
Long-Term Population Trends of House Sparrow and Eurasian Tree Sparrow in Spain

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
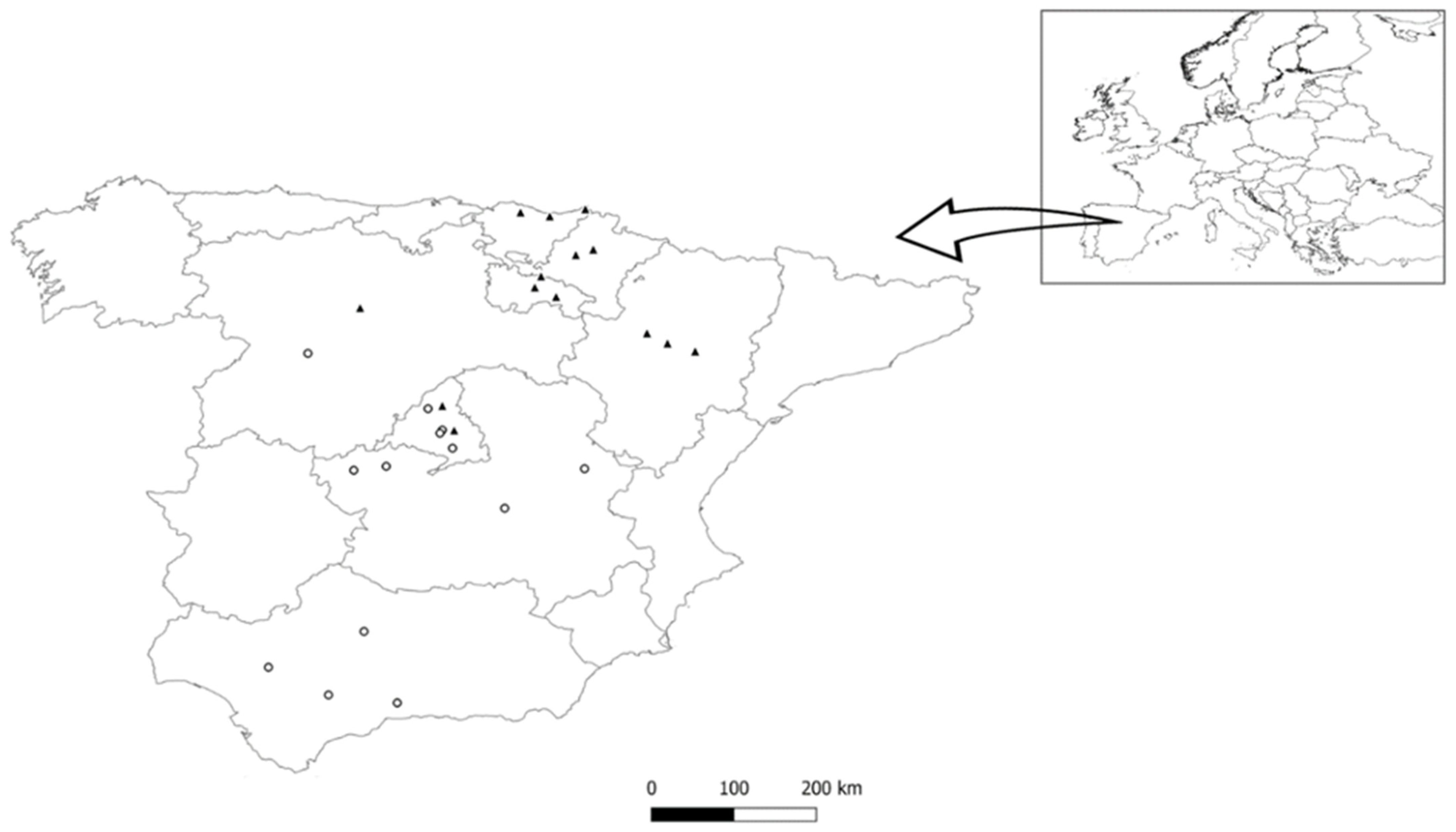
2.1. Sparrow Data
2.2. Statistical Methods
3. Results
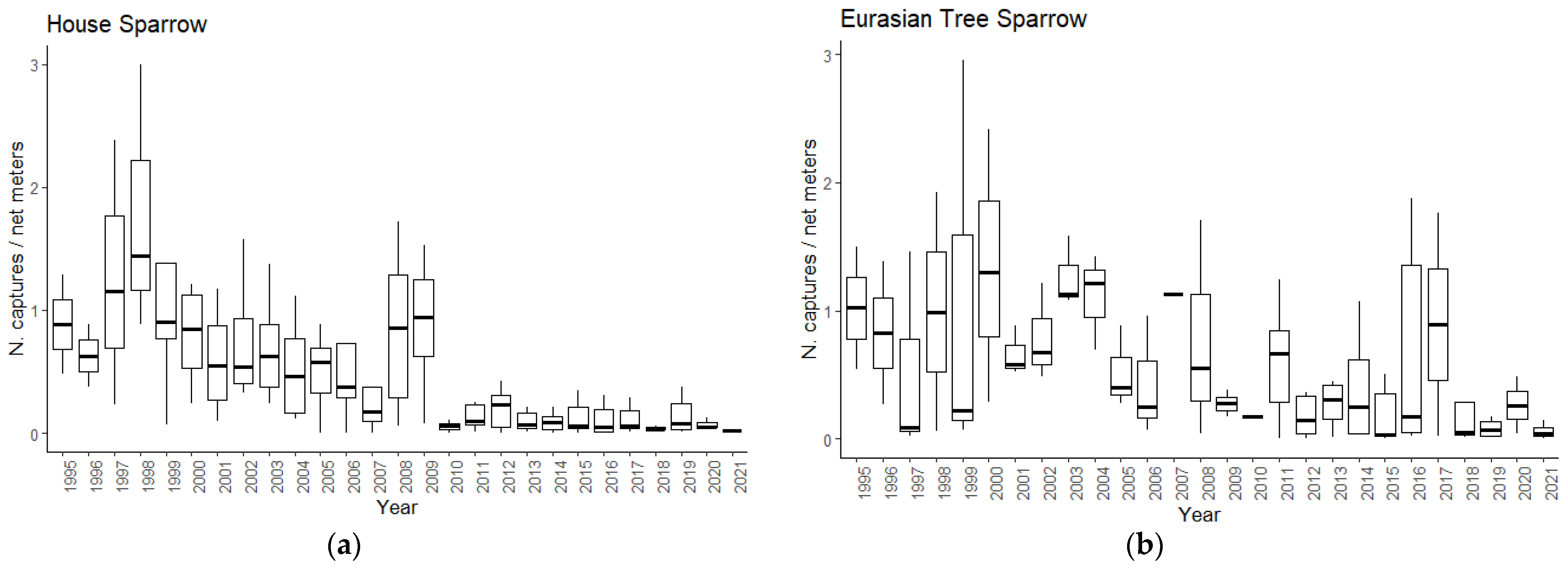
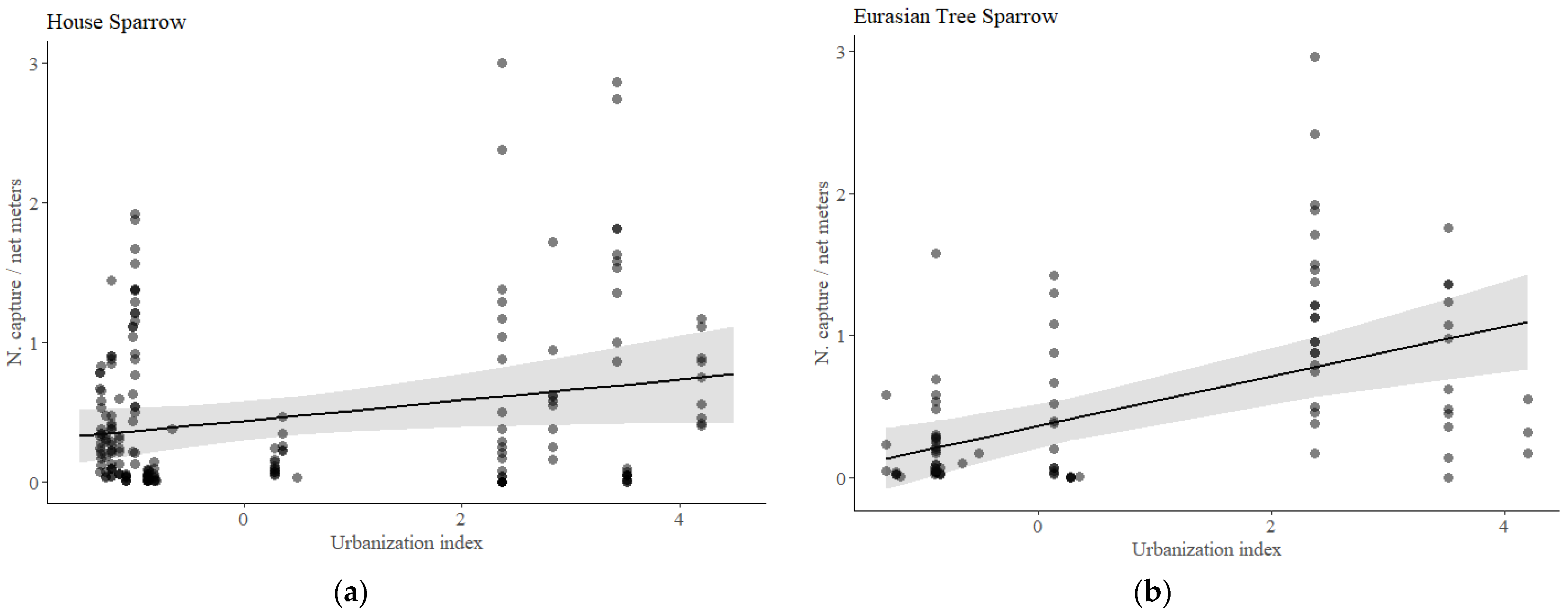
3.1. Population Trends
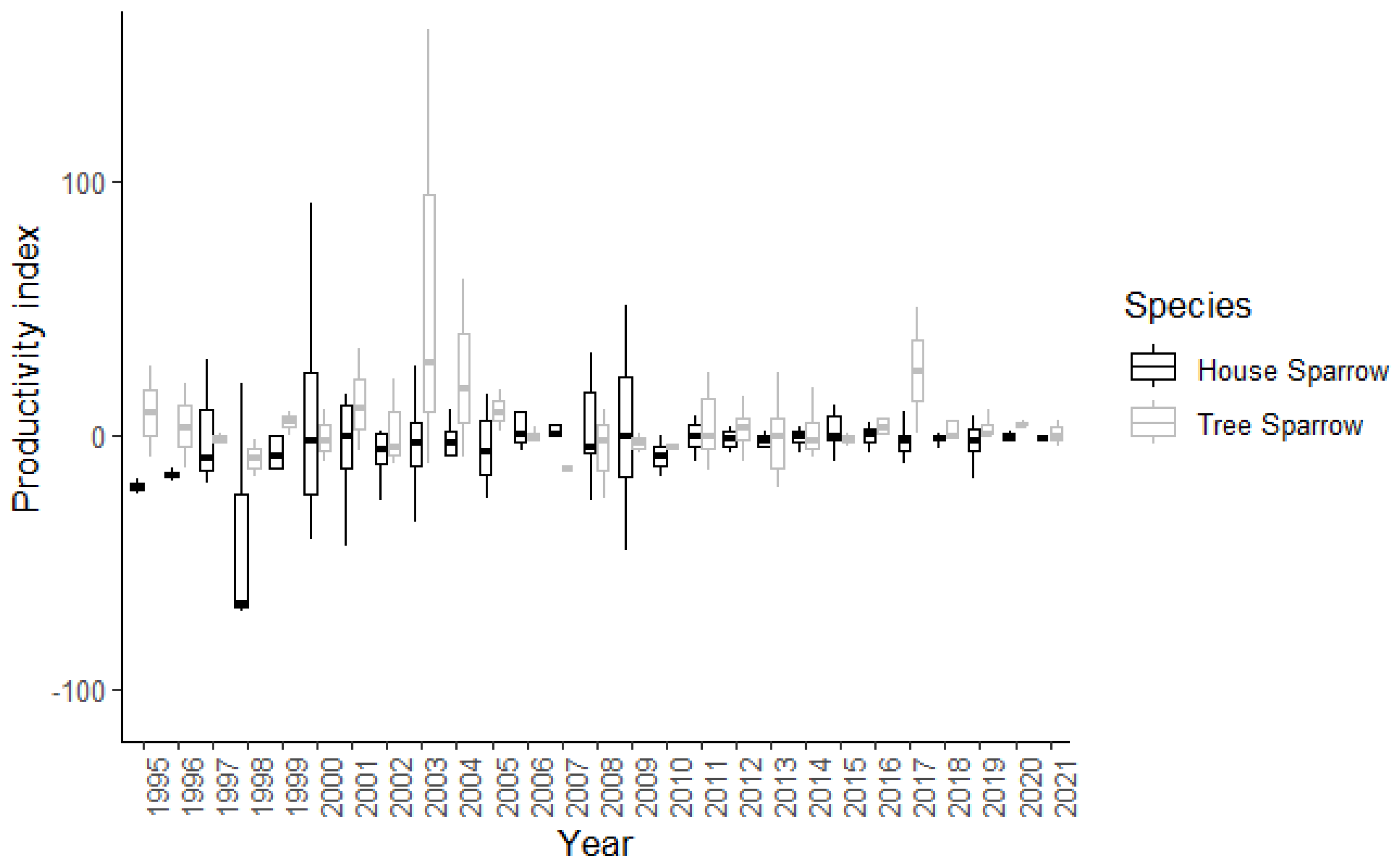
3.2. Productivity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comp. 1 | Comp. 2 | Comp. 3 | Comp. 4 | |
|---|---|---|---|---|
| Standard deviation | 1.6529 | 1.0838 | 0.3053 | 7.9799 × 10−3 |
| Proportion of Variance | 0.6830 | 0.2937 | 0.0233 | 1.5919 × 10−5 |
| Cumulative Proportion | 0.6830 | 0.9767 | 0.9999 | 1.0000 |
References
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef] [Green Version]
- Dri, G.F.; Fontana, C.S.; de Sales Dambros, C. Estimating the impacts of habitat loss induced by urbanization on bird local extinctions. Biol. Conserv. 2021, 256, 109064. [Google Scholar] [CrossRef]
- Seress, G.; Liker, A. Habitat urbanization and its effects on birds. Acta Zool. Acad. Sci. Hung. 2015, 61, 373–408. [Google Scholar] [CrossRef]
- Chace, J.F.; Walsh, J.J. Urban effects on native avifauna: A review. Landsc. Urban Plan. 2006, 74, 46–69. [Google Scholar] [CrossRef]
