
Effects of Night Illumination on Behavior, Body Mass and Learning in Male Zebra Finches

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
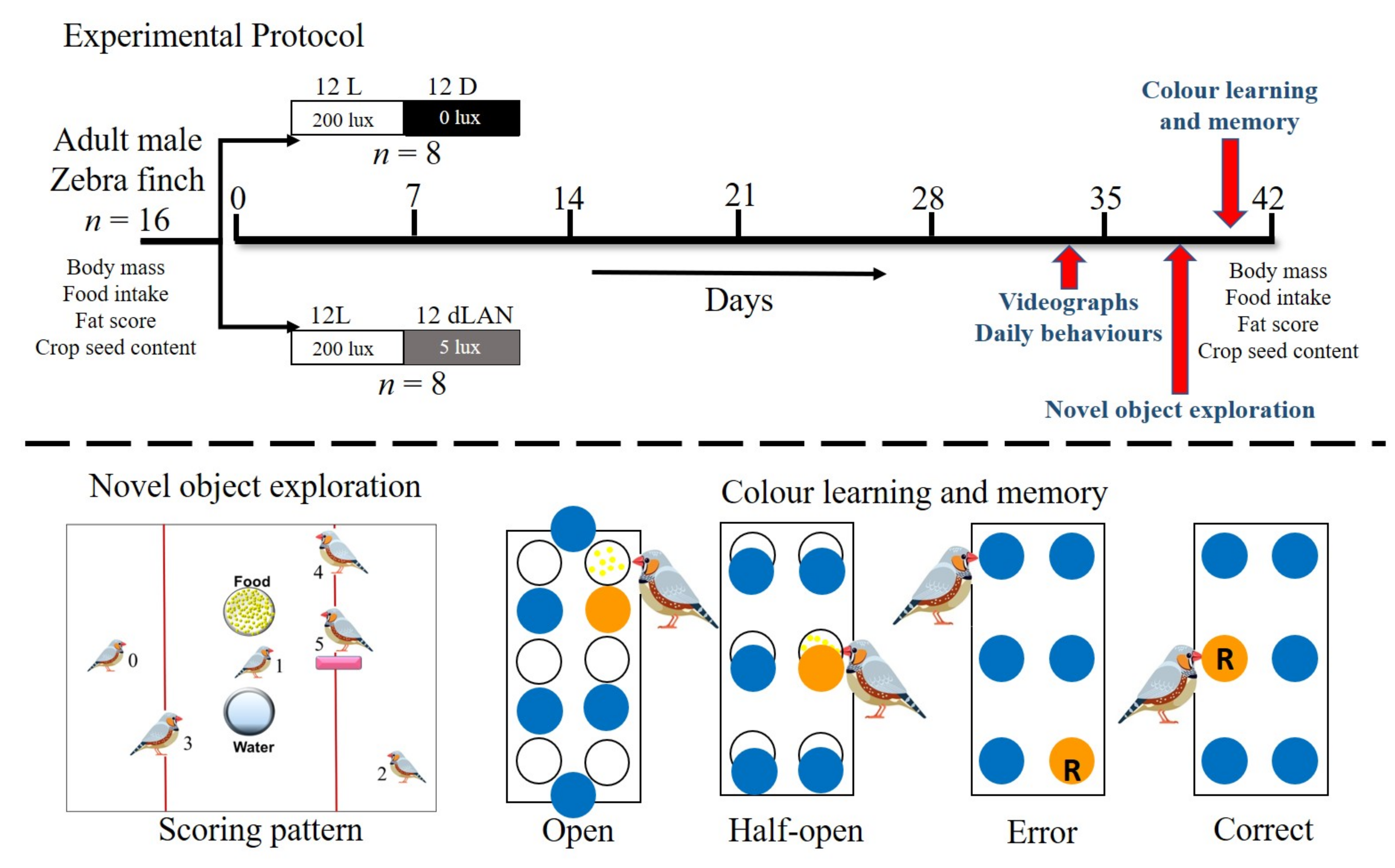
2.1. Animal Husbandry and Experimental Protocol
2.2. Measurement of Body Mass, Food Intake, Seed Storage in Crop and Fat Score
2.3. Assay of Daily Behavior
2.4. Behavioral Assay of Sleep
2.5. Novel Object Exploration (NOE) Test
2.6. Colour Learning and Memory Retrieval
2.7. Statistics
3. Results
3.1. Effect on Daily Behaviors
3.2. Effect on Body Mass, Fat Score, Food Intake and Crop Seed Content
3.3. Effect on Sleep and Novel Object Exploration
3.4. Colour Learning Task
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, V.; Singh, B.P.; Rani, S. The bird clock: A complex, multi-oscillatory and highly diversified system. Biol. Rhythm Res. 2004, 35, 121–144. [Google Scholar] [CrossRef]
- Dunlap, J.C.; Loros, J.J.; DeCoursey, P.J. Chronobiology: Biological Timekeeping, 1st ed.; Sinauer Associates: Sunderland, MA, USA, 2004. [Google Scholar]
- Albrecht, U. Circadian clocks and mood-related behaviors. Handb. Exp. Pharmacol. 2013, 217, 227–239. [Google Scholar]
- Golombek, D.A.; Rosenstein, R.E. Physiology of circadian entrainment. Physiol. Rev. 2010, 90, 1063–1102. [Google Scholar] [CrossRef] [Green Version]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Ann. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunn, R.M.; Blask, D.E.; Coogan, A.N.; Figueiro, M.G.; Gorman, M.R.; Hall, J.E.; Hansen, J.; Nelson, R.J.; Panda, S.; Smolensky, M.H.; et al. Health consequences of electric lighting practices in the modern world: A report on the National Toxicology Program’s workshop on shift work at night, artificial light at night, and circadian disruption. Sci. Total Environ. 2017, 607, 1073–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonken, L.K.; Workman, J.L.; Walton, J.C.; Weil, Z.M.; Morris, J.S.; Haim, A.; Nelson, R.J. Light at night increases body mass by shifting the time of food intake. Proc. Nat. Acad. Sci. USA 2010, 107, 18664–18669. [Google Scholar] [CrossRef] [Green Version]
- Fonken, L.K.; Aubrecht, T.G.; Meléndez-Fernández, O.H.; Weil, Z.M.; Nelson, R.J. Dim light at night disrupts molecular circadian rhythms and increases body weight. J. Biol. Rhythms 2013, 28, 262–271. [Google Scholar] [CrossRef]
- De Jong, M.; Ouyang, J.Q.; van Grunsven, R.H.; Visser, M.E.; Spoelstra, K. Do wild great tits avoid exposure to light at night? PLoS ONE 2016, 11, e0157357. [Google Scholar] [CrossRef]
- Taufique, S.T.; Jha, N.A.; Kumar, V. Circadian rhythm determines the timing of activity, and ingestive and grooming behaviours in Indian house crows, Corvus splendens. Curr. Sci. 2016, 110, 897–901. [Google Scholar]
- Bedrosian, T.A.; Nelson, R.J. Influence of the modern light environment on mood. Mol. Psychiatr. 2013, 18, 751–757. [Google Scholar] [CrossRef]
