila, A.; Bonder, M.J.; Fu, J.; Gevers, D.; Visschedijk, M.C.; Spekhorst, L.M.; Alberts, R.; Franke, L.; van Dullemen, H.M.; et al. Interplay of Host Genetics and Gut Microbiota Underlying the Onset and Clinical Presentation of Inflammatory Bowel Disease. Gut 2018, 67, 108–119. [Google Scholar] [CrossRef]
- Ananthakrishnan, A.N.; Luo, C.; Yajnik, V.; Khalili, H.; Garber, J.J.; Stevens, B.W.; Cleland, T.; Xavier, R.J. Gut Microbiome Function Predicts Response to Anti-Integrin Biologic Therapy in Inflammatory Bowel Diseases. Cell Host Microbe 2017, 21, 603–610.e3. [Google Scholar] [CrossRef] [Green Version]
- Effenberger, M.; Reider, S.; Waschina, S.; Bronowski, C.; Enrich, B.; Adolph, T.E.; Koch, R.; Moschen, A.R.; Rosenstiel, P.; Aden, K.; et al. Microbial Butyrate Synthesis Indicates Therapeutic Efficacy of Azathioprine in IBD Patients. J. Crohns. Colitis. 2021, 15, 88–98. [Google Scholar] [CrossRef]
- Aden, K.; Rehman, A.; Waschina, S.; Pan, W.-H.; Walker, A.; Lucio, M.; Nunez, A.M.; Bharti, R.; Zimmerman, J.; Bethge, J.; et al. Metabolic Functions of Gut Microbes Associate with Efficacy of Tumor Necrosis Factor Antagonists in Patients with Inflammatory Bowel Diseases. Gastroenterology 2019, 157, 1279–1292.e11. [Google Scholar] [CrossRef] [Green Version]
- Magnusson, M.K.; Strid, H.; Sapnara, M.; Lasson, A.; Bajor, A.; Ung, K.-A.; Öhman, L. Anti-TNF Therapy Response in Patients with Ulcerative Colitis Is Associated with Colonic Antimicrobial Peptide Expression and Microbiota Composition. ECCOJC 2016, 10, 943–952. [Google Scholar] [CrossRef]
- Sivaprakasam, S.; Bhutia, Y.D.; Yang, S.; Ganapathy, V. Short-Chain Fatty Acid Transporters: Role in Colonic Homeostasis. Compr. Physiol. 2017, 8, 299–314. [Google Scholar] [CrossRef]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The Microbial Metabolite Butyrate Regulates Intestinal Macrophage Function via Histone Deacetylase Inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of Colonic Regulatory T Cells by Indigenous Clostridium Species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal Microbe-Derived Butyrate Induces the Differentiation of Colonic Regulatory T Cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Yang, W.; Yu, T.; Huang, X.; Bilotta, A.J.; Xu, L.; Lu, Y.; Sun, J.; Pan, F.; Zhou, J.; Zhang, W.; et al. Intestinal Microbiota-Derived Short-Chain Fatty Acids Regulation of Immune Cell IL-22 Production and Gut Immunity. Nat. Commun. 2020, 11, 4457. [Google Scholar] [CrossRef]
- Wang, R.X.; Lee, J.S.; Campbell, E.L.; Colgan, S.P. Microbiota-Derived Butyrate Dynamically Regulates Intestinal Homeostasis through Regulation of Actin-Associated Protein Synaptopodin. Proc. Natl. Acad. Sci. USA 2020, 117, 11648–11657. [Google Scholar] [CrossRef]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef] [Green Version]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. GPR109A Is a G-Protein–Coupled Receptor for the Bacterial Fermentation Product Butyrate and Functions as a Tumor Suppressor in Colon. Cancer Res. 2009, 69, 2826–2832. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Lin, J.; Zhang, C.; Gao, H.; Lu, H.; Gao, X.; Zhu, R.; Li, Z.; Li, M.; Liu, Z. Microbiota Metabolite Butyrate Constrains Neutrophil Functions and Ameliorates Mucosal Inflammation in Inflammatory Bowel Disease. Gut Microbes 2021, 13, 1968257. [Google Scholar] [CrossRef]
