
Recent Developments in Protein Lactylation in PTSD and CVD: Novel Strategies and Targets


Abstract
:1. Introduction
2. Human Stress Response: A Quick Reminder
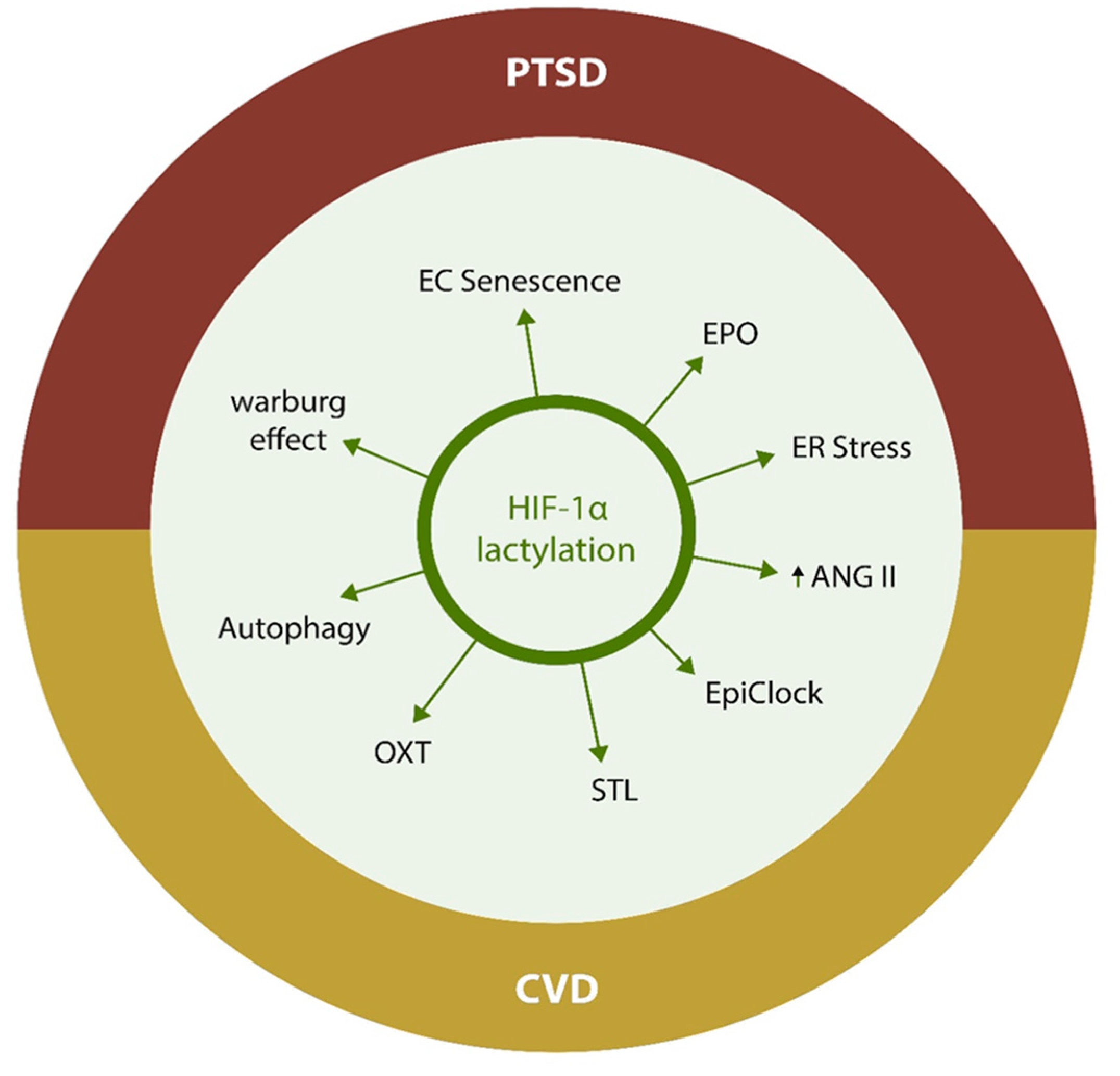
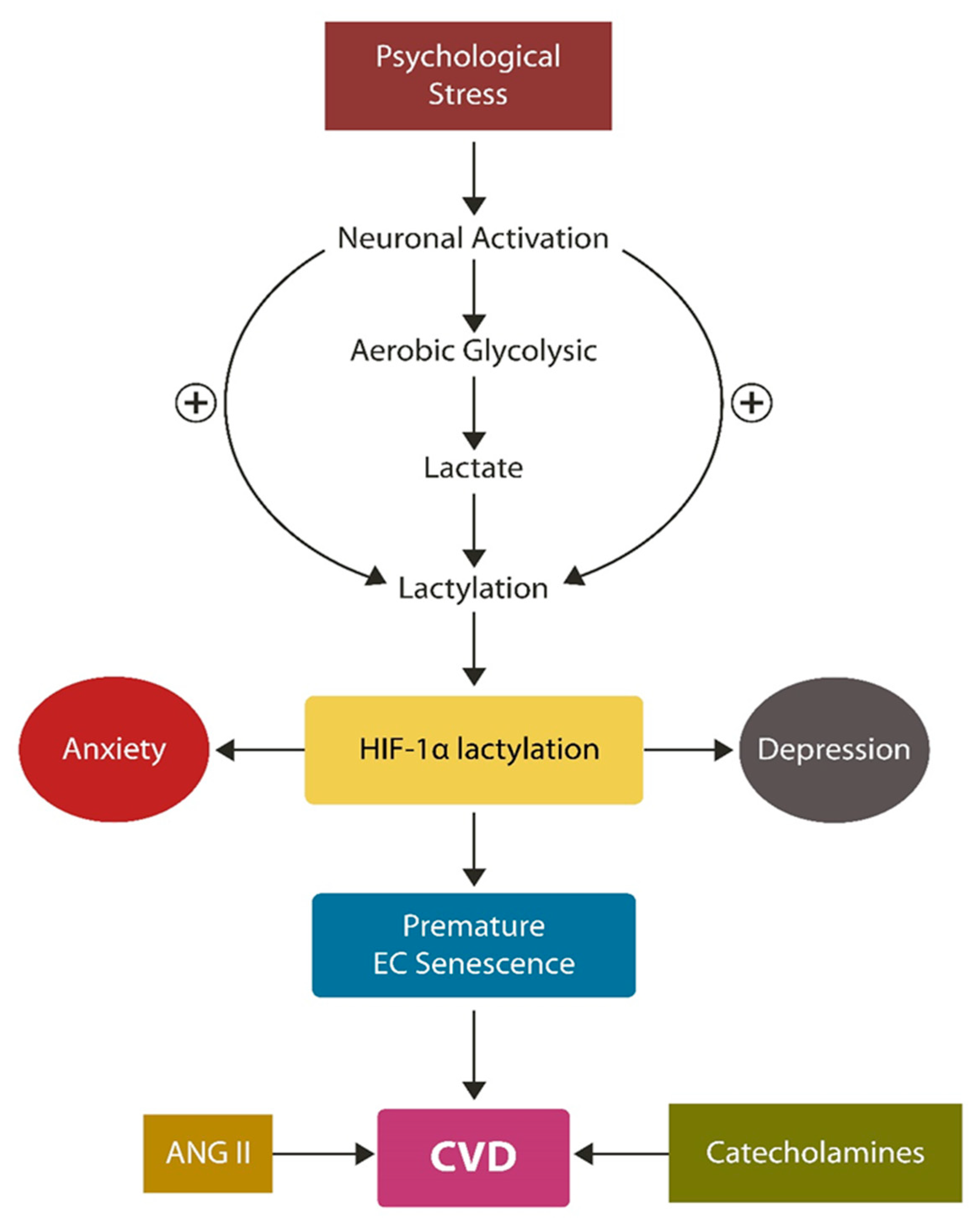
3. Protein Lactylation, the Warburg Effect 2.0
