
Assessing Radiation Effects on Chemo-Treated BT20 and 4T1 Breast Cancer, and Neuroblastoma Cell Lines: A Study of Single and Multiple-Cell Ionization via Infrared Laser Trapping

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Overview
2.2. DMDD Extraction and Preparation
2.3. Cell Culture and Treatment
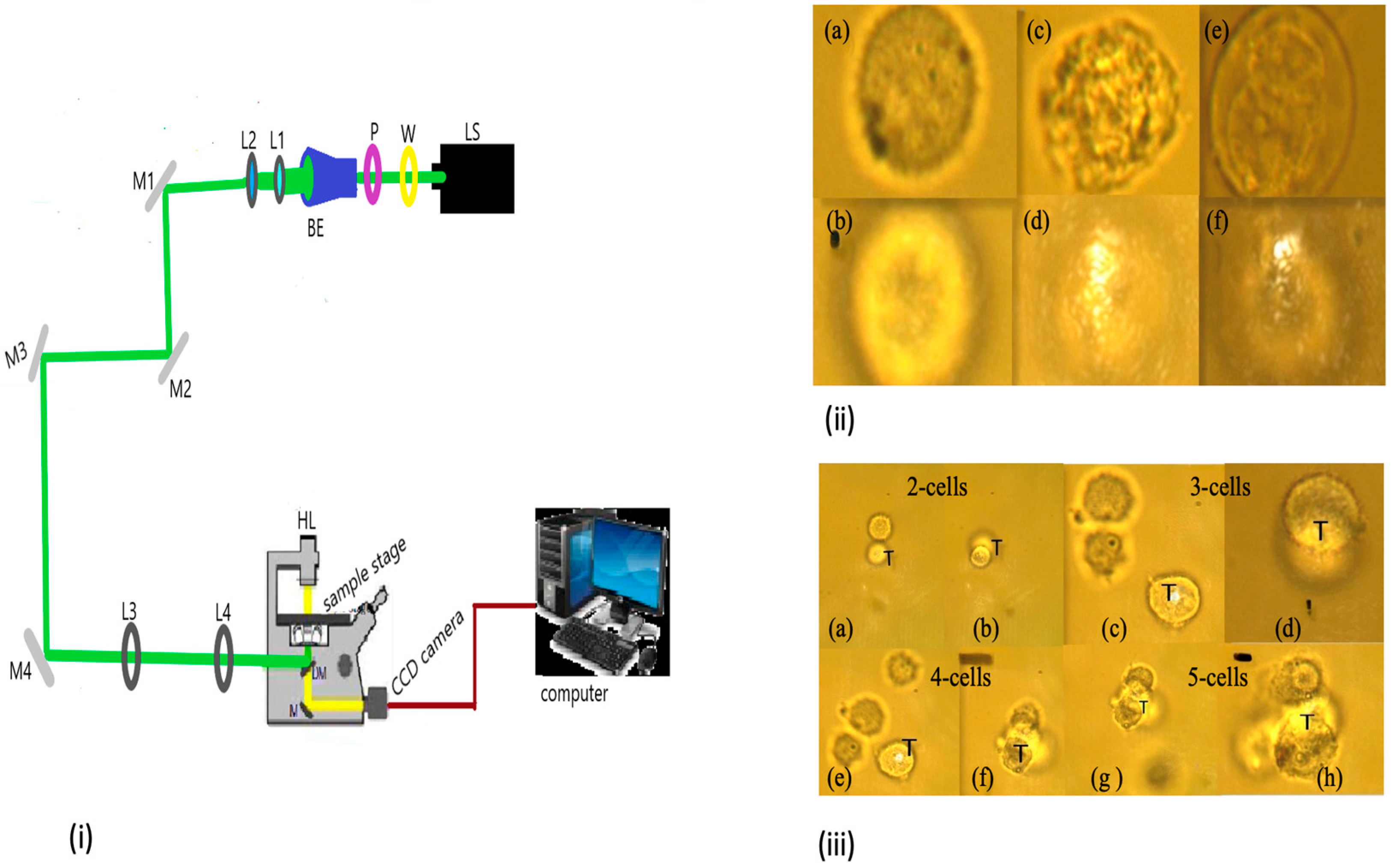