- Rizzello, F.; Spisni, E.; Giovanardi, E.; Imbesi, V.; Salice, M.; Alvisi, P.; Valerii, M.C.; Gionchetti, P. Implications of the Westernized Diet in the Onset and Progression of IBD. Nutrients 2019, 11, 1033. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.K.; Abraham, B.; El-Serag, H. Dietary Intake and Risk of Developing Inflammatory Bowel Disease: A Systematic Review of the Literature. Am. J. Gastroenterol. 2011, 106, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Dolan, K.T.; Chang, E.B. Diet, Gut Microbes, and the Pathogenesis of Inflammatory Bowel Diseases. Mol. Nutr. Food Res. 2017, 61, 1600129. [Google Scholar] [CrossRef] [Green Version]
- Kostic, A.D.; Xavier, R.J.; Gevers, D. The Microbiome in Inflammatory Bowel Disease: Current Status and the Future Ahead. Gastroenterology 2014, 146, 1489–1499. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Flint, H.J. Formation of Propionate and Butyrate by the Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Celiberto, L.S.; Graef, F.A.; Healey, G.R.; Bosman, E.S.; Jacobson, K.; Sly, L.M.; Vallance, B.A. Inflammatory Bowel Disease and Immunonutrition: Novel Therapeutic Approaches through Modulation of Diet and the Gut Microbiome. Immunology 2018, 155, 36–52. [Google Scholar] [CrossRef]
- Litvak, Y.; Byndloss, M.X.; Tsolis, R.M.; Bäumler, A.J. Dysbiotic Proteobacteria Expansion: A Microbial Signature of Epithelial Dysfunction. Curr. Opin. Microbiol. 2017, 39, 1–6. [Google Scholar] [CrossRef]
- Gasaly, N.; Hermoso, M.A.; Gotteland, M. Butyrate and the Fine-Tuning of Colonic Homeostasis: Implication for Inflammatory Bowel Diseases. IJMS 2021, 22, 3061. [Google Scholar] [CrossRef]
- Gonçalves, P.; Araújo, J.R.; Pinho, M.J.; Martel, F. Modulation of Butyrate Transport in Caco-2 Cells. Naunyn-Schmied. Arch. Pharm. 2009, 379, 325–336. [Google Scholar] [CrossRef]
- Gonçalves, P.; Martel, F. Regulation of Colonic Epithelial Butyrate Transport: Focus on Colorectal Cancer. Porto. Biomed. J. 2016, 1, 83–91. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short Chain Fatty Acids in Human Large Intestine, Portal, Hepatic and Venous Blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjiagapiou, C.; Schmidt, L.; Dudeja, P.K.; Layden, T.J.; Ramaswamy, K. Mechanism(s) of Butyrate Transport in Caco-2 Cells: Role of Monocarboxylate Transporter 1. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G775–G780. [Google Scholar] [CrossRef] [PubMed]
- Saksena, S.; Theegala, S.; Bansal, N.; Gill, R.K.; Tyagi, S.; Alrefai, W.A.; Ramaswamy, K.; Dudeja, P.K. Mechanisms Underlying Modulation of Monocarboxylate Transporter 1 (MCT1) by Somatostatin in Human Intestinal Epithelial Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G878–G885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blachier, F.; de Sá Resende, A.; da Silva Fogaça Leite, G.; Vasques da Costa, A.; Lancha Junior, A.H. Colon Epithelial Cells Luminal Environment and Physiopathological Consequences: Impact of Nutrition and Exercise. Nutrire 2018, 43, 2. [Google Scholar] [CrossRef]
- Donohoe, D.R.; Garge, N.; Zhang, X.; Sun, W.; O’Connell, T.M.; Bunger, M.K.; Bultman, S.J. The Microbiome and Butyrate Regulate Energy Metabolism and Autophagy in the Mammalian Colon. Cell Metab. 2011, 13, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Gill, R.K.; Saksena, S.; Alrefai, W.A.; Sarwar, Z.; Goldstein, J.L.; Carroll, R.E.; Ramaswamy, K.; Dudeja, P.K. Expression and Membrane Localization of MCT Isoforms along the Length of the Human Intestine. Am. J. Physiol. Cell Physiol. 2005, 289, C846–C852. [Google Scholar] [CrossRef] [Green Version]
