
A Bird in the Hand Is Worth Two in the Bush: Bird Visibility as a Predictor of the Perception of Birds by Humans


Abstract
:Simple Summary
Abstract
1. Introduction
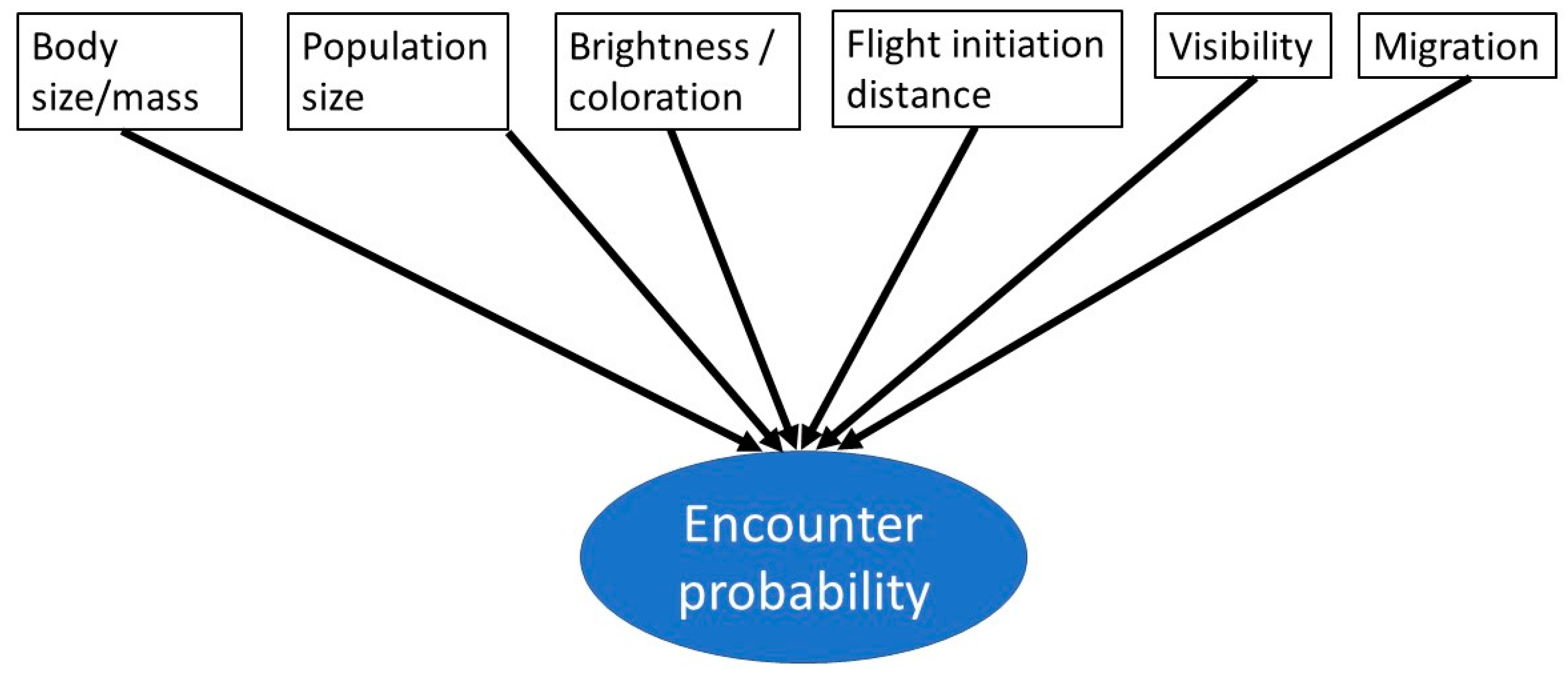
1.1. Why Encounter Probability Matters
1.2. The Theoretical Framing of Bird Encounter Probability
1.3. The Importance of Behavior on Visibility
1.4. Goals of the Study
2. Materials and Methods
2.1. Study Area
2.2. Fieldwork
2.3. Validation of the Observational Data with an Expert Assessment
2.4. Additional Species Traits
2.5. Statistical Analysis
2.5.1. Fieldwork
2.5.2. Expert Assessment
3. Results
4. Discussion
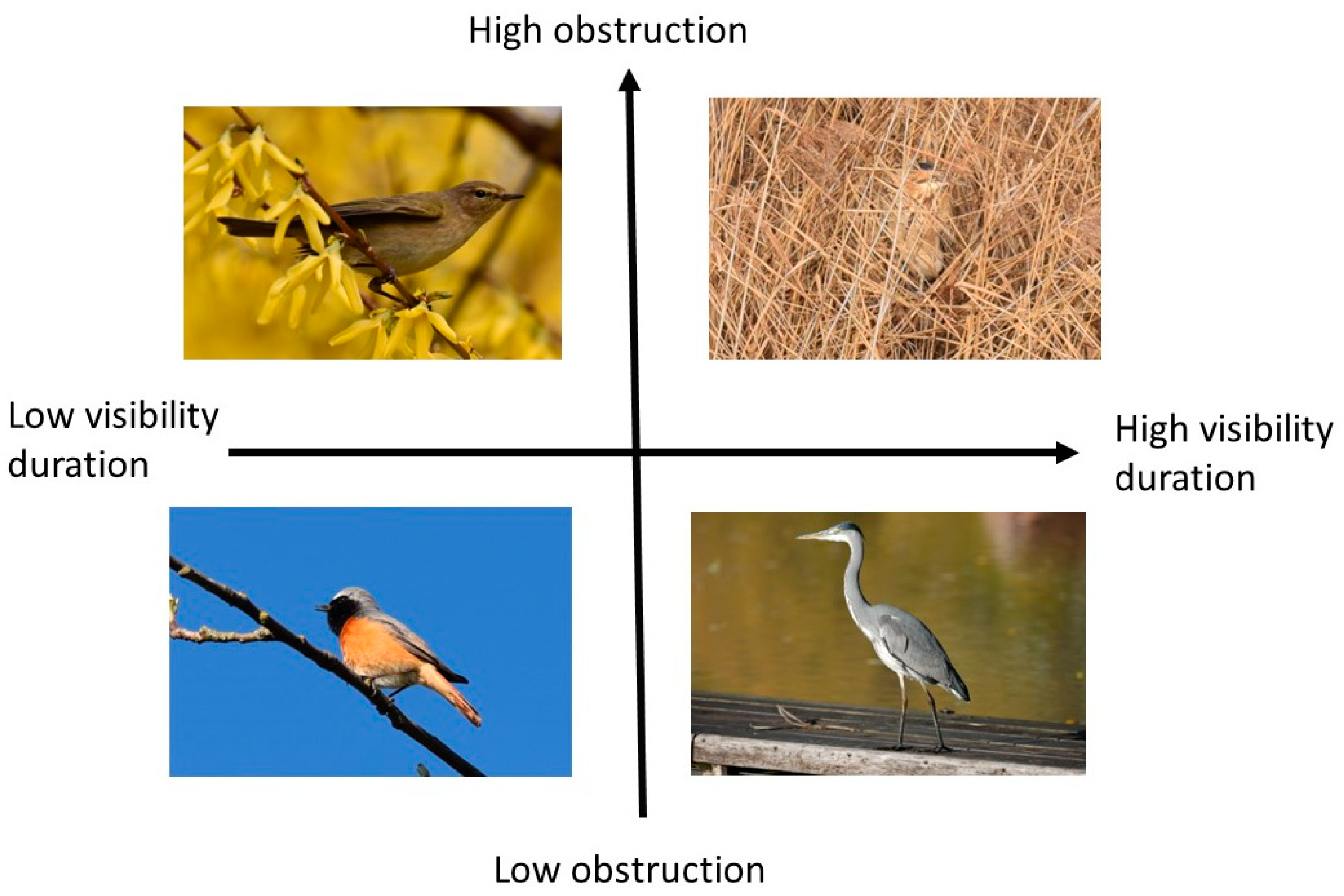
4.1. Confounders of Visibility Duration
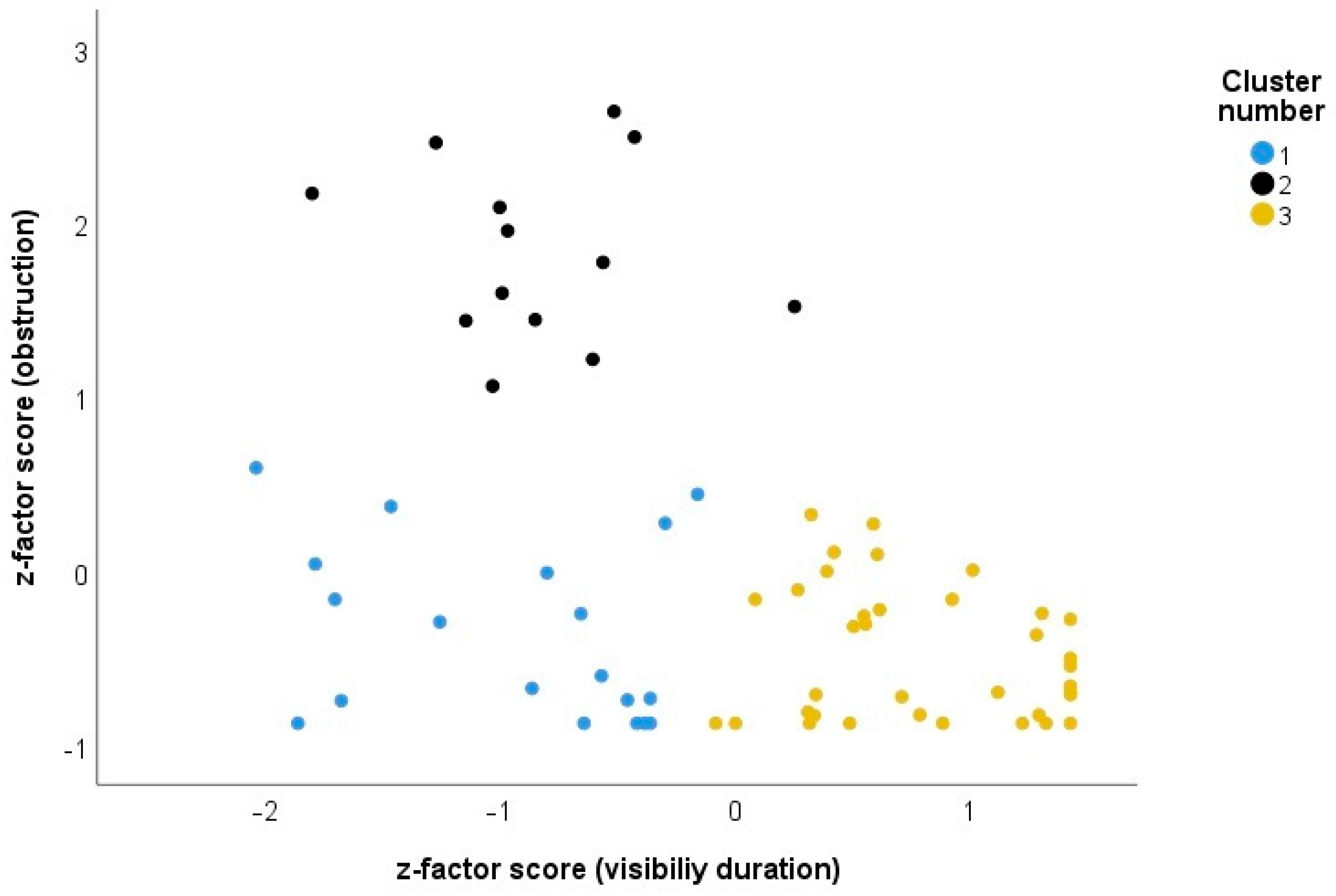
4.2. Clustering of the Species
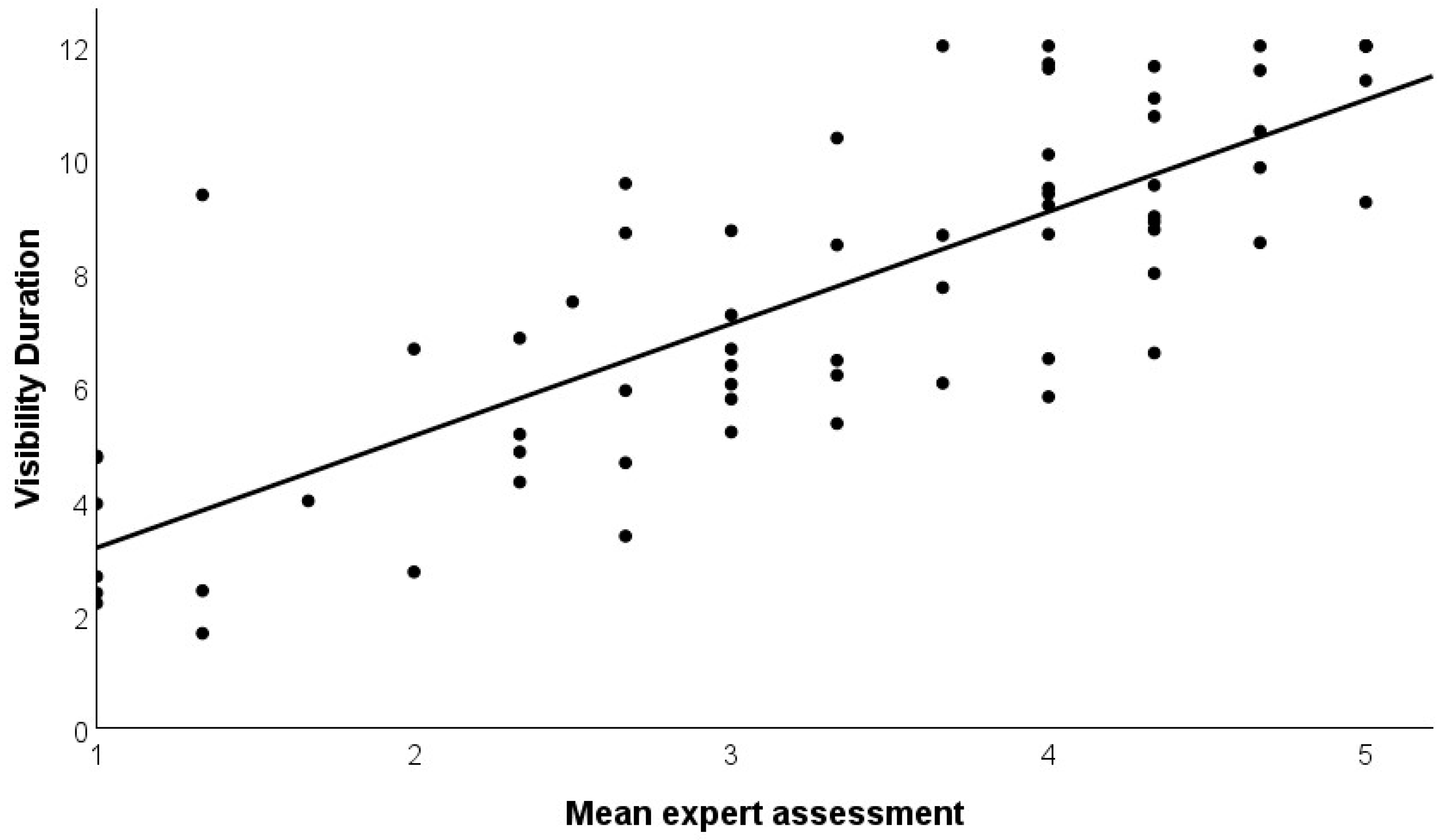
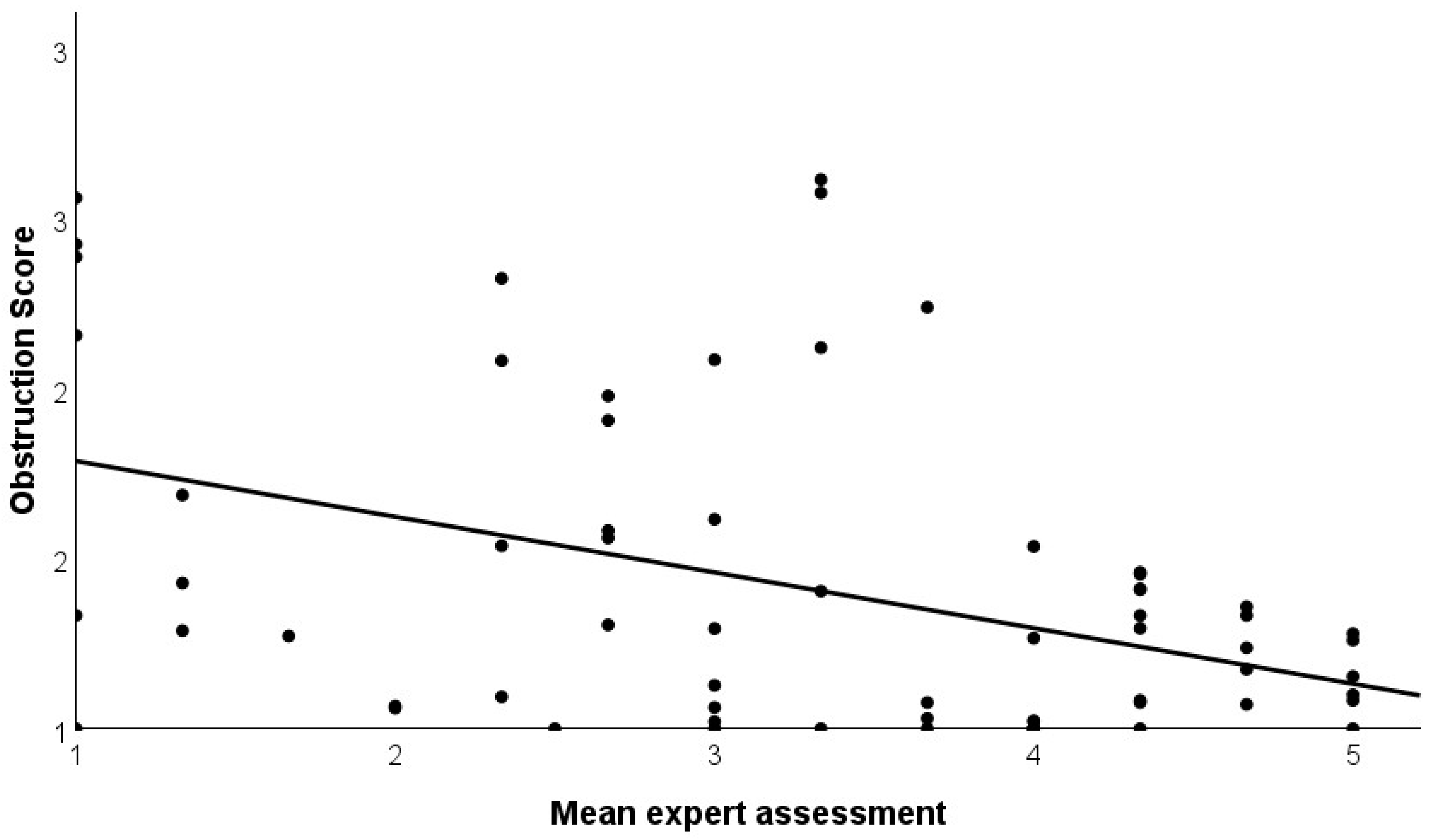
4.3. Validation with Expert Ratings
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

| Scientific Name | Visibility | N | SD | Obstruction | N | SD |
|---|---|---|---|---|---|---|
| Acrocephalus arundinaceus | 2.67 | 12 | 2.67 | 1.33 | 9 | 0.56 |
| Aegithalos caudatus | 4.86 | 14 | 3.01 | 2.33 | 13 | 1.05 |
| Alauda arvensis | 4.00 | 12 | 4.00 | 1.27 | 11 | 0.47 |
| Alcedo atthis | 2.75 | 12 | 2.30 | 1.06 | 11 | 0.20 |
| Alopochen aegyptiacus | 12.00 | 13 | 0.00 | 1.15 | 13 | 0.38 |
| Anas platyrhynchos | 12.00 | 21 | 0.00 | 1.17 | 21 | 0.45 |
| Anser anser | 12.00 | 14 | 0.00 | 1.28 | 14 | 0.43 |
| Apus apus | 8.69 | 16 | 4.06 | 1.00 | 16 | 0.00 |
| Ardea cinerea | 11.39 | 18 | 2.15 | 1.00 | 18 | 0.00 |
| Aythya fuligula | 12.00 | 16 | 0.00 | 1.00 | 16 | 0.00 |
| Buteo buteo | 7.75 | 12 | 4.12 | 1.00 | 12 | 0.00 |
| Carduelis carduelis | 5.79 | 14 | 3.87 | 1.29 | 14 | 0.82 |
| Chloris chloris | 7.27 | 11 | 3.61 | 1.62 | 11 | 0.51 |
| Choicocephalus ridibundus | 12.00 | 13 | 0.00 | 1.08 | 13 | 0.28 |
| Ciconia ciconia | 11.64 | 22 | 1.33 | 1.30 | 22 | 0.88 |
