
In Vitro Anti-HIV-1 Activity of Chitosan Oligomers N-Conjugated with Asparagine and Glutamine


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
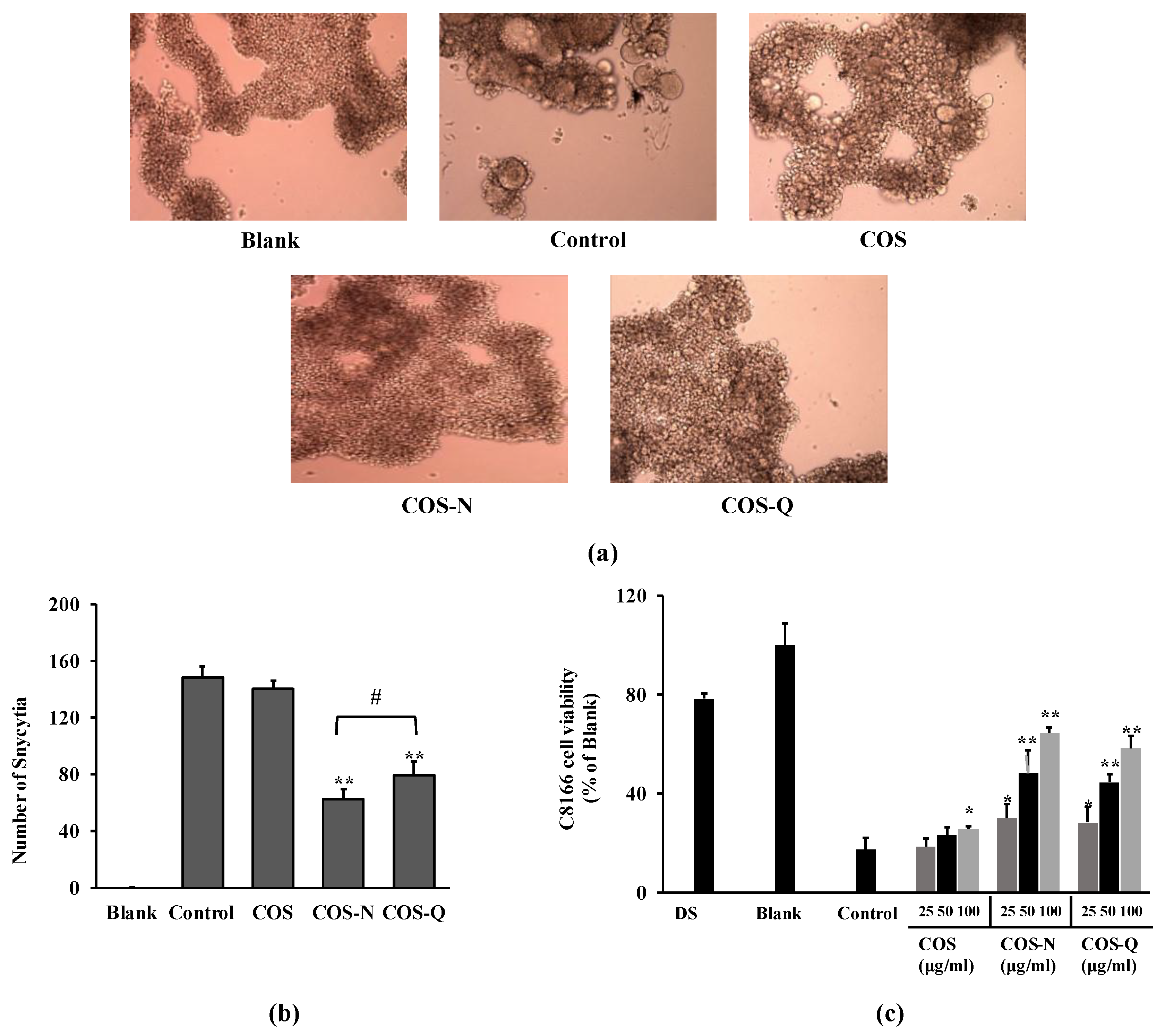
2.1. Inhibition of HIV-1 Infection
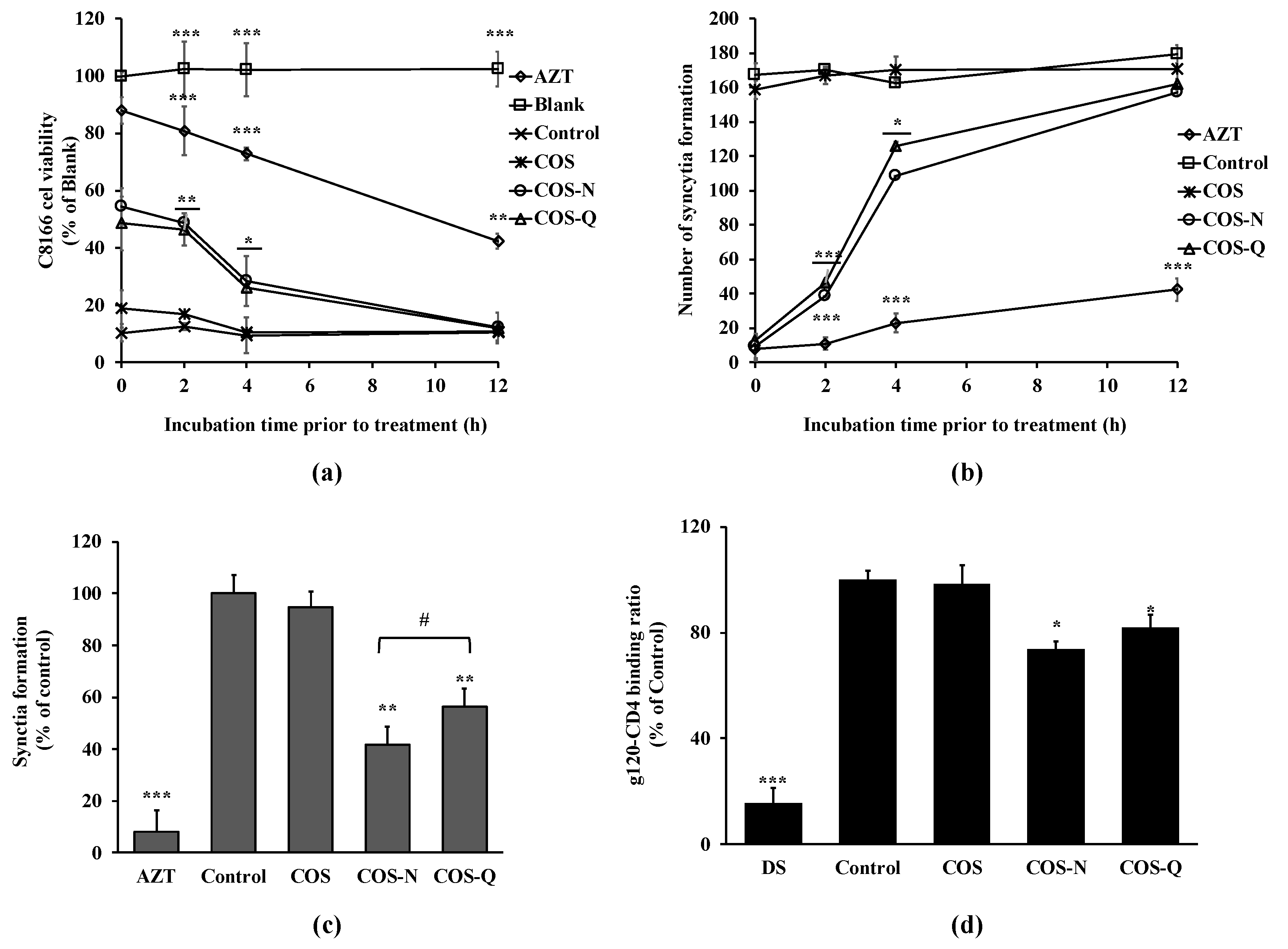
2.1.1. Inhibition of Syncytia Formation and HIV-Induced Lytic Effects
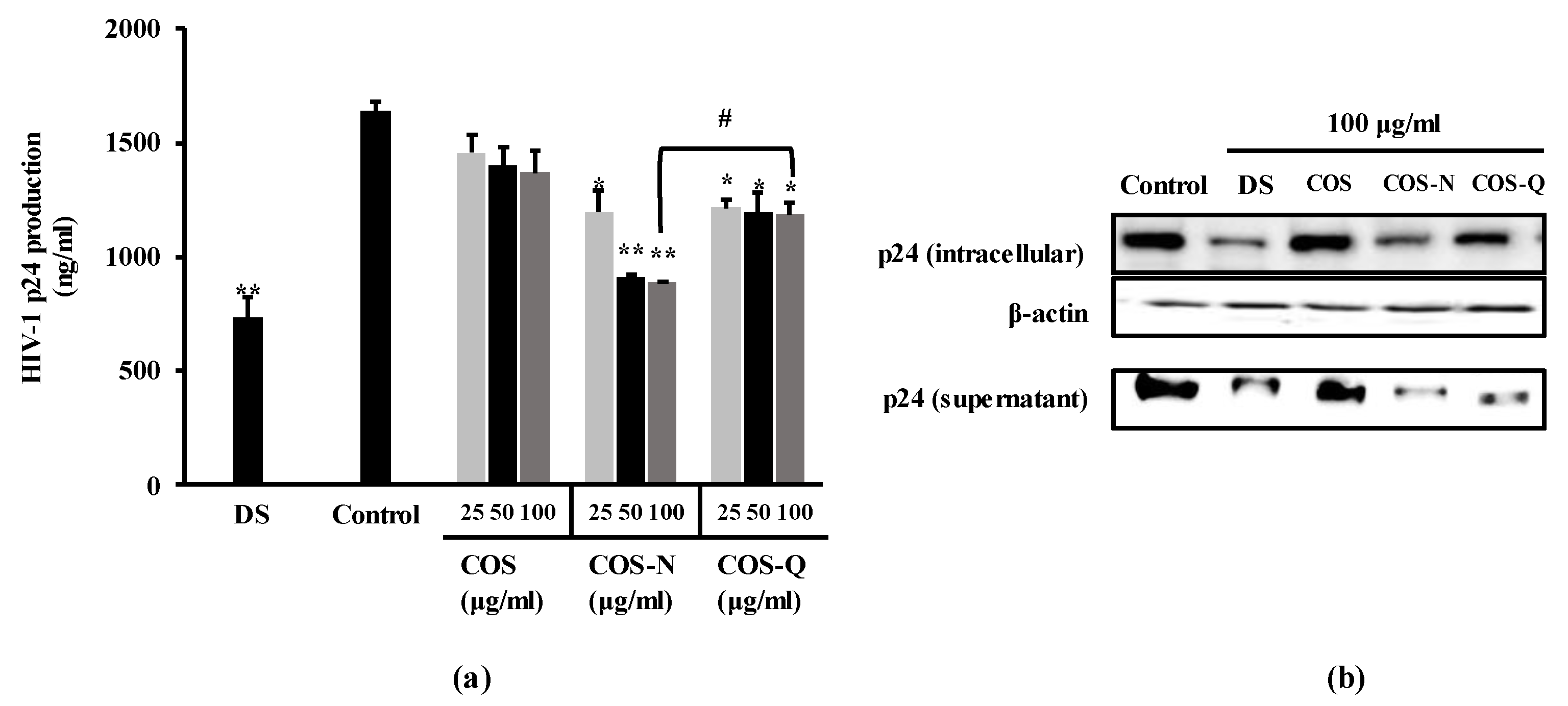
2.1.2. Inhibition of the HIV-1 p24 Antigen
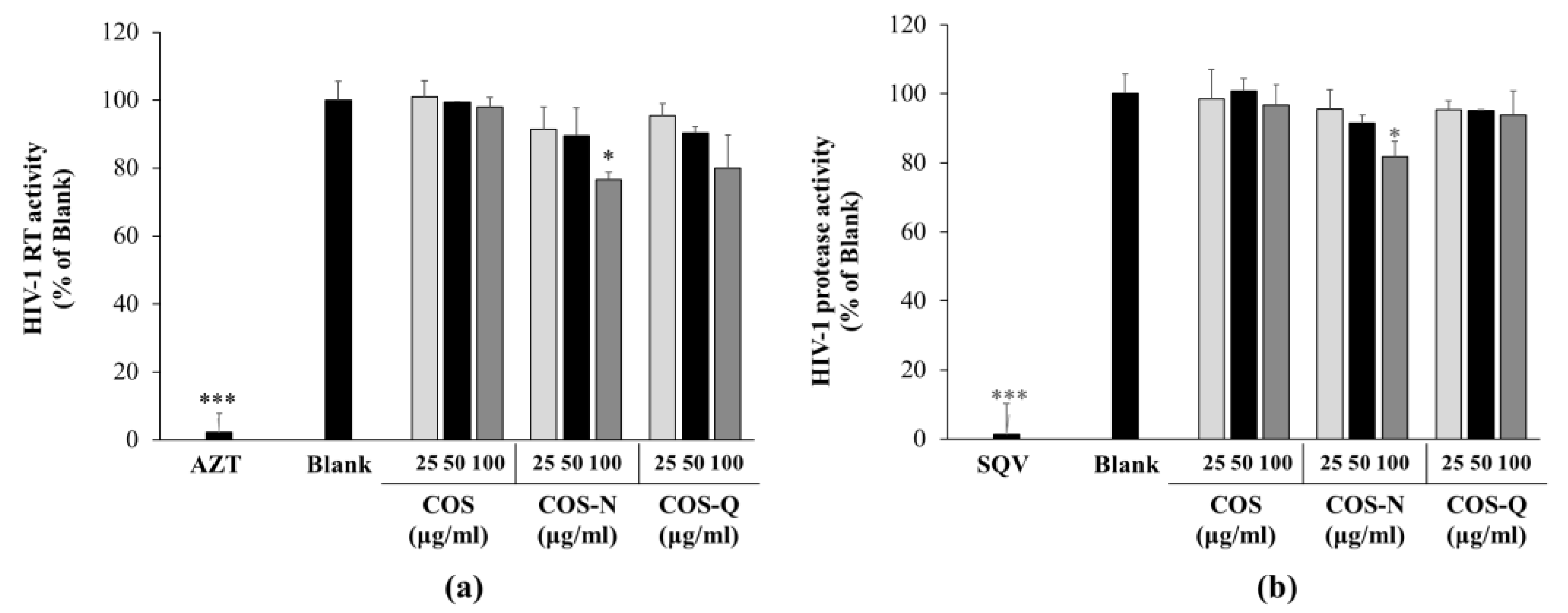
2.2. Inhibition of HIV-1 Reverse Transcriptase (RT) and Protease Activity
2.3. Inhibition of HIV-1 Entry
3. Materials and Methods
3.1. Materials
3.2. Addition of Amino Acids to COS
3.3. Cell Culture, HIV-1 Infection, Syncytia Formation, and Cell Viability Analysis
3.4. Measurement of p24 Antigen
3.5. HIV-1 RT and Protease Activity Assay
3.6. Delayed Addition and Co-Culture Assays
3.7. Analysis of gp120-CD4 Binding
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ginsberg, H.S.; Goebel, W.F.; Horsfall, F.L. The inhibitory effect of polysaccharide on mumps virus multiplication. J. Exp. Med. 1948, 87, 385–410. [Google Scholar] [PubMed]
- Green, R.H.; Woolley, D.W. Inhibition by certain polysaccharides of hemagglutination and of multiplication of influenza virus. J. Exp. Med. 1947, 86, 55–64. [Google Scholar]
- He, X.; Fang, J.; Guo, Q.; Wang, M.; Li, Y.; Meng, Y.; Huang, L. Advances in antiviral polysaccharides derived from edible and medicinal plants and mushrooms. Carbohydr. Polym. 2020, 229, 115548. [Google Scholar] [PubMed]