- Bonomo, R.R.; Cook, T.M.; Gavini, C.K.; White, C.R.; Jones, J.R.; Bovo, E.; Zima, A.V.; Brown, I.A.; Dugas, L.R.; Zakharian, E.; et al. Fecal Transplantation and Butyrate Improve Neuropathic Pain, Modify Immune Cell Profile, and Gene Expression in the PNS of Obese Mice. Proc. Natl. Acad. Sci. USA 2020, 117, 26482–26493. [Google Scholar] [CrossRef]
- Bloemen, J.G.; Venema, K.; van de Poll, M.C.; Olde Damink, S.W.; Buurman, W.A.; Dejong, C.H. Short Chain Fatty Acids Exchange across the Gut and Liver in Humans Measured at Surgery. Clin. Nutr. 2009, 28, 657–661. [Google Scholar] [CrossRef]
- Cuff, M.A.; Lambert, D.W.; Shirazi-beechey, S.P. Substrate-Induced Regulation of the Human Colonic Monocarboxylate Transporter, MCT1. J. Physiol. 2002, 539, 361–371. [Google Scholar] [CrossRef]
- Borthakur, A.; Saksena, S.; Gill, R.K.; Alrefai, W.A.; Ramaswamy, K.; Dudeja, P.K. Regulation of Monocarboxylate Transporter 1 (MCT1) Promoter by Butyrate in Human Intestinal Epithelial Cells: Involvement of NF-κB Pathway. J. Cell Biochem. 2008, 103, 1452–1463. [Google Scholar] [CrossRef]
- Kirat, D.; Kondo, K.; Shimada, R.; Kato, S. Dietary Pectin Up-Regulates Monocaboxylate Transporter 1 in the Rat Gastrointestinal Tract: Pectin Up-Regulates MCT1 in Rat Gastrointestinal Tract. Exp. Physiol. 2009, 94, 422–433. [Google Scholar] [CrossRef]
- Haenen, D.; Zhang, J.; Souza da Silva, C.; Bosch, G.; van der Meer, I.M.; van Arkel, J.; van den Borne, J.J.G.C.; Pérez Gutiérrez, O.; Smidt, H.; Kemp, B.; et al. A Diet High in Resistant Starch Modulates Microbiota Composition, SCFA Concentrations, and Gene Expression in Pig Intestine. J. Nutr. 2013, 143, 274–283. [Google Scholar] [CrossRef] [Green Version]
- Borthakur, A.; Priyamvada, S.; Kumar, A.; Natarajan, A.A.; Gill, R.K.; Alrefai, W.A.; Dudeja, P.K. A Novel Nutrient Sensing Mechanism Underlies Substrate-Induced Regulation of Monocarboxylate Transporter-1. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1126–G1133. [Google Scholar] [CrossRef] [Green Version]
- Suk, H.Y.; Zhou, C.; Yang, T.T.C.; Zhu, H.; Yu, R.Y.L.; Olabisi, O.; Yang, X.; Brancho, D.; Kim, J.-Y.; Scherer, P.E.; et al. Ablation of Calcineurin Aβ Reveals Hyperlipidemia and Signaling Cross-Talks with Phosphodiesterases. J. Biol. Chem. 2013, 288, 3477–3488. [Google Scholar] [CrossRef] [Green Version]
- Buyse, M.; Sitaraman, S.V.; Liu, X.; Bado, A.; Merlin, D. Luminal Leptin Enhances CD147/MCT-1-Mediated Uptake of Butyrate in the Human Intestinal Cell Line Caco2-BBE. J. Biol. Chem. 2002, 277, 28182–28190. [Google Scholar] [CrossRef] [Green Version]
- Thibault, R.; De Coppet, P.; Daly, K.; Bourreille, A.; Cuff, M.; Bonnet, C.; Mosnier, J.; Galmiche, J.; Shirazi–Beechey, S.; Segain, J. Down-Regulation of the Monocarboxylate Transporter 1 Is Involved in Butyrate Deficiency during Intestinal Inflammation. Gastroenterology 2007, 133, 1916–1927. [Google Scholar] [CrossRef]
- Villodre Tudela, C.; Boudry, C.; Stumpff, F.; Aschenbach, J.R.; Vahjen, W.; Zentek, J.; Pieper, R. Down-Regulation of Monocarboxylate Transporter 1 (MCT1) Gene Expression in the Colon of Piglets Is Linked to Bacterial Protein Fermentation and pro-Inflammatory Cytokine-Mediated Signalling. Br. J. Nutr. 2015, 113, 610–617. [Google Scholar] [CrossRef] [Green Version]