| Coleus monedula | 8.91 | 11 | 3.42 | 1.41 | 11 | 0.48 |
| Columba livia f. domestica | 9.86 | 14 | 3.53 | 1.07 | 14 | 0.27 |
| Columba oenas | 6.60 | 10 | 5.72 | 1.00 | 9 | 0.00 |
| Columba palumbus | 9.55 | 11 | 4.01 | 1.45 | 11 | 0.69 |
| Corvus cornix | 10.76 | 17 | 2.77 | 1.41 | 17 | 0.78 |
| Corvus corone | 10.50 | 16 | 2.34 | 1.33 | 16 | 0.71 |
| Cuculus canorus | 2.20 | 10 | 3.65 | 1.00 | 7 | 0.00 |
| Cyanistes caeruleus | 6.21 | 14 | 3.98 | 2.65 | 14 | 0.77 |
| Cygnus olor | 12.00 | 15 | 0.00 | 1.08 | 15 | 0.18 |
| Delichon urbicum | 6.50 | 12 | 4.66 | 1.00 | 12 | 0.00 |
| Dendrocopus major | 4.67 | 18 | 3.34 | 1.91 | 18 | 0.89 |
| Emberiza citrinella | 8.71 | 14 | 4.05 | 1.56 | 14 | 0.95 |
| Erithacus rubecula | 5.94 | 17 | 3.93 | 1.98 | 15 | 1.00 |
| Falco tinnunculus | 8.67 | 12 | 3.99 | 1.03 | 12 | 0.10 |
| Fringilla coelebs | 8.50 | 12 | 3.50 | 2.12 | 12 | 1.06 |
| Fulica atra | 11.57 | 14 | 1.60 | 1.24 | 14 | 0.62 |
| Gallinula chloropus | 5.36 | 11 | 3.85 | 1.40 | 9 | 0.71 |
| Garrulus glandarius | 5.21 | 14 | 4.69 | 2.09 | 14 | 0.95 |
| Hirundo rustica | 5.83 | 12 | 4.00 | 1.00 | 12 | 0.00 |
| Lanius collurio | 6.67 | 15 | 4.39 | 1.07 | 15 | 0.26 |
| Linaria cannabina | 5.17 | 12 | 2.89 | 1.09 | 12 | 0.29 |
| Luscinia megarhynchos | 1.67 | 12 | 1.78 | 1.69 | 8 | 0.88 |
| Mareca strepera | 11.69 | 13 | 0.75 | 1.00 | 13 | 0.00 |
| Merops apiaster | 9.40 | 15 | 3.23 | 1.27 | 15 | 0.46 |
| Milvus migrans | 9.20 | 10 | 3.55 | 1.00 | 10 | 0.00 |
| Milvus milvus | 8.77 | 13 | 3.35 | 1.08 | 13 | 0.28 |
| Motacilla alba | 9.00 | 13 | 2.31 | 1.46 | 13 | 0.85 |
| Motacilla cinerea | 6.67 | 12 | 4.33 | 1.00 | 12 | 0.00 |
| Motacilla flava | 6.38 | 16 | 4.37 | 1.06 | 16 | 0.17 |
| Netta rufina | 11.60 | 15 | 0.83 | 1.02 | 15 | 0.06 |
| Parus major | 6.07 | 15 | 3.49 | 2.24 | 15 | 0.86 |
| Passer domesticus | 8.54 | 13 | 3.69 | 1.36 | 13 | 0.66 |
| Passer montanus | 6.47 | 15 | 2.95 | 2.58 | 15 | 1.02 |