2.4. Laser Trapping and Ionization Setup
2.5. Quantitative Analysis and Comparison
2.6. Mechanistic Insights and Comparative Analysis
2.7. Summary
3. Results and Discussion
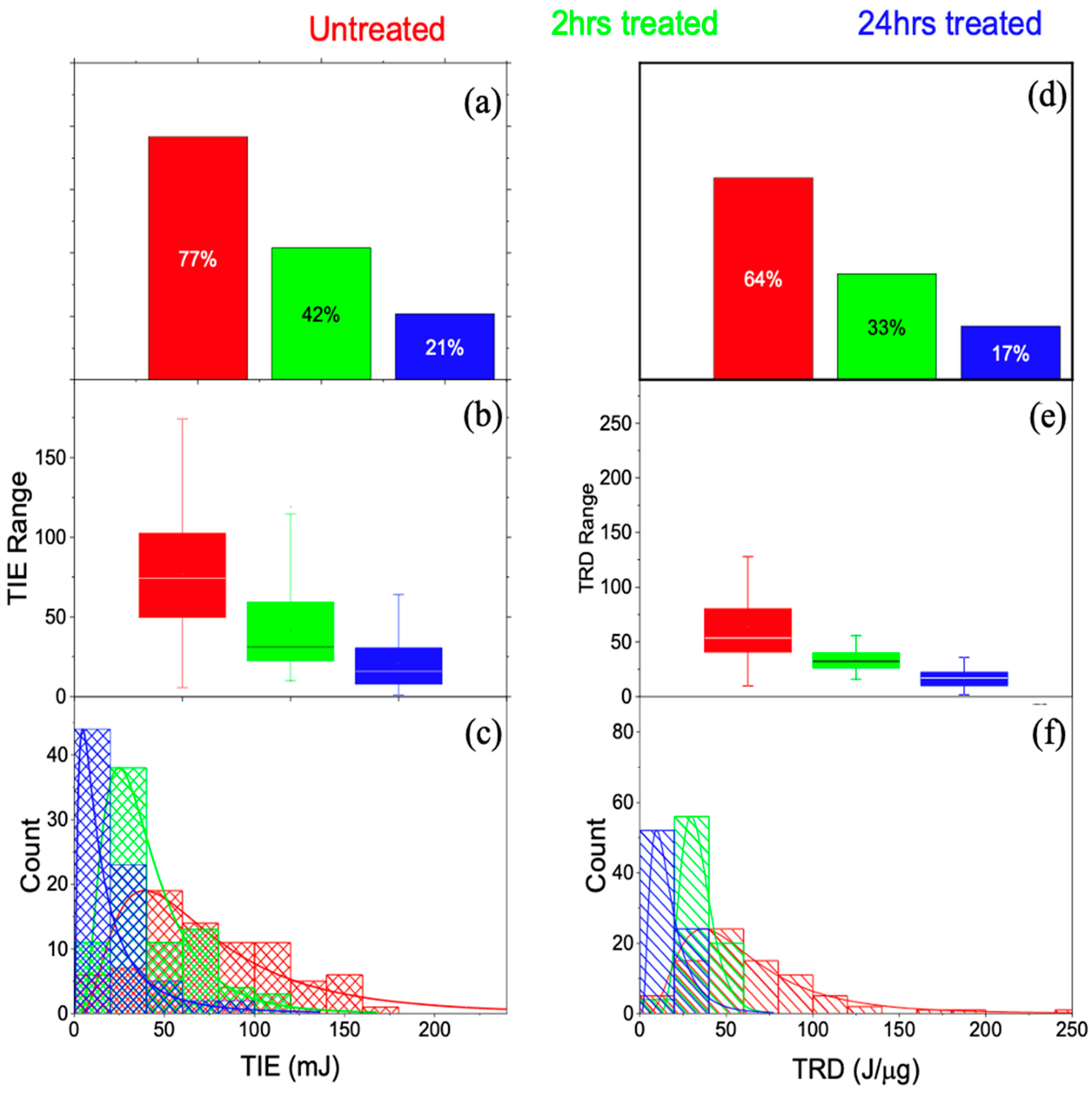
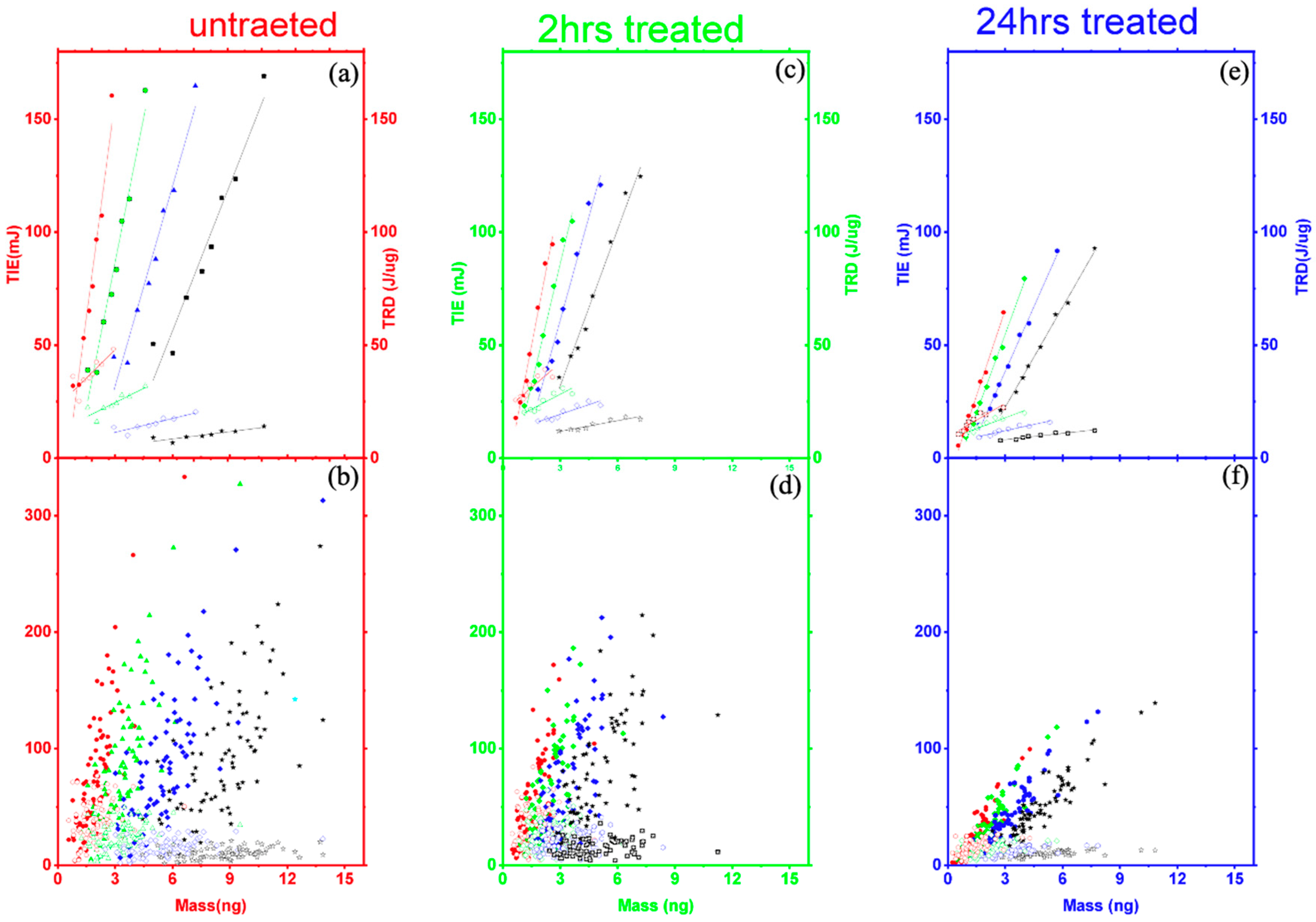
3.1. Single-Cell Ionization
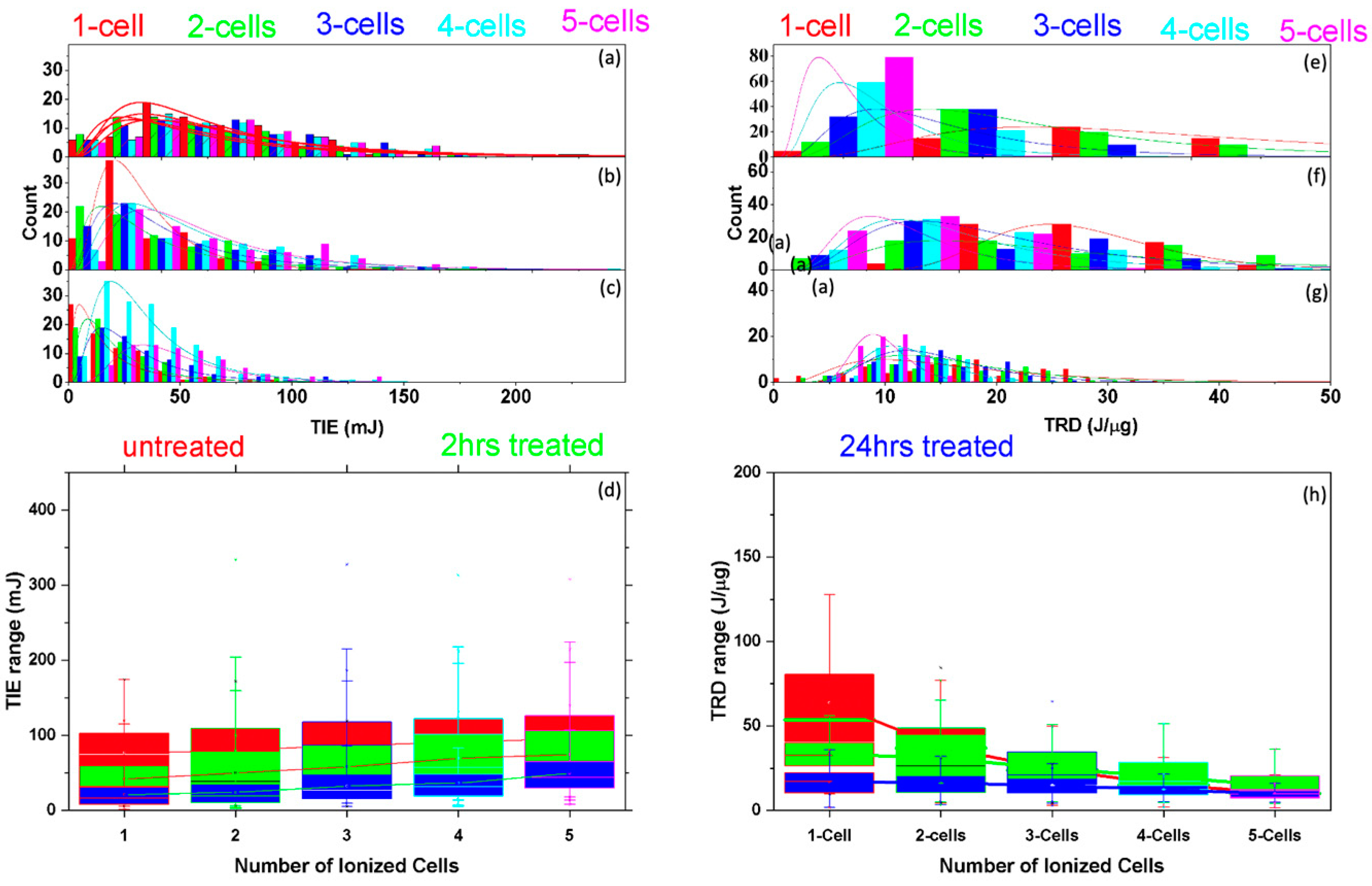
3.2. Multiple Cell Ionization
3.3. Mechanistic Framework
3.3.1. Threshold Ionization Energy and Radiation Dose Theory
3.3.2. The Mechanism of Cell Ionization
4. The Comparison with N2a and 4T1 Cells Lines
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Health Observatory; World Health Organization: Geneva, Switzerland, 2019; Available online: https://who.int/gho/database/en/ (accessed on 21 June 2019).