- Katlam, G.; Prasad, S.; Aggarwal, M.; Kumar, R. Trash on the menu: Patterns of animal visitation and foraging behaviour at garbage dumps. Curr. Sci. 2018, 115, 2322–2326. [Google Scholar] [CrossRef]
- Tryjanowski, P.; Skórka, P.; Sparks, T.H.; Biaduń, W.; Brauze, T.; Hetmański, T.; Martyka, R.; Indykiewicz, P.; Myczko, Ł.; Kunysz, P.; et al. Urban and rural habitats differ in number and type of bird feeders and in bird species consuming supplementary food. Environ. Sci. Pollut. Res. 2015, 22, 15097–15103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmon, P.; Nilsson, J.F.; Nord, A.; Bensch, S.; Isaksson, C. Urban environment shortens telomere length in nestling great tits, Parus major. Biol. Lett. 2016, 12, 20160155. [Google Scholar] [CrossRef] [Green Version]
- Toledo, A.; Andersson, M.N.; Wang, H.L.; Salmón, P.; Watson, H.; Burdge, G.C.; Isaksson, C. Fatty acid profiles of great tit (Parus major) eggs differ between urban and rural habitats, but not between coniferous and deciduous forests. Sci. Nat. 2016, 103, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clavel, J.; Julliard, R.; Devictor, V. Worldwide decline of specialist species: Toward a global functional homogenization? Front. Ecol. Environ. 