| Phalacrocorax carbo | 11.08 | 12 | 3.18 | 1.08 | 12 | 0.29 |
| Phoenicurus ochruros | 6.05 | 19 | 3.66 | 1.13 | 19 | 0.42 |
| Phoenicurus phoenicurus | 6.86 | 14 | 3.80 | 1.54 | 14 | 0.95 |
| Phylloscopus collybita | 4.76 | 17 | 3.68 | 2.39 | 16 | 0.77 |
| Pica pica | 9.50 | 12 | 2.94 | 1.54 | 12 | 0.85 |
| Picus viridis | 3.38 | 13 | 3.73 | 1.58 | 12 | 1.16 |
| Podiceps cristatus | 10.09 | 11 | 2.95 | 1.02 | 11 | 0.08 |
| Saxicola rubicola | 7.50 | 14 | 4.03 | 1.00 | 14 | 0.00 |
| Sitta europaea | 4.33 | 12 | 3.55 | 2.08 | 12 | 0.81 |
| Sturnus vulgaris | 8.00 | 15 | 3.55 | 1.33 | 15 | 0.63 |
| Sylvia atricapilla | 3.95 | 20 | 3.25 | 2.56 | 18 | 1.05 |
| Sylvia borin | 2.38 | 13 | 1.94 | 2.43 | 9 | 1.20 |
| Sylvia curruca | 4.79 | 14 | 4.04 | 2.16 | 11 | 1.57 |
| Tachybaptus ruficollis | 10.38 | 13 | 3.20 | 1.00 | 13 | 0.00 |
| Tadorna ferruginea | 12.00 | 10 | 0.00 | 1.10 | 10 | 0.32 |
| Troglodytes troglodytes | 2.42 | 12 | 3.78 | 1.43 | 7 | 0.79 |
| Turdus merula | 9.25 | 16 | 3.11 | 1.26 | 16 | 0.58 |
| Turdus philomelos | 9.38 | 13 | 3.60 | 1.29 | 13 | 0.45 |
| Turdus pilaris | 8.75 | 12 | 4.67 | 1.02 | 12 | 0.07 |
| Vanellus vanellus | 9.58 | 12 | 3.87 | 1.31 | 12 | 0.44 |
References
- Champness, B.S.; Fitzsimons, J.A.; Kendal, D.; Palmer, G.C. Perceptions of Birds by Urban Residents in an Australian Regional City and Implications for Conservation. Birds 2023, 4, 262–276. [Google Scholar] [CrossRef]
- Fisher, J.C.; Dallimer, M.; Irvine, K.N.; Aizlewood, S.G.; Austen, G.E.; Fish, R.D.; King, P.M. Human well-being responses to species’ traits. Nat. Sustain. 2023, 6, 1219–1227. [Google Scholar] [CrossRef]
- Randler, C.; Friedrich, S.; Koch, S. Psychological restoration, place attachment and satisfaction in birders and non-birding visitors. J. Outdoor Recreat. Tour. 2023, 44, 100679. [Google Scholar] [CrossRef]
- Randler, C.; Staller, N.; Kalb, N.; Tryjanowski, P. Charismatic Species and Birdwatching: Advanced Birders Prefer Small, Shy, Dull, and Rare Species. Anthrozoös 2023, 36, 427–445. [Google Scholar] [CrossRef]