- Battulga, T.; Tumurbaatar, O.; Ganzorig, O.; Ishimura, T.; Kanamoto, T.; Nakashima, H.; Miyazaki, K.; Yoshida, T. Analysis of interaction between sulfated polysaccharides and HIV oligopeptides by surface plasmon resonance. Int. J. Biol. Macromol. 2019, 125, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, H.; Tian, N.; Wang, H.; Yan, H. Sulfate Modification and Evaluation of in vitro Anti-HIV Activity of Lycium Barbarum Polysaccharides. ChemistrySelect 2022, 7, e202200695. [Google Scholar]
- Geetha Bai, R.; Tuvikene, R. Potential Antiviral Properties of Industrially Important Marine Algal Polysaccharides and Their Significance in Fighting a Future Viral Pandemic. Viruses 2021, 13, 1817. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Yang, Z.; Chen, J.; Wang, D.; Zhang, Y. Recent advances in antiviral activities and potential mechanisms of sulfated polysaccharides. Carbohydr. Polym. 2021, 272, 118526. [Google Scholar] [CrossRef]
- Lopes, J.L.; Quinteiro, V.S.T.; Wouk, J.; Darido, M.L.; Dekker, R.F.H.; Barbosa-Dekker, A.M.; Vetvicka, V.; Cunha, M.A.A.; Faccin-Galhardi, L.C.; Orsato, A. Sulfonated and Carboxymethylated β-Glucan Derivatives with Inhibitory Activity against Herpes and Dengue Viruses. Int. J. Mol. Sci. 2021, 22, 11013. [Google Scholar]
- Crini, G. Historical review on chitin and chitosan biopolymers. Environ. Chem. Lett. 2019, 17, 1623–1643. [Google Scholar] [CrossRef]
- El Knidri, H.; Belaabed, R.; Addaou, A.; Laajeb, A.; Lahsini, A. Extraction, chemical modification and characterization of chitin and chitosan. Int. J. Biol. Macromol. 2018, 120, 1181–1189. [Google Scholar]