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Bauer, K.R.; Brown, M.; Cress, R.D.; Parise, C.A.; Caggiano, V. Descriptive analysis of estrogen receptor (ER)-negative, progesterone receptor (PR)-negative, and HER2-negative invasive breast cancer, the so-called triple-negative phenotype: A population-based study from the California cancer Registry. Cancer 2007, 109, 1721–1728. [Google Scholar] [CrossRef] [PubMed]
- Sugita, B.; Gill, M.; Mahajan, A.; Duttargi, A.; Kirolikar, S.; Almeida, R.; Regis, K.; Oluwasanmi, O.L.; Marchi, F.; Marian, C.; et al. Differentially expressed miRNAs in triple negative breast cancer between African-American and non-Hispanic white women. Oncotarget 2016, 7, 79274. [Google Scholar] [CrossRef] [PubMed]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.W. Cancer and radiation therapy: Current advances and future directions. Int. J. Med. Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef]
- Liauw, S.L.; Connell, P.P.; Weichselbaum, R.R. New paradigms and future challenges in radiation oncology: An update of biological targets and technology. Sci. Transl. Med. 2013, 5, 173sr2. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.J.; Khin, N.Y.; Pavlakis, N.; Hugh, T.J.; Clarke, S.J.; Magnussen, J.; Rogan, C.; Flekser, R.L. Challenges in chemotherapy delivery: Comparison of standard chemotherapy delivery to locoregional vascular mass fluid transfer. Future Oncol. 2018, 14, 647–663. [Google Scholar] [CrossRef]
- Wang, L.; Correa, C.R.; Zhao, L.; Hayman, J.; Kalemkerian, G.P.; Lyons, S.; Cease, K.; Brenner, D.; Kong, F.-M. The effect of radiation dose and chemotherapy on overall survival in 237 patients with stage III non-small cell lung cancer. Int. J. Radiat. Oncol. Biol. Phys. 2009, 73, 1383–1390. [Google Scholar] [CrossRef]
- Begg, A.C.; Stewart, F.A.; Vens, C. Strategies to improve radiotherapy with targeted drugs. Nat. Rev. Cancer 2011, 11, 239–253. [Google Scholar] [CrossRef]
- Maingon, P.; Govaerts, A.-S.; Rivera, S.; Vens, C.; Shash, E.; Grégoire, V. New challenge of developing combined radio-drug therapy. Chin. Clin. Oncol. 2014, 3, 18. [Google Scholar] [CrossRef]
- Yoo, G.S.; Park, W.; Yu, J.I.; Choi, D.H.; Kim, Y.-J.; Shin, K.H.; Wee, C.W.; Kim, K.; Park, K.R.; Kim, Y.B.; et al. Comparison of Breast Conserving Surgery Followed by Radiation Therapy with Mastectomy Alone for Pathologic N1 Breast Cancer Patients in the Era of Anthracycline Plus Taxane-Based Chemotherapy: A Multicenter Retrospective Study (KROG 1418). Cancer Res. Treat. 2019, 51, 1041–1051. [Google Scholar] [CrossRef]
- Nguyen, P.T.; Abbosh, A.; Crozier, S. Three-Dimensional Microwave Hyperthermia for Breast Cancer Treatment in a Realistic Environment Using Particle Swarm Optimization. IEEE Trans. Biomed. Eng. 2017, 64, 1335–1344. [Google Scholar] [CrossRef]
- Ami, N.; Sato, H.; Hayakawa, Y. Paclitaxel-induced hypothermia and hypoperfusion increase breast cancer metastasis and angiogenesis in mice. Oncol. Lett. 2018, 15, 2330–2334. [Google Scholar] [CrossRef]
- Norouzi, H.; Khoshgard, K.; Akbarzadeh, F. In vitro outlook of gold nanoparticles in photo-thermal therapy: A literature review. Lasers Med. Sci. 2018, 33, 917. [Google Scholar] [CrossRef]
- Rastinehad, A.R.; Anastos, H.; Wajswol, E.; Winoker, J.S.; Sfakianos, J.P.; Doppalapudi, S.K.; Carrick, M.R.; Knauer, C.J.; Taouli, B.; Lewis, S.C.; et al. Gold nanoshell-localized photothermal ablation of prostate tumors in a clinical pilot device study. Proc. Natl. Acad. Sci. USA 2019, 116, 18590–18596. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, J. Current status and future directions of cancer immunotherapy. J. Cancer 2018, 9, 1773–1781. [Google Scholar] [CrossRef] [PubMed]