- Tubelis, D.P. Spatiotemporal Distribution of Photographic Records of Brazilian Birds Available in the WikiAves Citizen Science Database. Birds 2023, 4, 28–45. [Google Scholar] [CrossRef]
- Ducarme, F.; Luque, G.M.; Courchamp, F. What are “charismatic species” for conservation biologists. BioSci. Master Rev. 2013, 10, 1–8. [Google Scholar]
- Sergio, F.; Newton, I.A.N.; Marchesi, L.; Pedrini, P. Ecologically justified charisma: Preservation of top predators delivers biodiversity conservation. J. Appl. Ecol. 2006, 43, 1049–1055. [Google Scholar] [CrossRef]
- Whelan, C.J.; Wenny, D.G.; Marquis, R.J. Ecosystem services provided by birds. Ann. N. Y. Acad. Sci. 2008, 1134, 25–60. [Google Scholar] [CrossRef]
- Whelan, C.J.; Şekercioğlu, Ç.H.; Wenny, D.G. Why birds matter: From economic ornithology to ecosystem services. J. Ornithol. 2015, 156, 227–238. [Google Scholar] [CrossRef]
- McGinlay, J.; Parsons, D.J.; Morris, J.; Hubatova, M.; Graves, A.; Bradbury, R.B.; Bullock, J.M. Do charismatic species groups generate more cultural ecosystem service benefits? Ecosyst. Serv. 2017, 27, 15–24. [Google Scholar] [CrossRef]
- Ratcliffe, E.; Gatersleben, B.; Sowden, P.T. Bird sounds and their contributions to perceived attention restoration and stress recovery. J. Environ. Psychol. 2013, 36, 221–228. [Google Scholar] [CrossRef]
- Leong, R.A.; Fung, T.K.; Sachidhanandam, U.; Drillet, Z.; Edwards, P.J.; Richards, D.R. Use of structural equation modeling to explore influences on perceptions of ecosystem services and disservices attributed to birds in Singapore. Ecosyst. Serv. 2020, 46, 101211. [Google Scholar] [CrossRef]
- Tan, X.; Yan, P.; Liu, Z.; Qin, H.; Jiang, A. Demographics, behaviours, and preferences of birdwatchers and their implications for avitourism and avian conservation: A case study of birding in Nonggang, Southern China. Glob. Ecol. Conserv. 2023, 46, e02552. [Google Scholar] [CrossRef]