2011, 9, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Anderies, J.M.; Katti, M.; Shochat, E. Living in the city: Resource availability, predation, and bird population dynamics in urban areas. J. Theor. Biol. 2007, 247, 36–49. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; Mertig, A.G.; Liu, J. Landowners and cat predation across rural-to-urban landscapes. Biol. Conserv. 2004, 115, 191–201. [Google Scholar] [CrossRef]
- Beckerman, A.P.; Boots, M.; Gaston, K.J. Urban bird declines and the fear of cats. Anim. Conserv. 2007, 10, 320–325. [Google Scholar] [CrossRef]
- Khan, M.A.; Ghouri, A.M. Environmental Pollution: Its effects on life and its remedies. Res. World J. Arts Sci. Commer. 2011, 2, 276–285. [Google Scholar]
- Khatri, N.; Tyagi, S. Influences of natural and anthropogenic factors on surface and groundwater quality in rural and urban areas. Front. Life Sci. 2014, 8, 23–39. [Google Scholar] [CrossRef]
- Blumstein, D.T.; Fernández-Juricic, E.; Zollner, P.A.; Garity, S.C. Inter-specific variation in avian responses to human disturbance. J. Appl. Ecol. 2005, 42, 943–953. [Google Scholar] [CrossRef]
- De Laet, J.; Summers-Smith, J.D. The status of the urban house sparrow Passer domesticus in north-western Europe: A review. J. Ornithol. 2007, 148 (Suppl. 2), 275–278. [Google Scholar] [CrossRef]
- Anderson, T.R. Biology of the Ubiquitous House Sparrow; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Chávez-Zichinelli, C.A.; MacGregor-Fors, I.; Rohana, P.T.; Valdéz, R.; Romano, M.C.; Schondube, J.E. Stress responses of the House Sparrow (Passer domesticus) to different urban land uses. Landsc. Urban Plan. 2010, 98, 183–189. [Google Scholar] [CrossRef]
- Ericson, P.G.P.; Tyrberg, T.; Kjellberg, A.S.; Jonsson, L.; Ullén, I. The earliest record of house sparrows (Passer domesticus) in Northern Europe. J. Archaeol. Sci. 1997, 24, 183–190. [Google Scholar] [CrossRef]
- Sætre, G.-P.; Riyahi, S.; Aliabadian, M.; Hermansen, J.S.; Hogner, S.; Olsson, U.; Gonzalez, M.F.; Sæther, S.A.; Trier, C.N.; Elgvin, T.O. Single origin of human commensalism in the house sparrow. J. Evol. Biol. 2012, 25, 788–796. [Google Scholar] [CrossRef]
- Chamberlain, D.E.; Toms, M.P.; Cleary-McHarg, R.; Banks, A.N. House Sparrow (Passer domesticus) habitat use in urbanized landscapes. J. Ornithol. 2007, 148, 453–462. [Google Scholar] [CrossRef]
- Summers-Smith, J. The Sparrows; A&C Black Publishers: London, UK, 1988. [Google Scholar]