- De Preter, V.; Arijs, I.; Windey, K.; Vanhove, W.; Vermeire, S.; Schuit, F.; Rutgeerts, P.; Verbeke, K. Impaired Butyrate Oxidation in Ulcerative Colitis Is Due to Decreased Butyrate Uptake and a Defect in the Oxidation Pathway. Inflamm. Bowel Dis. 2012, 18, 1127–1136. [Google Scholar] [CrossRef]
- Ferrer-Picón, E.; Dotti, I.; Corraliza, A.M.; Mayorgas, A.; Esteller, M.; Perales, J.C.; Ricart, E.; Masamunt, M.C.; Carrasco, A.; Tristán, E.; et al. Intestinal Inflammation Modulates the Epithelial Response to Butyrate in Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2020, 26, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Paramsothy, S.; Nielsen, S.; Kamm, M.A.; Deshpande, N.P.; Faith, J.J.; Clemente, J.C.; Paramsothy, R.; Walsh, A.J.; van den Bogaerde, J.; Samuel, D.; et al. Specific Bacteria and Metabolites Associated with Response to Fecal Microbiota Transplantation in Patients with Ulcerative Colitis. Gastroenterology 2019, 156, 1440–1454.e2. [Google Scholar] [CrossRef]
- Daillère, R.; Vétizou, M.; Waldschmitt, N.; Yamazaki, T.; Isnard, C.; Poirier-Colame, V.; Duong, C.P.M.; Flament, C.; Lepage, P.; Roberti, M.P.; et al. Enterococcus Hirae and Barnesiella Intestinihominis Facilitate Cyclophosphamide-Induced Therapeutic Immunomodulatory Effects. Immunity 2016, 45, 931–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Chu, C.-Q. Gut Microbiota–Medication Interaction in Rheumatic Diseases. Front. Immunol. 2021, 12, 796865. [Google Scholar] [CrossRef] [PubMed]
- Maier, L.; Pruteanu, M.; Kuhn, M.; Zeller, G.; Telzerow, A.; Anderson, E.E.; Brochado, A.R.; Fernandez, K.C.; Dose, H.; Mori, H.; et al. Extensive Impact of Non-Antibiotic Drugs on Human Gut Bacteria. Nature 2018, 555, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Roediger, W.E. Utilization of Nutrients by Isolated Epithelial Cells of the Rat Colon. Gastroenterology 1982, 83, 424–429. [Google Scholar] [CrossRef]
- Thibault, R.; Blachier, F.; Darcy-Vrillon, B.; de Coppet, P.; Bourreille, A.; Segain, J.-P. Butyrate Utilization by the Colonic Mucosa in Inflammatory Bowel Diseases: A Transport Deficiency. Inflamm. Bowel Dis. 2010, 16, 684–695. [Google Scholar] [CrossRef]
- De Preter, V.; Geboes, K.P.; Bulteel, V.; Vandermeulen, G.; Suenaert, P.; Rutgeerts, P.; Verbeke, K. Kinetics of Butyrate Metabolism in the Normal Colon and in Ulcerative Colitis: The Effects of Substrate Concentration and Carnitine on the β-Oxidation Pathway: Kinetics of Butyrate Metabolism in Ulcerative Colitis. Aliment. Pharmacol. Ther. 2011, 34, 526–532. [Google Scholar] [CrossRef]
- Santhanam, S.; Venkatraman, A.; Ramakrishna, B.S. Impairment of Mitochondrial Acetoacetyl CoA Thiolase Activity in the Colonic Mucosa of Patients with Ulcerative Colitis. Gut 2007, 56, 1543–1549. [Google Scholar] [CrossRef] [Green Version]
- Michielan, A.; D’Incà, R. Intestinal Permeability in Inflammatory Bowel Disease: Pathogenesis, Clinical Evaluation, and Therapy of Leaky Gut. Mediat. Inflamm. 2015, 2015, 628157. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Chen, L.; Zhou, R.; Wang, X.; Song, L.; Huang, S.; Wang, G.; Xia, B. Increased Proportions of Bifidobacterium and the Lactobacillus Group and Loss of Butyrate-Producing Bacteria in Inflammatory Bowel Disease. J. Clin. Microbiol. 2014, 52, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Ota, S.; Sakuraba, H.; Hiraga, H.; Yoshida, S.; Satake, M.; Akemoto, Y.; Tanaka, N.; Watanabe, R.; Takato, M.; Murai, Y.; et al. Cyclosporine Protects from Intestinal Epithelial Injury by Modulating Butyrate Uptake via Upregulation of Membrane Monocarboxylate Transporter 1 Levels. Biochem. Biophys. Rep. 2020, 24, 100811. [Google Scholar] [CrossRef]