- Morrison, C.A.; Auniņš, A.; Benkő, Z.; Brotons, L.; Chodkiewicz, T.; Chylarecki, P.; Escandell, V.; Eskildsen, D.P.; Gamero, A.; Herrando, S.; et al. Bird population declines and species turnover are changing the acoustic properties of spring soundscapes. Nat. Commun. 2021, 12, 6217. [Google Scholar] [CrossRef] [PubMed]
- Soga, M.; Gaston, K.J. The ecology of human–nature interactions. Proc. R. Soc. B 2020, 287, 20191882. [Google Scholar] [CrossRef] [PubMed]
- Houlden, V.; Jani, A.; Hong, A. Is biodiversity of greenspace important for human health and wellbeing? A bibliometric analysis and systematic literature review. Urban For. Urban Green. 2021, 66, 127385. [Google Scholar] [CrossRef]
- Johansson, M.; Flykt, A.; Frank, J.; Hartig, T. Appraisals of wildlife during restorative opportunities in local natural settings. Front. Environ. Sci. 2021, 9, 635757. [Google Scholar] [CrossRef]
- Randler, C.; Höllwarth, A.; Schaal, S. Urban park visitors and their knowledge of animal species. Anthrozoös 2007, 20, 65–74. [Google Scholar] [CrossRef]
- Blumstein, D.T. Developing an evolutionary ecology of fear: How life history and natural history traits affect disturbance tolerance in birds. Anim. Behaviour. 2006, 71, 389–399. [Google Scholar] [CrossRef]
- Bateson, M.; Martin, P. Measuring Behaviour: An Introductory Guide; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Boyer, J.S.; Hass, L.L.; Lurie, M.H.; Blumstein, D.T. Effect of visibility on time allocation and escape decisions in crimson rosellas. Aust. J. Zool. 2006, 54, 363–367. [Google Scholar] [CrossRef]
- Randler, C. Risk assessment by crow phenotypes in a hybrid zone. J. Ethol. 2008, 26, 309–316. [Google Scholar] [CrossRef]
- Bauer, H.G.; Bezzel, E.; Fiedler, W. Das Kompendium der Vögel Mitteleuropas. Alles über Biologie, Gefährdung und Schutz; Aula Verlag: Wiebelsheim, Germany, 2005. [Google Scholar]