- Brichetti, P.; Rubolini, D.; Galeotti, P.; Fasola, M. Recent declines in urban Italian Sparrow Passer (domesticus) italiae populations in northern Italy. Ibis 2008, 150, 177–181. [Google Scholar] [CrossRef]
- Siriwardena, G.M.; Baillie, S.R.; Buckland, S.T.; Fewster, R.M.; Marchant, J.H.; Wilson, J.D. Trends in the abundance of farmland birds: A quantitative comparison of smoothed Common Birds Census indices. J. Appl. Ecol. 1998, 35, 24–43. [Google Scholar] [CrossRef]
- Klok, C.; Holtkamp, R.; van Apeldoorn, R.; Visser, M.E.; Hemerik, L. Analysing Population Numbers of the House Sparrow in the Netherlands With a Matrix Model and Suggestions for Conservation Measures. Acta Biotheor. 2006, 54, 161–178. [Google Scholar] [CrossRef]
- Robinson, R.A.; Siriwardena, G.M.; Crick, H.Q.P. Size and trends of the House Sparrow Passer domesticus population in Great Britain. Ibis 2005, 147, 552–562. [Google Scholar] [CrossRef]
- Murgui, E. Gorrión común Passer domesticus. In III Atlas de las Aves en época de Reproducción en España; Molina, B., Nebreda, A., Muñoz, A.R., Seoane, J., Real, R., Bustamante, J., del Moral, J.C., Eds.; SEO/BirdLife: Madrid, Spain, 2022; Available online: https://atlasaves.seo.org/ave/gorrion-comun/ (accessed on 14 April 2022).