- Naveed, M.; Phil, L.; Sohail, M.; Hasnat, M.; Baig, M.M.F.A.; Ihsan, A.U.; Shumzaid, M.; Kakar, M.U.; Khan, T.M.; Akabar, M.D.; et al. Chitosan oligosaccharide (COS): An overview. Int. J. Biol. Macromol. 2019, 129, 827–843. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Li, K.; Li, P. Review: Advances in preparation of chitooligosaccharides with heterogeneous sequences and their bioactivity. Carbohydr. Polym. 2021, 252, 117206. [Google Scholar] [CrossRef]
- Singh, A.; Mittal, A.; Benjakul, S. Chitosan, Chitooligosaccharides and Their Polyphenol Conjugates: Preparation, Bioactivities, Functionalities and Applications in Food Systems. Food Rev. Int. 2021, 1–23. [Google Scholar] [CrossRef]
- Park, H.-H.; Ko, S.-C.; Oh, G.-W.; Jang, Y.-M.; Kim, Y.-M.; Park, W.S.; Choi, I.-W.; Jung, W.-K. Characterization and biological activity of PVA hydrogel containing chitooligosaccharides conjugated with gallic acid. Carbohydr. Polym. 2018, 198, 197–205. [Google Scholar] [PubMed]
- Kumar, M.; Rajput, M.; Soni, T.; Vivekanand, V.; Pareek, N. Chemoenzymatic Production and Engineering of Chitooligosaccharides and N-acetyl Glucosamine for Refining Biological Activities. Front. Chem. 2020, 8, 469. [Google Scholar] [PubMed]
- Tabassum, N.; Ahmed, S.; Azam Ali, M. Chitooligosaccharides for Drug Delivery. In Chitooligosaccharides; Springer International Publishing: Cham, Switzerland, 2022; pp. 309–332. [Google Scholar]