- Kennedy, G.M.; Min, M.Y.; Fitzgerald, J.F.; Nguyen, M.T.; Schultz, S.L.; Crum, M.T.; Starke, J.A.; Butkus, M.A.; Bowman, D.D.; Labare, M.P. Inactivation of the Bacterial Pathogens Staphylococcus Pseudintermedius and Acinetobacter Baumannii by Butanoic Acid. J. Appl. Microbiol. 2019, 126, 752–763. [Google Scholar] [CrossRef]
- Schauber, J. Expression of the Cathelicidin LL-37 Is Modulated by Short Chain Fatty Acids in Colonocytes: Relevance of Signalling Pathways. Gut 2003, 52, 735–741. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Chen, N.; Wu, Z.; Song, Y.; Zhang, Y.; Wu, N.; Zhang, F.; Ren, X.; Liu, Y. 5-Aminosalicylic Acid Alters the Gut Bacterial Microbiota in Patients with Ulcerative Colitis. Front. Microbiol. 2018, 9, 1274. [Google Scholar] [CrossRef] [PubMed]
- Sanchis-Artero, L.; Martínez-Blanch, J.F.; Manresa-Vera, S.; Cortés-Castell, E.; Valls-Gandia, M.; Iborra, M.; Paredes-Arquiola, J.M.; Boscá-Watts, M.; Huguet, J.M.; Gil-Borrás, R.; et al. Evaluation of Changes in Intestinal Microbiota in Crohn’s Disease Patients after Anti-TNF Alpha Treatment. Sci. Rep. 2021, 11, 10016. [Google Scholar] [CrossRef]
- Sossai, P. Butyric Acid: What Is the Future for This Old Substance? Swiss Med. Wkly 2012, 42, w13596. [Google Scholar] [CrossRef]
- Facchin, S.; Vitulo, N.; Calgaro, M.; Buda, A.; Romualdi, C.; Pohl, D.; Perini, B.; Lorenzon, G.; Marinelli, C.; D’Incà, R.; et al. Microbiota Changes Induced by Microencapsulated Sodium Butyrate in Patients with Inflammatory Bowel Disease. Neurogastroenterol. Motil. 2020, 32, e13914. [Google Scholar] [CrossRef]
- Edelman, M.J.; Bauer, K.; Khanwani, S.; Tait, N.; Trepel, J.; Karp, J.; Nemieboka, N.; Chung, E.-J.; Van Echo, D. Clinical and Pharmacologic Study of Tributyrin: An Oral Butyrate Prodrug. Cancer Chemother. Pharm. 2003, 51, 439–444. [Google Scholar] [CrossRef]
- Gill, P.A.; van Zelm, M.C.; Muir, J.G.; Gibson, P.R. Review Article: Short Chain Fatty Acids as Potential Therapeutic Agents in Human Gastrointestinal and Inflammatory Disorders. Aliment. Pharm. Ther. 2018, 48, 15–34. [Google Scholar] [CrossRef] [Green Version]
- Clarke, J.M.; Topping, D.L.; Christophersen, C.T.; Bird, A.R.; Lange, K.; Saunders, I.; Cobiac, L. Butyrate Esterified to Starch Is Released in the Human Gastrointestinal Tract. Am. J. Clin. Nutr. 2011, 94, 1276–1283. [Google Scholar] [CrossRef] [Green Version]
- Clarke, J.M.; Bird, A.R.; Topping, D.L.; Cobiac, L. Excretion of Starch and Esterified Short-Chain Fatty Acids by Ileostomy Subjects after the Ingestion of Acylated Starches. Am. J. Clin. Nutr. 2007, 86, 1146–1151. [Google Scholar] [CrossRef]
- Nielsen, T.; Canibe, N.; Larsen, F. Butyrylation of Maize and Potato Starches and Characterization of the Products by Nuclear Magnetic Resonance and In Vitro Fermentation. Foods 2018, 7, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Cheng, L.; Li, Z.; Li, C.; Hong, Y.; Gu, Z. Butyrylated Starch Protects Mice from DSS-Induced Colitis: Combined Effects of Butyrate Release and Prebiotic Supply. Food Funct. 2021, 12, 11290–11302. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Kinashi, Y.; Li, J.; Yoshikawa, T.; Kishimura, A.; Tanaka, M.; Matsui, T.; Mori, T.; Hase, K.; Katayama, Y. Polyvinyl Butyrate Nanoparticles as Butyrate Donors for Colitis Treatment. ACS Appl. Bio Mater. 2021, 4, 2335–2341. [Google Scholar] [CrossRef]