- Field, R.H.; Anderson, G.Q.A. Habitat use by breeding Tree Sparrows Passer montanus. Ibis 2004, 146, 60–68. [Google Scholar] [CrossRef]
- García-Navas, V.; Sanz, J.J. Yearly and Seasonal Variation of Breeding Parameters in a Declining Multi-Brooded Passerine, the Tree Sparrow. Ardea 2012, 100, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S. Habitat use of urban tree Sparrows in the process of urbanization: Beijing as a case study. Front. Biol. China 2008, 3, 308–314. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, G. Effect of urbanization on the abundance and distribution of Tree Sparrows (Passer montanus) in Beijing. Chin. Birds 2010, 1, 188–197. [Google Scholar] [CrossRef]
- Tomiałojć, L. Changes In Tree Sparrow Passer Montanus Populations From Urban Parks. Intern. Stud. Sparrows 2012, 36, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Jokimäki, J.; Suhonen, J.; Kaisanlahti-Jokimäki, M.-L. Differential Long-Term Population Responses of Two Closely Related Human-Associated Sparrow Species with Respect to Urbanization. Birds 2021, 2, 230–249. [Google Scholar] [CrossRef]
- SEO/BirdLife. Resultados del Programa Sacre de SEO/BirdLife; SEO/BirdLife: Madrid, Spain, 2013. [Google Scholar]
- BirdLife International. European Birds of Conservation Concern: Populations, Trends and National Responsibilities; Birdlife International: Cambridge, UK, 2017. [Google Scholar]
- Vepsäläinen, V.; Pakkala, T.; Tiainen, J. Population increase and aspects of colonization of the Tree Sparrow Passer montanus, and its relationships with the House Sparrow Passer domesticus, in the agricultural landscapes of Southern Finland. Ornis Fenn. 2005, 82, 117–128. [Google Scholar]
- Shaw, L.M.; Chamberlain, D.; Evans, M. The House Sparrow Passer domesticus in urban areas: Reviewing a possible link between post-decline distribution and human socioeconomic status. J. Ornithol. 2008, 149, 293–299. [Google Scholar] [CrossRef]
- Hooge, P.N.; Stanback, M.T.; Koenig, W.D. Nest-Site Selection in the Acorn Woodpecker. Auk 1999, 116, 45–54. [Google Scholar] [CrossRef]
- Tordoff, H.B.; Martell, M.S.; Redig, P.T. Midwest Peregrine Falcon Restoration, 1996 Report; Bell Museum of Natural History and the Raptor Center, University of Minnesota: St. Paul, MN, USA, 1997. [Google Scholar]
- Summers-Smith, J.D. The decline of the House Sparrow: A review. Br. Birds 2003, 96, 439–446. [Google Scholar]
- SEO/BirdLife. SEO/BirdLife 2011 Birds Monitoring Programmes; SEO/BirdLife: Madrid, Spain, 2012; pp. 14–15. [Google Scholar]
- Bermejo, A.; Palomino, D. Ringing Plan for Breeding Species Monitoring. In Programs of Monitoring of SEO/BirdLife in 2005; Escandell, V., Ed.; PASER Program; SEO/BirdLife: Madrid, Spain, 2007. [Google Scholar]
- Arizaga, J.; Crespo, A.; Iraeta, A. Sociedad de Ciencias Aranzadi. Noticias EMAN 2020, 8. Available online: http://www.aranzadi.eus/oficina-anillamiento (accessed on 14 April 2022).
- Robinson, R.A.; Julliard, R.; Saracco, J.F. Constant effort: Studying avian population processes using standardised ringing. Ringing Migr. 2009, 24, 199–204. [Google Scholar] [CrossRef]
- Kampichler, C.; van der Jeugd, H.P. Monitoring Passerine Reproduction by Constant Effort Ringing: Evaluation of the Efficiency of Trend Detection. Ardea 2011, 99, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Liker, A.; Papp, Z.; Bókony, V.; Lendvai, Á.Z. Lean birds in the city: Body size and condition of house Sparrows along the urbanization gradient. J. Anim. Ecol. 2008, 77, 789–795. [Google Scholar] [CrossRef]