- El-Sayed, N.S.; Shirazi, A.N.; El-Meligy, M.G.; El-Ziaty, A.K.; Nagieb, Z.A.; Parang, K.; Tiwari, R.K. Design, synthesis, and evaluation of chitosan conjugated GGRGDSK peptides as a cancer cell-targeting molecular transporter. Int. J. Biol. Macromol. 2016, 87, 611–622. [Google Scholar]
- Dang, Z.; Zhu, L.; Lai, W.; Bogerd, H.; Lee, K.-H.; Huang, L.; Chen, C.-H. Aloperine and Its Derivatives as a New Class of HIV-1 Entry Inhibitors. ACS Med. Chem. Lett. 2016, 7, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Fan, L.; Liu, Y.; Li, J. Recent advances on formation mechanism and functionality of chitosan-based conjugates and their application in o/w emulsion systems: A review. Food Chem. 2022, 380, 131838. [Google Scholar] [CrossRef]
- Pattnaik, A.; Pati, S.; Samal, S.K. Chitosan-Polyphenol Conjugates for Human Health. Life 2022, 12, 1768. [Google Scholar] [CrossRef]
- Ai, H.; Wang, F.; Xia, Y.; Chen, X.; Lei, C. Antioxidant, antifungal and antiviral activities of chitosan from the larvae of housefly, Musca domestica L. Food Chem. 2012, 132, 493–498. [Google Scholar]
- Davydova, V.N.; Nagorskaya, V.P.; Gorbach, V.I.; Kalitnik, A.A.; Reunov, A.V.; Solov’eva, T.F.; Ermak, I.M. Chitosan antiviral activity: Dependence on structure and depolymerization method. Appl. Biochem. Microbiol. 2011, 47, 103–108. [Google Scholar]
- Su, X.; Zivanovic, S.; D’souza, D.H. Effect of Chitosan on the Infectivity of Murine Norovirus, Feline Calicivirus, and Bacteriophage MS2. J. Food Prot. 2009, 72, 2623–2628. [Google Scholar] [PubMed]
- Zheng, M.; Qu, D.; Wang, H.; Sun, Z.; Liu, X.; Chen, J.; Li, C.; Li, X.; Chen, Z. Intranasal Administration of Chitosan Against Influenza A (H7N9) Virus Infection in a Mouse Model. Sci. Rep. 2016, 6, 28729. [Google Scholar] [CrossRef] [PubMed]