- Malekian, S.; Rahmati, M.; Sari, S.; Kazemimanesh, M.; Kheirbakhsh, R.; Muhammadnejad, A.; Amanpour, S. Expression of Diverse Angiogenesis Factor in Different Stages of the 4T1 Tumor as a Mouse Model of Triple-Negative Breast Cancer. Adv. Pharm. Bull. 2020, 10, 323. [Google Scholar] [CrossRef] [PubMed]
- Muhammed, E.; Chen, L.; Gao, Y.; Erenso, D. Chemo treated 4T1 breast cancer cells radiation response measured by single and multiple cells ionization using infrared laser trap. Sci. Rep. 2019, 9, 17547. [Google Scholar] [CrossRef] [PubMed]
- Muhammed, E.; Erenso, D.B.; Gao, Y.; Chen, L.; Kelley, M.; Vazquez, C.; Gale, M.; Nichols, C.; Crogman, H.T. Measurement of Charge and Refractive Indices in Optically Trapped and Ionized Living Cells. Tomography 2023, 9, 70–88. [Google Scholar] [CrossRef] [PubMed]
- Goangul, M.S.; Solomon, R.M.; Devito, D.L.; Brown, C.A.; Coopper, J.; Erenso, D.B.; Gao, Y.; Pellizzaro, A.; Revalee, J.M.; Crogman, H.T. The Effectiveness of Suffruticosol B in Treating Lung Cancer by the Laser Trapping Technique. Biophysica 2023, 3, 109–120. [Google Scholar] [CrossRef]
- Kelley, M.; Gao, Y.; Erenso, D. Single cell ionization by a laser trap: A preliminary study in measuring radiation dose and charge in BT20 breast carcinoma cells. Biomed. Opt. Express 2016, 7, 3438–3448. [Google Scholar] [CrossRef] [PubMed]
- Kelley, M.; Devito, D.; Mushi, R.; Aguinaga, M.d.P.; Erenso, D.B. Laser trap ionization for identification of human erythrocytes with variable hemoglobin quantitation. J. Biomed. Opt. 2018, 23, 055005. [Google Scholar] [CrossRef]
- Pasquerilla, M.; Kelley, M.; Mushi, R.; Aguinaga, M.D.P.; Erenso, D. Laser trapping ionization of single human red blood cell. Biomed. Phys. Eng. Express 2018, 4, 045020. [Google Scholar] [CrossRef]
- Pellizzaro, A.; Welker, G.; Scott, D.; Solomon, R.; Cooper, J.; Farone, A.; Farone, M.; Mushi, R.S.; Aguinaga, M.d.P.; Erenso, D. Direct laser trapping for measuring the behavior of transfused erythrocytes in a sickle cell anemia patient. Biomed. Opt. Express 2012, 3, 2190–2199. [Google Scholar] [CrossRef]
- Goangul, M.S.; Stewart, W.C.; Erenso, D.; Crogman, H.T. The radiation response measurement of a single and multiple cell ionization of neuroblastoma cells by infrared laser trap. J. Radiat. Res. 2022, 64, 113–125. [Google Scholar] [CrossRef]
- Ashkin, A. Applications of laser radiation pressure. Science 1980, 210, 1081–1088. [Google Scholar] [CrossRef]
- Chu, S.; Bjorkholm, J.E.; Ashkin, A.; Cable, A. Experimental observation of optically trapped atoms. Phys. Rev. Lett. 1986, 57, 314. [Google Scholar] [CrossRef]
- Neuman, K.C.; Block, S.M. Optical trapping. Rev. Sci. Instrum. 2004, 75, 2787–2809. [Google Scholar] [CrossRef] [PubMed]
- Essiambre, R.J. Arthur Ashkin: Father of the optical tweezers. Proc. Natl. Acad. Sci. USA 2021, 118, e2026827118. [Google Scholar] [CrossRef] [PubMed]
- Boyd, R. Contemporary Nonlinear Optics; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Keldysh, L.V. Ionization in the field of a strong electromagnetic wave. J. Exp. Theor. Phys. 1965, 20, 1307–1314. [Google Scholar]
- Miziolek, A.W.; Palleschi, V.; Schechter, I. (Eds.) Laser Induced Breakdown Spectroscopy; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Dahotre, N.B.; Samant, A. Laser Machining of Advanced Materials; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Corkum, P.Á.; Krausz, F. Attosecond science. Nat. Phys. 2007, 3, 381–387. [Google Scholar] [CrossRef]