- Demongin, L. Identification Guide to Birds in the Hand; Privately Published: Beauregard-Vendon, France, 2016. [Google Scholar]
- Šálek, M.; Riegert, J.; Grill, S. House Sparrows Passer domesticus and Tree Sparrows Passer montanus: Fine-Scale Distribution, Population Densities, and Habitat Selection in a Central European city. Acta Ornithol. 2015, 50, 221–232. [Google Scholar] [CrossRef]
- Węgrzynowicz, A. Importance Of Nest Sites Availability For Abundance And Changes In Number Of House- And Tree Sparrow In Warsaw. Intern. Stud. Sparrows 2012, 36, 56–65. [Google Scholar] [CrossRef]
- Summers-Smith, J.D. Densities of House Sparrows in Different Urban Habitats in a Small Town in NE England. J. Yamash. Inst. Ornithol. 2009, 40, 117–124. [Google Scholar] [CrossRef]
- Murgui, E.; Macias, A. Changes in the House Sparrow Passer domesticus population in Valencia (Spain) from 1998 to 2008. Bird Stud. 2010, 57, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L. Urban Birds Using Insects on Front Panels of Cars. Birds 2023, 4, 15–27. [Google Scholar] [CrossRef]
- Cooper, C.B.; Hochachka, W.M.; Dhondt, A.A. Contrasting natural experiments confirm competition between house finches and house Sparrows. Ecology 2007, 88, 864–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeland, D.B. Some Food Preferences and Aggressive Behavior by Monk Parakeets. Wil. Bull. 1973, 85, 332–334. [Google Scholar]
- Appelt, C.W.; Ward, L.C.; Bender, C.; Fasenella, J.; Vossen, B.J.V.; Knight, L. Examining potential relationships between exotic monk parakeets (Myiopsitta monachus) and avian communities in an urban environment. Wilson J. Ornithol. 2016, 128, 556–566. [Google Scholar] [CrossRef]
- Jessup, D.A. The welfare of feral cats and wildlife. J. Am. Vet. Med. Assoc. 2004, 225, 1377–1383. [Google Scholar] [CrossRef] [Green Version]
- Balmori, A.; Hallberg, Ö. The Urban Decline of the House Sparrow (Passer domesticus): A Possible Link with Electromagnetic Radiation. Electromagn. Biol. Med. 2007, 26, 141–151. [Google Scholar] [CrossRef]
- Summers-Smith, J.D. Is unleaded petrol a factor in urban House Sparrow decline? Br. Birds 2007, 100, 558–559. [Google Scholar]
- Cordero, P.J.; Senar, J.C. Interspecific Nest Defence in European Sparrows: Different Strategies to Deal with a Different Species of Opponent? Ornis Scand. 1990, 21, 71. [Google Scholar] [CrossRef]
- Chamberlain, D.E.; Cannon, A.R.; Toms, M.P.; Leech, D.I.; Hatchwell, B.J.; Gaston, K.J. Avian productivity in urban landscapes: A review and meta-analysis. Ibis 2009, 151, 1–18. [Google Scholar] [CrossRef]





| Station Name | Town | Latitude | Longitude | Urbanization Index | Mist Net Pool Length (m) | Data Range |
|---|---|---|---|---|---|---|
| Parque del Alamillo | Sevilla | 37°23′00 | −5°57′00 | 4.1964 | 84 | 2001–2009 |
| Real Jardín Botánico UCM | Madrid | 40°26′00 | −3°43′00 | 3.5131 | 42 | 2011–2021 |
| La Murtera | Oda | 39°58′00 | −1°50′00 | 3.4190 | 43 | 1999–2009 |
| Embalse la Portiña | Talavera de la Reina | 39°57′00 | −4°50′00 | 2.8300 | 69 | 2000–2007 |
| Parque del Oeste | Madrid | 40°28′00 | −3°41′00 | 2.3658 | 12 | 1995–2016 |
| Barrutibasoko Lezkadia | Gautegiz-Arteaga | 43°19′0″ | −2°40′0″ | 0.4869 | 120 | 2010–2019 |
| La Tejera | Nalda | 42°20′0″ | −2°29′0″ | 0.3534 | 120 | 2015–2020 |