- Randler, C. Frequency of bird hybrids: Does detectability make all the difference? J. Ornithol. 2004, 145, 123–128. [Google Scholar] [CrossRef]
- Randler, C. Elite Recreation Specialization and Motivations among Birdwatchers: The Case of Club 300 Members. Int. J. Sociol. Leis. 2023, 6, 209–223. [Google Scholar] [CrossRef]
- Ladle, R.J.; Jepson, P.; Correia, R.A.; Malhado, A.C. A culturomics approach to quantifying the salience of species on the global internet. People Nat. 2019, 1, 524–532. [Google Scholar] [CrossRef]
- Tobias, J.A.; Sheard, C.; Pigot, A.L.; Devenish, A.J.; Yang, J.; Sayol, F.; Neate-Clegg, M.H.C.; Alioravainen, N.; Weeks, T.L.; Barber, R.A.; et al. AVONET: Morphological, ecological and geographical data for all birds. Ecol. Lett. 2022, 25, 581–597. [Google Scholar] [CrossRef]
- Fuller, R.A.; Irvine, K.N.; Devine-Wright, P.; Warren, P.H.; Gaston, K.J. Psychological benefits of greenspace increase with biodiversity. Biol. Lett. 2007, 3, 390–394. [Google Scholar] [CrossRef]
- Møller, A.P.; Nielsen, J.T.; Garamszegi, L.Z. Song post exposure, song features, and predation risk. Behav. Ecol. 2006, 17, 155–163. [Google Scholar] [CrossRef]
- Campos, D.P.; Bander, L.A.; Raksi, A.; Blumstein, D.T. Perch exposure and predation risk: A comparative study in passerines. Acta Ethol. 2009, 12, 93–98. [Google Scholar] [CrossRef]
- Conway, C.J.; Gibbs, J.P. Summary of Intrinsic and Extrinsic Factors Affecting Detection Probability of Marsh Birds. Wetlands 2011, 31, 403–411. [Google Scholar] [CrossRef]
- Tozer, D.C.; Drake, K.L.; Falconer, C.M. Modeling detection probability to improve marsh bird surveys in southern Canada and the Great Lakes states. Avian Conserv. Ecol. 2016, 11, 3. [Google Scholar] [CrossRef]
- Härtel, T.; Vanhöfen, J.; Randler, C. Selection of Indicator Bird Species as a Baseline for Knowledge Assessment in Biodiversity Survey Studies. Animals 2023, 13, 2230. [Google Scholar] [CrossRef] [PubMed]