- Raap, T.; Casasole, G.; Costantini, D.; AbdElgawad, H.; Asard, H.; Pinxten, R.; Eens, M. Artificial light at night affects body mass but not oxidative status in free-living nestling songbirds: An experimental study. Sci. Rep. 2016, 6, 35626. [Google Scholar] [CrossRef]
- Jha, N.A.; Kumar, V. Effect of no-night light environment on behaviour, learning performance and personality in zebra finches. Anim. Behav. 2017, 132, 29–47. [Google Scholar] [CrossRef]
- Taufique, S.T.; Prabhat, A.; Kumar, V. Illuminated night alters hippocampal gene expressions and induces depressive-like responses in diurnal corvids. Eur. J. Neurosci. 2018, 48, 3005–3018. [Google Scholar] [CrossRef]
- Singh, J.; Rani, S.; Kumar, V. Functional similarity in relation to the external environment between circadian behavioral and melatonin rhythms in the subtropical Indian weaver bird. Horm. Behav. 2012, 61, 527–534. [Google Scholar] [CrossRef]
- Raap, T.; Pinxten, R.; Eens, M. Light pollution disrupts sleep in free-living animals. Sci. Rep. 2015, 5, 13557. [Google Scholar] [CrossRef] [Green Version]
- Batra, T.; Malik, I.; Kumar, V. Illuminated night alters behaviour and negatively affects physiology and metabolism in diurnal zebra finches. Environ. Poll. 2019, 254, 112916. [Google Scholar] [CrossRef]
- Batra, T.; Malik, I.; Prabhat, A.; Bhardwaj, S.K.; Kumar, V. Sleep in unnatural times: Illuminated night negatively affects sleep and associated hypothalamic gene expressions in diurnal zebra finches. Proceed. R. Soc. B 2020, 287, 20192952. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Harpole, C.E.; Trivedi, A.K.; Cassone, V.M. Circadian regulation of bird song, call, and locomotor behavior by pineal melatonin in the zebra finch. J. Biol. Rhythms 2012, 27, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Prabhat, A.; Jha, N.A.; Taufique, S.T.; Kumar, V. Dissociation of circadian activity and singing behavior from gene expression rhythms in the hypothalamus, song control nuclei and cerebellum in diurnal zebra finches. Chronobiol. Int. 2019, 36, 1268–1284. [Google Scholar] [CrossRef]
- Prabhat, A.; Malik, I.; Jha, N.A.; Bhardwaj, S.K.; Kumar, V. Developmental effects of constant light on circadian behaviour and gene expressions in zebra finches: Insights into mechanisms of metabolic adaptation to aperiodic environment in diurnal animals. J. Photochem. Photobiol. B 2020, 211, 111995. [Google Scholar] [CrossRef]
- Kumar, J.; Malik, S.; Bhardwaj, S.K.; Rani, S. Bright light at night alters the perception of daylength in Indian weaver bird (Ploceus philippinus). J. Exp. Zool. A Ecol. Integr. Physiol. 2018, 329, 488–496. [Google Scholar] [CrossRef]