| Jaizubia | Hondarribia | 43°21′0″ | −1°49′0″ | 0.2795 | 216 | 2010–2020 |
| Laguna Manjavacas | Mota del Cuervo | 39°27′00 | −2°52′00 | 0.1331 | 60 | 2011–2016 |
| Barajas | Paracuellos del Jarama | 40°28′11″ | −3°31′37″ | −0.5088 | 78 | 2019 |
| Río Guadalix—Embalse Pedrezuela | Guadalix de la Sierra | 40°47′9″ | −3°40′42″ | −0.6537 | 90 | 2019–2020 |
| Mejana del Casetón | Sobradiel-Utebo | 41°44′0″ | −1°1′0″ | −0.7962 | 84 | 2017–2020 |
| Ubagua | Riezu-Muez | 42°45′13″ | −1°56′33″ | −0.8099 | 126 | 2019–2020 |
| Loza | Loza | 42°50′0″ | −1°43′0″ | −0.8176 | 156 | 2012–2021 |
| La Nava | Fuentes de Nava | 42°4′0″ | −4°45′0″ | −0.8440 | 60 | 2013–2020 |
| Las Cañas | Viana | 42°29′0″ | −2°24′0″ | −0.8694 | 120 | 2013–2021 |
| Las Minas | San Martín de la Vega | 40°14′00 | −3°33′00 | −0.8802 | 138 | 1995–2021 |
| Autillos | Pina de Ebro | 41°30′0″ | −0°23′0″′ | −0.8892 | 60 | 2017–2019 |
| Cortijo de Auta | Riogordo | 36°55′00 | −4°16′00 | −0.9939 | 52 | 1997–2007 2019–2020 |
| Río Salado | Almargen | 37°1′00 | −5°10′00 | −1.0148 | 27 | 2000–2007 |
| Motondo | Orio | 43°16′0″ | −2°17′0″ | −1.0825 | 174 | 2010–2020 |
| Arroyo Samburiel | Manzanares el Real | 40°45′00 | −3°52′00 | −1.14033 | 67 | 2002–2009 |
| Soto del Rincón Falso | Pastriz | 41°36′0″ | −0°45′0″′ | −1.1800 | 72 | 2017–2019 |
| La Higueruela | Santa Olalla | 40°00′00 | −4°25′00 | −1.2186 | 48 | 1995–2009 |
| Santa Eulalia | Arnedo | 42°13′0″ | −2°12′0″ | −1.2621 | 120 | 2012–2020 |
| La Veguilla | Córdoba | 37°51′00 | −4°42′00 | −1.3021 | 60 | 2002–2009 |
| Dehesa de Castrillo | Toro | 41°28′00 | −5°26′00 | −1.3142 | 54 | 1999–2004 |
| HOUSE SPARROW | Estimate | SE | F | Df | p | |
| Population trends | Intercept | 0.9550 | 0.2846 | 11.2266 | 1 | 0.0010 |
| urban index | 0.0738 | 0.0378 | 3.8047 | 1 | 0.0683 | |
| year | −0.0171 | 0.3236 | 2.9104 | 26 | <0.0001 | |
| Productivity | Intercept | −15.9132 | 12.4865 | 1.6213 | 1 | 0.2047 |
| urban index | 0.0941 | 2.3035 | 0.0942 | 1 | 0.7627 | |
| year | 4.5000 | 16.1885 | 1.2715 | 26 | 0.1872 | |
| TREE SPARROW | Estimate | SE | F | Df | p | |
| Population trends | Intercept | 0.7432 | 0.3013 | 5.9202 | 1 | 0.0177 |
| urban index | 0.1749 | 0.0412 | 16.5448 | 1 | 0.0043 | |
| year | -0.1950 | 0.4059 | 2.3302 | 26 | 0.0037 | |
| Productivity | Intercept | 11.3803 | 14.1087 | 0.6329 | 1 | 0.4291 |
| urban index | 0.2997 | 1.9202 | 0.0224 | 1 | 0.8851 | |
| year | 3.4740 | 17.4493 | 1.3893 | 26 | 0.1479 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos-Elvira, E.; Banda, E.; Arizaga, J.; Martín, D.; Aguirre, J.I. Long-Term Population Trends of House Sparrow and Eurasian Tree Sparrow in Spain. Birds 2023, 4, 159-170. https://doi.org/10.3390/birds4020013
Ramos-Elvira E, Banda E, Arizaga J, Martín D, Aguirre JI. Long-Term Population Trends of House Sparrow and Eurasian Tree Sparrow in Spain. Birds. 2023; 4(2):159-170. https://doi.org/10.3390/birds4020013
Chicago/Turabian StyleRamos-Elvira, Elena, Eva Banda, Juan Arizaga, David Martín, and José I. Aguirre. 2023. "Long-Term Population Trends of House Sparrow and Eurasian Tree Sparrow in Spain" Birds 4, no. 2: 159-170. https://doi.org/10.3390/birds4020013
APA StyleRamos-Elvira, E., Banda, E., Arizaga, J., Martín, D., & Aguirre, J. I. (2023). Long-Term Population Trends of House Sparrow and Eurasian Tree Sparrow in Spain. Birds, 4(2), 159-170. https://doi.org/10.3390/birds4020013