- Peterman, E.J.; Gittes, F.; Schmidt, C.F. Laser-induced heating in optical traps. Biophys. J. 2003, 84, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Tianen, Y. A study of the radiosensitive effects on mammary carcinoma in mice by Chinese medicine (Salvia plus Astragalus) and aspirin. Chin. J. Cancer Res. 1989, 1, 54–59. [Google Scholar] [CrossRef]
- Zhang, P.; Cui, Z.; Liu, Y.; Wang, D.; Liu, N.; Yoshikawa, M. Quality evaluation of traditional Chinese drug toad venom from different origins through a simultaneous determination of bufogenins and indole alkaloids by HPLC. Chem. Pharm. Bull. 2005, 53, 1582–1586. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Huang, R.; Gong, Y.; Park, H.S.; Wen, Q.; Almosnid, N.M.; Chippada-Venkata, U.D.; Hosain, N.A.; Vick, E.; Farone, A.; et al. The antidiabetic compound 2-dodecyl-6-methoxycyclohexa-2,5-diene-1,4-dione, isolated from Averrhoa carambola L., demonstrates significant antitumor potential against human breast cancer cells. Oncotarget 2015, 6, 24304–24319. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Nong, Z.; Xie, Q.; He, J.; Cai, W.; Tang, X.; Chen, X.; Huang, R.; Gao, Y. 2-Dodecyl-6-methoxycyclohexa-2,5-diene-1,4-dione inhibits the growth and metastasis of breast carcinoma in mice. Sci. Rep. 2017, 7, 6704. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Guo, S.; Li, Z. In situ characterizing membrane lipid phenotype of breast cancer cells using mass spectrometry profiling. Sci. Rep. 2015, 5, 11298. [Google Scholar] [CrossRef] [PubMed]
- Chavez, K.J.; Garimella, S.V.; Lipkowitz, S. Triple negative breast cancer cell lines: One tool in the search for better treatment of triple negative breast cancer. Breast Dis. 2010, 32, 35. [Google Scholar] [CrossRef] [PubMed]
- Lasfargues, E.Y.; Ozzello, L. Cultivation of human breast carcinomas. J. Natl. Cancer Inst. 1958, 21, 1131–1147. [Google Scholar]
- Pulaski, B.A.; Ostrand-Rosenberg, S. Mouse 4T1 breast tumor model. Current protocols in immunology, American Type Culture Collection (ATCC): BT20 (ATCC HTB-19) Hum. Cells 2000, 39, 20–22. [Google Scholar] [CrossRef]
- Wen, Q.; Lin, X.; Liu, Y.; Xu, X.; Liang, T.; Zheng, N.; Kintoko; Huang, R. Phenolic and Lignan Glycosides from the Butanol Extract of Averrhoa carambola L. Root. Molecules 2012, 17, 12330–12340. [Google Scholar] [CrossRef]
- Ni, Z.; Lin, X.; Wen, Q.; Kintoko; Zhang, S.; Huang, J.; Xu, X.; Huang, R. Effect of 2-dodecyl-6-methoxycyclohexa-2,5-diene-1,4-dione, isolated from Averrhoa carambola L. (Oxalidaceae) roots, on advanced glycation end-product-mediated renal injury in type 2 diabetic kkay mice. Toxicol. Lett. 2021, 339, 88–96. [Google Scholar] [CrossRef]
- Grover, W.H.; Bryan, A.K.; Diez-Silva, M.; Suresh, S.; Higgins, J.M.; Manalis, S.R. Measuring single-cell density. Proc. Natl. Acad. Sci. USA 2011, 108, 10992–10996. [Google Scholar] [CrossRef]
- Zhao, Y.; Lai, H.S.S.; Zhang, G.; Lee, G.B.; Li, W.J. Measurement of single leukemia cell’s density and mass using optically induced electric field in a microfluidics chip. Biomicrofluidics 2015, 9, 022406. [Google Scholar] [CrossRef] [PubMed]
- Baniyash, M.; Netanel, T.; Witz, I.P. Differences in Cell Density Associated with Differences in Lung-colonizing Ability of B16 Melanoma Cells. Cancer Res. 1981, 41, 433–437. [Google Scholar]
- Stenvall, A.; Larsson, E.; Holmqvist, B.; Strand, S.E.; Jönsson, B.A. Quantitative γ-H2AX immunofluorescence method for DNA double-strand break analysis in testis and liver after intravenous administration of 111 InCl 3. EJNMMI Res. 2020, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Mirzayans, R.; Murray, D. Do TUNEL and other apoptosis assays detect cell death in preclinical studies? Int. J. Mol. Sci. 2020, 21, 9090. [Google Scholar] [CrossRef] [PubMed]