- Ciejka, J.; Wolski, K.; Nowakowska, M.; Pyrc, K.; Szczubiałka, K. Biopolymeric nano/microspheres for selective and reversible adsorption of coronaviruses. Mater. Sci. Eng. C 2017, 76, 735–742. [Google Scholar] [CrossRef]
- Nishimura, S.-I.; Kai, H.; Shinada, K.; Yoshida, T.; Tokura, S.; Kurita, K.; Nakashima, H.; Yamamoto, N.; Uryu, T. Regioselective syntheses of sulfated polysaccharides: Specific anti-HIV-1 activity of novel chitin sulfates. Carbohydr. Res. 1998, 306, 427–433. [Google Scholar] [CrossRef]
- Artan, M.; Karadeniz, F.; Karagozlu, M.Z.; Kim, M.-M.; Kim, S.-K. Anti-HIV-1 activity of low molecular weight sulfated chitooligosaccharides. Carbohydr. Res. 2010, 345, 656–662. [Google Scholar] [CrossRef]
- Lv, H.-X.; Zhang, Z.-H.; Wang, X.-P.; Cheng, Q.-Q.; Wang, W.; Huang, X.-H.; Zhou, J.-P.; Zhang, Q.; Hou, L.-L.; Huo, W. A Biomimetic Chitosan Derivates: Preparation, Characterization and Transdermal Enhancement Studies of N-Arginine Chitosan. Molecules 2011, 16, 6778–6790. [Google Scholar]
- Jaber, N.; Al-Remawi, M.; Al-Akayleh, F.; Al-Muhtaseb, N.; Al-Adham, I.S.I.; Collier, P.J. A review of the antiviral activity of Chitosan, including patented applications and its potential use against COVID-19. J. Appl. Microbiol. 2022, 132, 41–58. [Google Scholar] [CrossRef]
- Perfettini, J.-L.; Castedo, M.; Roumier, T.; Andreau, K.; Nardacci, R.; Piacentini, M.; Kroemer, G. Mechanisms of apoptosis induction by the HIV-1 envelope. Cell Death Differ. 2005, 12, 916–923. [Google Scholar] [CrossRef]