- Navara, K.J.; Nelson, R.J. The dark side of light at night: Physiological, epidemiological, and ecological consequences. J. Pineal Res. 2007, 43, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.Q.; de Jong, M.; van Grunsven, R.H.; Matson, K.D.; Haussmann, M.F.; Meerlo, P.; Visser, M.E.; Spoelstra, K. Restless roosts: Light pollution affects behavior, sleep, and physiology in a free-living songbird. Glob. Chang. Biol. 2017, 23, 4987–4994. [Google Scholar] [CrossRef]
- Crusio, W.E. Genetic dissection of mouse exploratory behaviour. Behav. Brain Res. 2001, 125, 127–132. [Google Scholar] [CrossRef]
- Mishra, I.; Kumar, V. The quantity–quality trade-off: Differential effects of daily food availability times on reproductive performance and offspring quality in diurnal zebra finches. J. Exp. Biol. 2019, 222, jeb196667. [Google Scholar] [CrossRef] [Green Version]
- Prabhat, A.; Batra, T.; Kumar, V. Effects of timed food availability on reproduction and metabolism in zebra finches: Molecular insights into homeostatic adaptation to food-restriction in diurnal vertebrates. Horm. Behav. 2020, 125, 104820. [Google Scholar] [CrossRef]
- Griffith, S.C.; Ton, R.; Hurley, L.L.; McDiarmid, C.S.; Pacheco-Fuentes, H. The ecology of the Zebra finch makes it a great laboratory model but an outlier amongst passerine birds. Birds 2021, 2, 60–76. [Google Scholar] [CrossRef]
- Yadav, A.; Kumar, R.; Tiwari, J.; Kumar, V.; Rani, S. Sleep in birds: Lying on the continuum of activity and rest. Biol. Rhythm Res. 2017, 48, 805–814. [Google Scholar] [CrossRef]
- Raap, T.; Pinxten, R.; Eens, M. Artificial light at night causes an unexpected increase in oxalate in developing male songbirds. Conserv. Physiol. 2018, 6, coy005. [Google Scholar] [CrossRef] [PubMed]
- Steinmeyer, C.; Schielzeth, H.; Mueller, J.C.; Kempenaers, B. Variation in sleep behaviour in free-living blue tits, Cyanistes caeruleus: Effects of sex, age and environment. Anim. Behav. 2010, 80, 853–864. [Google Scholar] [CrossRef]
- Mainwaring, M.C.; Beal, J.L.; Hartley, I.R. Zebra finches are bolder in an asocial, rather than social, context. Behav. Process. 2011, 87, 171–175. [Google Scholar] [CrossRef]
- Boogert, N.J.; Giraldeau, L.A.; Lefebvre, L. Song complexity correlates with learning ability in zebra finch males. Anim. Behav. 2008, 76, 1735–1741. [Google Scholar] [CrossRef]
- Fonken, L.K.; Kitsmiller, E.; Smale, L.; Nelson, R.J. Dim nighttime light impairs cognition and provokes depressive-like responses in a diurnal rodent. J. Biol. Rhythms 2012, 27, 319–327. [Google Scholar] [CrossRef]
- Cassone, V.M.; Westneat, D.F. The bird of time: Cognition and the avian biological clock. Front. Mol. Neurosci. 2012, 5, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, N.; Mishra, I.; Komal, R.; Rani, S.; Kumar, V. Circannual testis and moult cycles persist under photoperiods that disrupt circadian activity and clock gene cycles in spotted munia. J. Exp. Biol. 2017, 220, 4162–4168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonken, L.K.; Nelson, R.J. The effects of light at night on circadian clocks and metabolism. Endocr. Rev. 2014, 35, 648–670. [Google Scholar] [CrossRef]