- Mah, L.J.; El-Osta, A.; Karagiannis, T.C. γH2AX: A sensitive molecular marker of DNA damage and repair. Leukemia 2010, 24, 679–686. [Google Scholar] [CrossRef] [PubMed]
- van Meer, G.; de Kroon, A.I. Lipid map of the mammalian cell. J. Cell Sci. 2011, 124, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Escribá, P.V.; González-Ros, J.M.; Goñi, F.M.; Kinnunen, P.K.; Vigh, L.; Sánchez-Magraner, L.; Fernández, A.M.; Busquets, X.; Horváth, I.; Barceló-Coblijn, G. Membranes: A meeting point for lipids, proteins and therapies. J. Cell Mol. Med. 2008, 12, 829–875. [Google Scholar] [CrossRef]
- Field, C.J.; Thomson, C.A.; Van Aerde, J.E.; Parrott, A.; Lien, E.; Clandinin, M.T. Lower proportion of CD45R0(+) cells and deficient interleukin-10 production by formula-fed infants, compared with human-fed, is corrected with supplementation of long-chain polyunsaturated fatty acids. J. Pediatr. Gastr. Nutr. 2000, 31, 291–299. [Google Scholar]
- Volmer, R.; van der Ploeg, K.; Ron, D. Membrane lipid saturation activates endoplasmic reticulum unfolded protein response transducers through their transmembrane domains. Proc. Natl. Acad. Sci. USA 2013, 110, 4628–4633. [Google Scholar] [CrossRef] [PubMed]
- Hercules, W.; Lindesay, J.; Coble, A.; Schmukler, R. Electroporation of Biological Cells Embedded in a Polycarbonate Filter; Howard University: Washington, DC, USA, 2003; p. 20059. [Google Scholar]
- Neumann, E.; Sowers, A.; Jordan, C. Electroporation and Electrofusion in Cell. Biology 1990, 94, 1048–1049. [Google Scholar]
- Chang, D.; Chassy, B.; Saunders, J.; Sowers, A. Guide to Electroporation and Electrofusion; Academic Press: New York, NY, USA, 1992. [Google Scholar]
- Bhattacharya, S.; Aroumougame, A. Repurposing DNA repair factors to eradicate tumor cells upon radiotherapy. Transl. Cancer Res. 2017, 6, S822. [Google Scholar] [CrossRef]
- Alhmoud, J.F.; Woolley, J.F.; Al Moustafa, A.E.; Mallei, M.I. DNA damage/repair management in cancers. Cancers 2020, 12, 1050. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.P.; Jiri, B. The DNA-damage response in human biology and disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef]
- Nickoloff, J.A.; Lynn, T.; Neelam, S.; Kato, T.A. Exploiting DNA repair pathways for tumor sensitization, mitigation of resistance, and normal tissue protection in radiotherapy. Cancer Drug Resist. 2021, 4, 244. [Google Scholar] [CrossRef]
- Yuan, A.; Rao, M.-V.; Nixon, R.-A. Neurofilaments and neurofilament proteins in health and disease. Cold Spring Harb. Perspect. Biol. 2017, 9, a018309. [Google Scholar] [CrossRef]
- Zhong, M.C.; Wei, X.B.; Zhou, J.H.; Wang, Z.Q.; Li, Y.M. Trapping red blood cells in living animals using optical tweezers. Nat. Commun. 2013, 4, 1768. [Google Scholar] [CrossRef]
- Hason, M.; Bartůněk, P. Zebrafish models of cancer—New insights on modeling human cancer in a non-mammalian vertebrate. Genes 2019, 10, 935. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
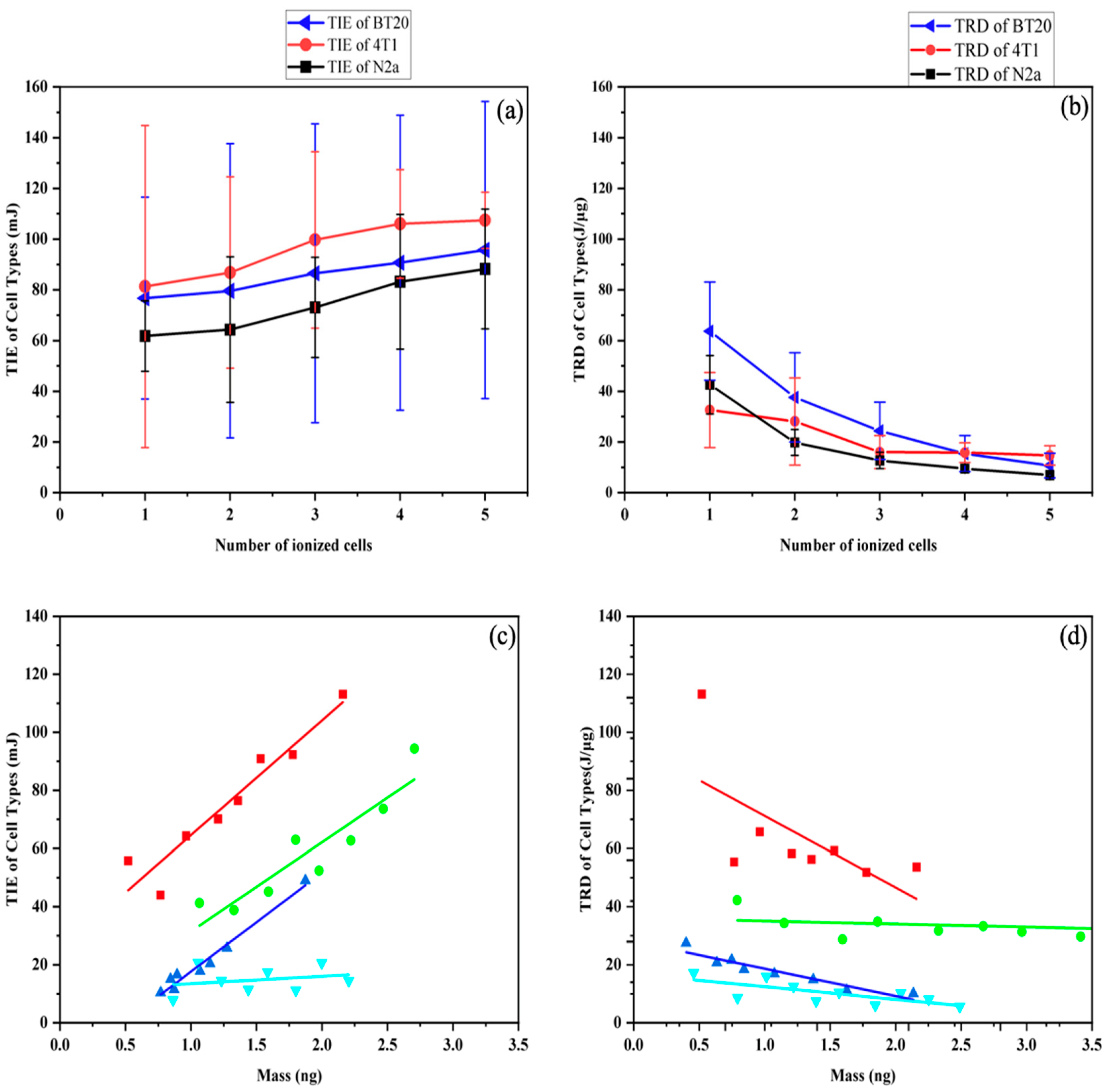{kind=link}
| Homogeneity Variance Test for Untreated, 2 h and 24 h Treated Groups | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TIE | TRD | |||||||||||||||
| DF | SS | MS | F-Value | Prob. > F | DF | SS | MS | F-Value | Prob. > F | |||||||
| Model | 2 | 13,342.8 | 6671.4 | 21.4 | 2.9343 | 2 | 23,704.9 | 11,852.5 | 43.4 | 1.1 | ||||||
| Error | 233 | 72,635.3 | 311.7 | 233 | 63,591.1 | 272.9 | ||||||||||
| Mean comparison for untreated (1), 2 h treated (2) and 24 h treated (3) groups | ||||||||||||||||