- Khandazhinskaya, A.; Matyugina, E.; Shirokova, E. Anti-HIV therapy with AZT prodrugs: AZT phosphonate derivatives, current state and prospects. Expert. Opin. Drug Metab. Toxicol. 2010, 6, 701–714. [Google Scholar]
- Pereira, M.; Vale, N. Saquinavir: From HIV to COVID-19 and Cancer Treatment. Biomolecules 2022, 12, 944. [Google Scholar] [PubMed]
- Hussain, W.; Majeed, A.; Akhtar, A.; Rasool, N. Computational Studies of 3D-QSAR on a Highly Active Series of Naturally Occurring Nonnucleoside Inhibitors of HIV-1 RT (NNRTI). J. Comput. Biophys. Chem. 2021, 20, 3–11. [Google Scholar]
- Wong, J.H.; Ng, T.B.; Wang, H.; Cheung, R.C.F.; Ng, C.C.W.; Ye, X.; Yang, J.; Liu, F.; Ling, C.; Chan, K.; et al. Antifungal Proteins with Antiproliferative Activity on Cancer Cells and HIV-1 Enzyme Inhibitory Activity from Medicinal Plants and Medicinal Fungi. Curr. Protein Pept. Sci. 2019, 20, 265–276. [Google Scholar] [PubMed]
- Emaminia, F.; Skarnovich, M.; Yu, P.A.; Cheresiz, S.V. An Amide Derivative of Betulonic Acid as a New Inhibitor of Sars-CoV-2 Spike Protein-Mediated Cell Entry and Sars-CoV-2 Infection. Insights Chem. Biochem. 2022, 2, 1–11. [Google Scholar]
- Moonsamy, S.; Dash, R.; Soliman, M. Integrated Computational Tools for Identification of CCR5 Antagonists as Potential HIV-1 Entry Inhibitors: Homology Modeling, Virtual Screening, Molecular Dynamics Simulations and 3D QSAR Analysis. Molecules 2014, 19, 5243–5265. [Google Scholar]