- Plano, S.A.; Casiraghi, L.P.; García Moro, P.; Paladino, N.; Golombek, D.A.; Chiesa, J.J. Circadian and metabolic effects of light: Implications in weight homeostasis and health. Front. Neurol. 2017, 8, 558. [Google Scholar] [CrossRef] [Green Version]
- Coomans, C.P.; van den Berg, S.A.; Houben, T.; van Klinken, J.B.; van den Berg, R.; Pronk, A.C.; Havekes, L.M.; Romijn, J.A.; van Dijk, K.W.; Biermasz, N.R.; et al. Detrimental effects of constant light exposure and high-fat diet on circadian energy metabolism and insulin sensitivity. FASEB J. 2017, 27, 1721–1732. [Google Scholar] [CrossRef] [Green Version]
- Borniger, J.C.; Weil, Z.M.; Zhang, N.; Nelson, R.J. Dim light at night does not disrupt timing or quality of sleep in mice. Chronobiol. Int. 2013, 30, 1016–1023. [Google Scholar] [CrossRef]
- Leveau, L.M. Artificial light at night (ALAN) is the main driver of nocturnal feral pigeon (Columba livia f. domestica) foraging in urban areas. Animals 2020, 10, 554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, A.; Samplonius, J.M.; Schlicht, E.; Valcu, M.; Kempenaers, B. Artificial night lighting rather than traffic noise affects the daily timing of dawn and dusk singing in common European songbirds. Behav. Ecol. 2014, 25, 1037–1047. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V. Avian photoreceptors and their role in the regulation of daily and seasonal physiology. Gen. Comp. Endocrinol. 2015, 220, 13–22. [Google Scholar] [CrossRef]
- Hattar, S.; Kumar, M.; Park, A.; Tong, P.; Tung, J.; Yau, K.W.; Berson, D.M. Central projections of melanopsin-expressing retinal ganglion cells in the mouse. J. Comp. Neurol. 2006, 497, 326–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romeo, S.; Viaggi, C.; Di Camillo, D.; Willis, A.W.; Lozzi, L.; Rocchi, C.; Capannolo, M.; Aloisi, G.; Vaglini, F.; Maccarone, R.; et al. Bright light exposure reduces TH-positive dopamine neurons: Implications of light pollution in Parkinson’s disease epidemiology. Sci. Rep. 2013, 3, 1395. [Google Scholar] [CrossRef] [PubMed]
- Taufique, S.T.; Kumar, V. Differential activation and tyrosine hydroxylase distribution in the hippocampal, pallial and midbrain brain regions in response to cognitive performance in Indian house crows exposed to abrupt light environment. Behav. Brain Res. 2016, 314, 21–29. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. Effect of dLAN on change in body mass, food intake, crop seed content, and fat score (final-initial value) | ||||
| Parameter | LD | dLAN | Analysis | |