- Kułaga, K.; Budka, M. Bird species detection by an observer and an autonomous sound recorder in two different environments: Forest and farmland. PLoS ONE 2019, 14, e0211970. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.; Robinson, W.D. Mobbing behavior of songbirds in response to calls of an ambush-predator, the Northern Pygmy-Owl. Front. Ecol. Evol. 2023, 11, 1092323. [Google Scholar] [CrossRef]
- Randler, C.; Vanhöfen, J.; Härtel, T.; Neunhoeffer, F.; Engeser, C.; Fischer, C. Psychological restoration depends on curiosity, motivation, and species richness during a guided bird walk in a suburban blue space. Front. Psychol. 2023, 14, 1176202. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




| Dependent Variable | Measurement | Question |
|---|---|---|
| Visibility duration | 3 min, split into twelve 15 s periods | How long is the individual visible to the observer? |
| Obstruction | Minimum percentage of obstruction, 3 min, split into twelve 15 s periods | To what extent is the individual obstructed by vegetation? |
| Predictor variable | ||
| Species | Bird species | Do bird species differ in visibility/obstruction? |
| Sex | Male/female in dichromatic species | Are there sex differences? |
| Flock | Flocking yes/no | Is the bird a member of a flock/group or single? |
| Detection mode | Acoustic/visual | Was the bird discovered by acoustic or visual cues? |
| Observation distance | In meters | Does the distance between observer and bird influence visibility? |
| Behavior | In flight or not | Was the bird in flight or not? |
| Source of Variance | Type III Sum of Squares | df | Mean of Squares | F | Sig. | Partial Eta-Squared |
|---|---|---|---|---|---|---|
| Sex | 0.011 | 1 | 0.011 | 0.232 | 0.631 | 0.002 |
| Flock | 0.036 | 1 | 0.036 | 0.750 | 0.388 | 0.006 |
| Detection mode | 0.033 | 1 | 0.033 | 0.683 | 0.410 | 0.005 |
| Species | 5.109 | 33 | 0.155 | 3.193 | <0.001 *** | 0.440 |