- Dang, Z.; Qian, K.; Ho, P.; Zhu, L.; Lee, K.-H.; Huang, L.; Chen, C.-H. Synthesis of betulinic acid derivatives as entry inhibitors against HIV-1 and bevirimat-resistant HIV-1 variants. Bioorg. Med. Chem. Lett. 2012, 22, 5190–5194. [Google Scholar] [CrossRef]
- Yoshida, T. Anti-HIV Mechanism of Sulfated Poly and Oligosaccharides. J. Fiber Sci. Technol. 2020, 76, 387–402. [Google Scholar]
- Jeon, Y.-J.; Kim, S.-K. Effect of Antimicrobial Activity by Chitosan Oligosaccharide N-Conjugated with Asparagine. J. Microbiol. Biotechnol. 2001, 11, 281–286. [Google Scholar]
- Crublet, E.; Andrieu, J.-P.; Vives, R.R.; Lortat-Jacob, H. The HIV-1 envelope glycoprotein gp120 features four heparan sulfate binding domains, including the co-receptor binding site. J. Biol. Chem. 2008, 283, 193–200. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karadeniz, F. In Vitro Anti-HIV-1 Activity of Chitosan Oligomers N-Conjugated with Asparagine and Glutamine. BioTech 2023, 12, 18. https://doi.org/10.3390/biotech12010018
Karadeniz F. In Vitro Anti-HIV-1 Activity of Chitosan Oligomers N-Conjugated with Asparagine and Glutamine. BioTech. 2023; 12(1):18. https://doi.org/10.3390/biotech12010018
Chicago/Turabian StyleKaradeniz, Fatih. 2023. "In Vitro Anti-HIV-1 Activity of Chitosan Oligomers N-Conjugated with Asparagine and Glutamine" BioTech 12, no. 1: 18. https://doi.org/10.3390/biotech12010018
APA StyleKaradeniz, F. (2023). In Vitro Anti-HIV-1 Activity of Chitosan Oligomers N-Conjugated with Asparagine and Glutamine. BioTech, 12(1), 18. https://doi.org/10.3390/biotech12010018