| MD | SEM | q-v | Prob. | α | Sign | LCL | UCL | MD | SEM | q-v | Prob. | α | LCL | UCL | ||
| 1 vs. 2 | 55.9 | 4.7 | 16.7 | 0 | 0.05 | 1 | 67.1 | 44.7 | 30.3 | 3.78 | 11.3 | 0 | 0 | 39.2 | 21.3 | |
| 1 vs. 3 | 35.1 | 4.7 | 10.6 | 0 | 0.05 | 1 | 46.1 | 24.1 | 46.2 | 3.83 | 17.2 | 0 | 0 | 55.8 | 37.7 | |
| 2 vs. 3 | 20.8 | 4.7 | 6.2 | 4. | 0.05 | 1 | 9.6 | 31.9 | 16.5 | 3.83 | 6.1 | 7.8 | 0 | 25.5 | 7.4 | |
| Untreated BT20 Group | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No | # of Cells | TIE (mJ) | Mass (ng) | TRD (J/µg) | |||||||||
| Mean | sd | Min | Max | Mean | SD | Min | Max | Mean | sd | Min | Max | ||
| 1 | 80 | 76.7 | 39.8 | 5.7 | 74.3 | 1.3 | 0.7 | 0.3 | 1.3 | 63.7 | 39.4 | 9.8 | 53.6 |
| 2 | 80 | 79.6 | 58.0 | 3.7 | 70.7 | 1.9 | 0.9 | 0.6 | 1.9 | 37.6 | 17.6 | 4.2 | 36.9 |
| 3 | 80 | 86.5 | 58.9 | 5.1 | 78.9 | 3.4 | 1.2 | 1.2 | 3.3 | 24.4 | 11.3 | 2.9 | 23.7 |
| 4 | 80 | 90.7 | 58.2 | 6.5 | 82.7 | 5.6 | 1.7 | 2.5 | 5.6 | 15.4 | 7.1 | 1.9 | 14.9 |
| 5 | 80 | 95.7 | 58.6 | 7.9 | 88.5 | 8.8 | 2.3 | 4.4 | 8.8 | 10.7 | 4.8 | 1.4 | 10.1 |
| 2-h Treated BT20 group | |||||||||||||
| TIE (mJ) | Mass (ng) | TRD (J/µg) | |||||||||||
| 1 | 80 | 41.6 | 25.8 | 9.9 | 31.1 | 1.2 | 0.6 | 0.4 | 1.0 | 33.4 | 9.0 | 15.9 | 32.3 |
| 2 | 80 | 49.8 | 38.5 | 6.2 | 38.3 | 1.5 | 0.7 | 0.5 | 1.3 | 31.1 | 16.7 | 4.7 | 26.1 |
| 3 | 80 | 57.8 | 41.3 | 9.5 | 46.4 | 2.2 | 0.9 | 0.9 | 1.9 | 24.7 | 12.4 | 4.9 | 20.9 |
| 4 | 80 | 69.4 | 46.6 | 13.6 | 57.3 | 3.3 | 1.2 | 1.5 | 2.9 | 20.1 | 9.7 | 4.6 | 16.9 |
| 5 | 80 | 74.5 | 46.2 | 17.8 | 62.7 | 4.9 | 1.6 | 2.4 | 4.4 | 10.4 | 6.7 | 4.1 | 12.4 |
| 24-h Treated BT20 group | |||||||||||||
| TIE (mJ) | Mass (ng) | TRD (J/µg) | |||||||||||
| 1 | 76 | 20.9 | 18.3 | 0.9 | 15.9 | 1.1 | 0.6 | 0.2 | 0.6 | 16.9 | 7.6 | 1.6 | 35.8 |
| 2 | 76 | 24.2 | 19.4 | 2.3 | 19.2 | 1.4 | 0.7 | 0.2 | 4.3 | 16.1 | 6.3 | 0.2 | 31.9 |
| 3 | 76 | 32.3 | 22.9 | 4.8 | 26.7 | 2.0 | 0.9 | 0.5 | 5.7 | 14.8 | 5.0 | 4.3 | 27.5 |
| 4 | 76 | 40.9 | 24.8 | 8.7 | 35.2 | 3.1 | 1.2 | 0.9 | 7.8 | 12.3 | 3.6 | 0.9 | 21.5 |
| 5 | 76 | 48.9 | 25.6 | 13.4 | 43.5 | 4.8 | 1.6 | 1.7 | 10.8 | 9.8 | 2.4 | 4.9 | 15.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goangul, M.S.; Erenso, D.B.; Gao, Y.; Chen, L.; Eshun, K.O.; Alvarez, G.; Crogman, H.T. Assessing Radiation Effects on Chemo-Treated BT20 and 4T1 Breast Cancer, and Neuroblastoma Cell Lines: A Study of Single and Multiple-Cell Ionization via Infrared Laser Trapping. Radiation 2024, 4, 85-100. https://doi.org/10.3390/radiation4010007
Goangul MS, Erenso DB, Gao Y, Chen L, Eshun KO, Alvarez G, Crogman HT. Assessing Radiation Effects on Chemo-Treated BT20 and 4T1 Breast Cancer, and Neuroblastoma Cell Lines: A Study of Single and Multiple-Cell Ionization via Infrared Laser Trapping. Radiation. 2024; 4(1):85-100. https://doi.org/10.3390/radiation4010007
Chicago/Turabian StyleGoangul, Mulugeta S., Daniel B. Erenso, Ying Gao, Li Chen, Kwame O. Eshun, Gisela Alvarez, and Horace T. Crogman. 2024. "Assessing Radiation Effects on Chemo-Treated BT20 and 4T1 Breast Cancer, and Neuroblastoma Cell Lines: A Study of Single and Multiple-Cell Ionization via Infrared Laser Trapping" Radiation 4, no. 1: 85-100. https://doi.org/10.3390/radiation4010007
APA StyleGoangul, M. S., Erenso, D. B., Gao, Y., Chen, L., Eshun, K. O., Alvarez, G., & Crogman, H. T. (2024). Assessing Radiation Effects on Chemo-Treated BT20 and 4T1 Breast Cancer, and Neuroblastoma Cell Lines: A Study of Single and Multiple-Cell Ionization via Infrared Laser Trapping. Radiation, 4(1), 85-100. https://doi.org/10.3390/radiation4010007