| Observation distance | 0.305 | 1 | 0.305 | 6.296 | 0.013 ** | 0.045 |
| Source of Variance | Type III Sum of Squares | df | Mean of Squares | F | Sig. | Partial Eta-Squared |
|---|---|---|---|---|---|---|
| Sex | 0.003 | 1 | 0.003 | 0.004 | 0.949 | <0.009 |
| Flock | 0.039 | 1 | 0.039 | 0.057 | 0.812 | <0.009 |
| Detection mode | 3.926 | 1 | 3.926 | 5.769 | 0.018 ** | 0.041 |
| Species | 47.579 | 33 | 1.442 | 2.118 | 0.001 *** | 0.343 |
| Observation distance | 4.54 | 1 | 4.540 | 6.670 | 0.011 ** | 0.047 |
| Mean Expert Assessment | Visibility Duration | Obstruction | ||
|---|---|---|---|---|
| Coloration assessment by experts | Spearman rho | −0.116 | −0.176 | 0.085 |
| P | 0.345 | 0.152 | 0.489 | |
| N | 68 | 68 | 68 | |
| Assessment of brightness (by lay people) | Spearman rho | −0.146 | −0.205 | 0.249 |
| P | 0.376 | 0.210 | 0.126 | |
| N | 39 | 39 | 39 | |
| Body mass | Spearman rho | 0.701 | 0.757 | −0.412 |
| P | <0.001 | <0.001 | <0.001 | |
| N | 68 | 68 | 68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Randler, C.; Härtel, T.; Kalb, N.; Vanhöfen, J. A Bird in the Hand Is Worth Two in the Bush: Bird Visibility as a Predictor of the Perception of Birds by Humans. Birds 2024, 5, 24-37. https://doi.org/10.3390/birds5010002
Randler C, Härtel T, Kalb N, Vanhöfen J. A Bird in the Hand Is Worth Two in the Bush: Bird Visibility as a Predictor of the Perception of Birds by Humans. Birds. 2024; 5(1):24-37. https://doi.org/10.3390/birds5010002
Chicago/Turabian StyleRandler, Christoph, Talia Härtel, Nadine Kalb, and Janina Vanhöfen. 2024. "A Bird in the Hand Is Worth Two in the Bush: Bird Visibility as a Predictor of the Perception of Birds by Humans" Birds 5, no. 1: 24-37. https://doi.org/10.3390/birds5010002
APA StyleRandler, C., Härtel, T., Kalb, N., & Vanhöfen, J. (2024). A Bird in the Hand Is Worth Two in the Bush: Bird Visibility as a Predictor of the Perception of Birds by Humans. Birds, 5(1), 24-37. https://doi.org/10.3390/birds5010002