- Henn, M.R.; O’Brien, E.J.; Diao, L.; Feagan, B.G.; Sandborn, W.J.; Huttenhower, C.; Wortman, J.R.; McGovern, B.H.; Wang-Weigand, S.; Lichter, D.I.; et al. A Phase 1b Safety Study of SER-287, a Spore-Based Microbiome Therapeutic, for Active Mild to Moderate Ulcerative Colitis. Gastroenterology 2021, 160, 115–127.e30. [Google Scholar] [CrossRef]
- Vernero, M.; De Blasio, F.; Ribaldone, D.G.; Bugianesi, E.; Pellicano, R.; Saracco, G.M.; Astegiano, M.; Caviglia, G.P. The Usefulness of Microencapsulated Sodium Butyrate Add-On Therapy in Maintaining Remission in Patients with Ulcerative Colitis: A Prospective Observational Study. JCM 2020, 9, 3941. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Laeremans, T.; Vanhove, W.; Arnauts, K.; Ramalho, A.S.; Farré, R.; Cleynen, I.; Ferrante, M.; Vermeire, S. Butyrate Does Not Protect Against Inflammation-Induced Loss of Epithelial Barrier Function and Cytokine Production in Primary Cell Monolayers from Patients with Ulcerative Colitis. J. Crohns Colitis 2019, 13, 1351–1361. [Google Scholar] [CrossRef] [Green Version]
- Kaiko, G.E.; Ryu, S.H.; Koues, O.I.; Collins, P.L.; Solnica-Krezel, L.; Pearce, E.J.; Pearce, E.L.; Oltz, E.M.; Stappenbeck, T.S. The Colonic Crypt Protects Stem Cells from Microbiota-Derived Metabolites. Cell 2016, 165, 1708–1720. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; He, Z.; Chen, W.; Holzman, I.R.; Lin, J. Effects of Butyrate on Intestinal Barrier Function in a Caco-2 Cell Monolayer Model of Intestinal Barrier. Pediatr. Res. 2007, 61, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Perman, J.A.; Modler, S.; Olson, A.C. Role of PH in Production of Hydrogen from Carbohydrates by Colonic Bacterial Flora. Studies In Vivo and In Vitro. J. Clin. Investig. 1981, 67, 643–650. [Google Scholar] [CrossRef]
- Breuer, R.I.; Buto, S.K.; Christ, M.L.; Bean, J.; Vernia, P.; Paoluzi, P.; Di Paolo, M.C.; Caprilli, R. Rectal Irrigation with Short-Chain Fatty Acids for Distal Ulcerative Colitis: Preliminary Report. Dig. Dis. Sci. 1991, 36, 185–187. [Google Scholar] [CrossRef]
- Scheppach, W.; Sommer, H.; Kirchner, T.; Paganelli, G.-M.; Bartram, P.; Christl, S.; Richter, F.; Dusel, G.; Kasper, H. Effect of Butyrate Enemas on the Colonic Mucosa in Distal Ulcerative Colitis. Gastroenterology 1992, 103, 51–56. [Google Scholar] [CrossRef]
- Scheppach, W.; German-Austrian Scfa Study Group. Treatment of Distal Ulcerative Colitis with Short-Chain Fatty Acid Enemas a Placebo-Controlled Trial. Dig. Dis. Sci. 1996, 41, 2254–2259. [Google Scholar] [CrossRef] [PubMed]
- Vernia, P.; Marcheggiano, A.; Caprilli, R.; Frieri, G.; Corrao, G.; Valpiani, D.; Paolo, M.C.D.; Paoluzi, P.; Torsoli, A. Short-Chain Fatty Acid Topical Treatment in Distal Ulcerative Colitis. Aliment. Pharmacol. Ther. 2007, 9, 309–313. [Google Scholar] [CrossRef]
- Steinhart, A.H.; Hiruki, T.; Brzezinski, A.; Baker, J.P. Treatment of Left-Sided Ulcerative Colitis with Butyrate Enemas: A Controlled Trial. Aliment. Pharmacol. Ther. 1996, 10, 729–736. [Google Scholar] [CrossRef]
- Breuer, R.I.; Soergel, K.H.; Lashner, B.A.; Christ, M.L.; Hanauer, S.B.; Vanagunas, A.; Harig, J.M.; Keshavarzian, A.; Robinson, M.; Sellin, J.H.; et al. Short Chain Fatty Acid Rectal Irrigation for Left-Sided Ulcerative Colitis: A Randomised, Placebo Controlled Trial. Gut 1997, 40, 485–491. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Drug | Patients | Outcome | Ref. |