| ∆Body mass (g) | −0.08 ± 0.27 | 1.25 ± 0.35 | t14 = 2.98; p = 0.01 | |
| ∆Food intake (g) | Day: −0.24 ± 0.31 Night: 0.0 ± 0.0 | Day: −1.2 ± 0.29 Night: 1.02 ± 0.21 | t14 = 3.62; p = 0.003 t14 = 4.71; p < 0.001 | |
| ∆Crop seed content (at lights on) | 0 | 12.88 ± 4.24 | t14 = 3.03; p = 0.009 | |
| ∆Fat score (median) | 0 | 3 | Mann-Whitney U = 10.50; p = 0.02 | |
| B. Effect of dLAN on daily activity (perch-hopping and preening), feeding, drinking, and preening | ||||
| Parameter | Time | LD | dLAN | Analysis |
| Feeding frequency | Day | 26.06 ± 3.58 | 20.88 ± 3.56 | t14 = 1.03; p = 0.32 |
| Night | 0.0 ± 0.0 | 14.06 ± 1.99 | t14 = 7.06; p < 0.0001 | |
| Feeding duration (min) | Day | 105.9 ± 19.48 | 70.91 ± 15.42 | t14 = 1.40; p = 0.18 |
| Night | 0.0 ± 0.0 | 39.07 ± 4.97 | t14 = 7.86; p < 0.0001 | |
| Perch hopping | Day | 1752 ± 405.5 | 1728 ± 552.8 | t13 = 0.03; p = 0.97 |
| Night | 0.0 ± 0.0 | 401.9 ± 174.6 | t13 = 2.30; p = 0.04 | |
| Drinking frequency | Day | 20.44 ± 1.32 | 26.19 ± 6.07 | t14 = 0.93; p = 0.37 |
| Night | 0.0 ± 0.0 | 8.88 ± 2.62 | t14 = 3.39; p = 0.004 | |
| Drinking duration (min) | Day | 8.68 ± 2.35 | 8.84 ± 3.27 | t14 = 0.04; p = 0.97 |
| Night | 0.0 ± 0.0 | 3.66 ± 1.63 | t14 = 2.24; p = 0.04 | |
| Preening frequency | Day | 38.13 ± 7.26 | 30.81 ± 5.41 | t14 = 0.43; p = 0.43 |
| Night | 0.0 ± 0.0 | 10.25 ± 1.69 | t14 = 6.06; p < 0.0001 | |
| LD | dLAN | |||
|---|---|---|---|---|
| Bird | 1-h RI | 24-h RI | 1-h RI | 24-h RI |
| 1 | ✓ ✓ ✓ ✓ ✓ | ✓ X ✓ ✓ ✓ | ----- | ----- |
| 2 | ----- | ----- | ✓ ✓ N ✓ N | ✓ ✓ ✓ N N |
| 3 | ✓ ✓ ✓ ✓ ✓ | ✓ ✓ ✓ ✓ ✓ | ----- | ----- |
| 4 | ✓ ✓ ✓ ✓ ✓ | ✓ ✓ ✓ ✓ ✓ | ✓ ✓ N N ✓ | ✓ ✓ N ✓ N |
| 5 | ✓ ✓ ✓ ✓ ✓ | ✓ ✓ ✓ ✓ ✓ | ✓ X X ✓✓ | X ✓ X ✓ N |
| 6 | ----- | ----- | ✓ ✓ ✓ ✓ ✓ | ✓ ✓ ✓ ✓ ✓ |
| 7 | ✓ ✓ ✓ ✓ ✓ | ✓ ✓ ✓ ✓ ✓ | ----- | ----- |
| 8 | ✓ ✓ ✓ ✓ N | X ✓ ✓ ✓ ✓ | ✓ ✓ N ✓ X | ✓ ✓ ✓ X ✓ |
| X | X | |||
| X | X | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prabhat, A.; Kumar, M.; Kumar, A.; Kumar, V.; Bhardwaj, S.K. Effects of Night Illumination on Behavior, Body Mass and Learning in Male Zebra Finches. Birds 2021, 2, 381-394. https://doi.org/10.3390/birds2040028
Prabhat A, Kumar M, Kumar A, Kumar V, Bhardwaj SK. Effects of Night Illumination on Behavior, Body Mass and Learning in Male Zebra Finches. Birds. 2021; 2(4):381-394. https://doi.org/10.3390/birds2040028
Chicago/Turabian StylePrabhat, Abhilash, Mayank Kumar, Ashwani Kumar, Vinod Kumar, and Sanjay K. Bhardwaj. 2021. "Effects of Night Illumination on Behavior, Body Mass and Learning in Male Zebra Finches" Birds 2, no. 4: 381-394. https://doi.org/10.3390/birds2040028
APA StylePrabhat, A., Kumar, M., Kumar, A., Kumar, V., & Bhardwaj, S. K. (2021). Effects of Night Illumination on Behavior, Body Mass and Learning in Male Zebra Finches. Birds, 2(4), 381-394. https://doi.org/10.3390/birds2040028