|---|---|---|---|
| anti-TNF-α | 56 UC (50 IFX, 6 ADA) | Lower dysbiosis indexes and higher abundance of Faecalibacterium prausnitzii predict responders | [10] |
| vedolizumab | 43 UC, 42 CD | Butyrate producer promote response of vedolizmab treatment | [7] |
| anti-TNF-α | 18 CD, 17 UC | In low response patients, lower exchanged among bacterial communities of butyrate, ethanol or acetaldehyde (involved in butyrate synthesis) | [9] |
| azathioprine | 43 CD, 22 UC | Butyrate production of patients in remission is at baseline higher compared with patients without remission | [8] |
| Disease | Sample Size | Treatment | Dose, Duration | Outcome | Ref. |
|---|---|---|---|---|---|
| distal UC | 12 | SCFAs (80:30:40) | 100 mL, twice daily, 6 weeks | 9 patients improved disease activity index and mucosal histrogical scores | [80] |
| 10 | 100 mM butyrate vs. placebo | 100 mL, twice daily, 2 weeks | Improving endoscopic and histrogical scores | [81] | |
| 47 | 130 mM SCFAs (46:23:31) or 100 mM butyrate alone vs. saline | 60 mL, twice daily, 8 weeks | No differences in disease activity, clinical response, endoscopic, histological scores | [82] | |
| 40 | 150 mM SCFAs (53:20:27) vs. saline | 100 mL, twice daily, 6 weeks | Improving intestinal bleeding, urgency, the patient self-evaluation scores | [83] | |
| 38 | 80 mM butyrate vs. saline | 60 mL, once nightly, 6 weeks | No difference in clinical disease activity index scores | [84] | |
| 103 | 150 mM SCFAs (53:20:27) vs. saline | 100 mL, twice daily, 6 weeks | No difference in clinical and histological activity scores | [85] |
| Model | Result | Mechanism | Ref. |
|---|---|---|---|
| DSS induced colitis mice with antibiotics pretreatment | Not ameliorate colitis | Decreased IL-6 production by butyrate treatment inhibit intestinal tissue repair and cytoprotection | [12] |
| Caco-2 cells | Low concentration; intestinal barrier ↑ | Excessive butyrate induces epithelial cell apoptosis | [78] |
| Excessive concentration; intestinal barrier ↓ | |||
| Primary intestinal epithelial monolayer cultures from UC patients + TNF-α and IFN-γ | Intestinal barrier ↓, IL-8 mRNA ↑ many inflammatory protein ↑ | unknown (There is a possibility that butyrate suppresses the expansion of proliferating cells under inflammatory mediators) | [76] |
| Mouse and human colonic and small intestinal crypts | Suppressing colonic epithelial stem/progenitor cell proliferation, delayed wound repair | By HDAC inhibition, promoter activity for the negative cell-cycle regulator Foxo3 was increased | [77] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ota, S.; Sakuraba, H. Uptake and Advanced Therapy of Butyrate in Inflammatory Bowel Disease. Immuno 2022, 2, 692-702. https://doi.org/10.3390/immuno2040042
Ota S, Sakuraba H. Uptake and Advanced Therapy of Butyrate in Inflammatory Bowel Disease. Immuno. 2022; 2(4):692-702. https://doi.org/10.3390/immuno2040042
Chicago/Turabian StyleOta, Shinji, and Hirotake Sakuraba. 2022. "Uptake and Advanced Therapy of Butyrate in Inflammatory Bowel Disease" Immuno 2, no. 4: 692-702. https://doi.org/10.3390/immuno2040042
APA StyleOta, S., & Sakuraba, H. (2022). Uptake and Advanced Therapy of Butyrate in Inflammatory Bowel Disease. Immuno, 2(4), 692-702. https://doi.org/10.3390/immuno2040042