3.1. Virus-Induced PTSD and CVD
3.2. Virus-Induced Senescence (VIS)
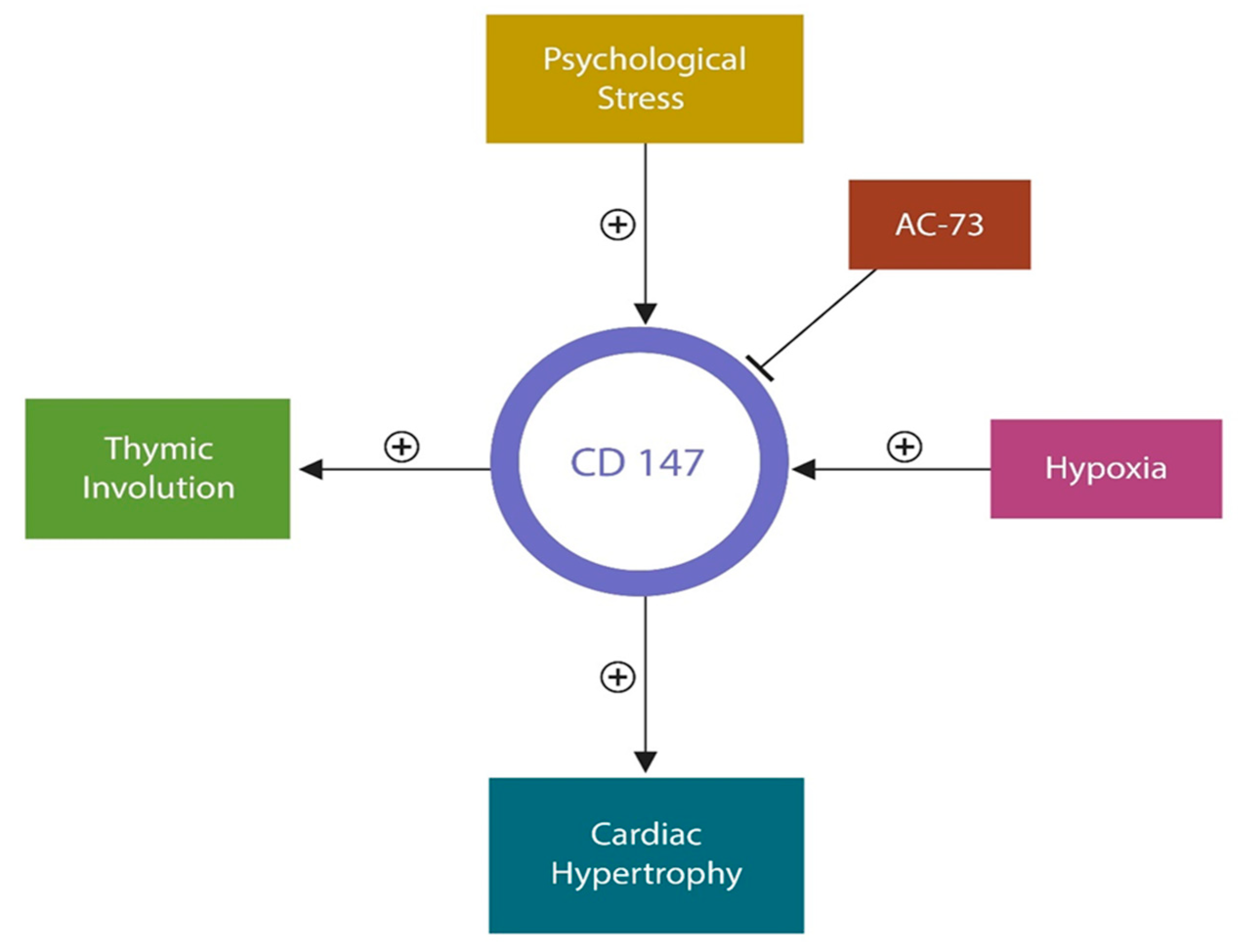
4. From Cellular to Organismal Stress
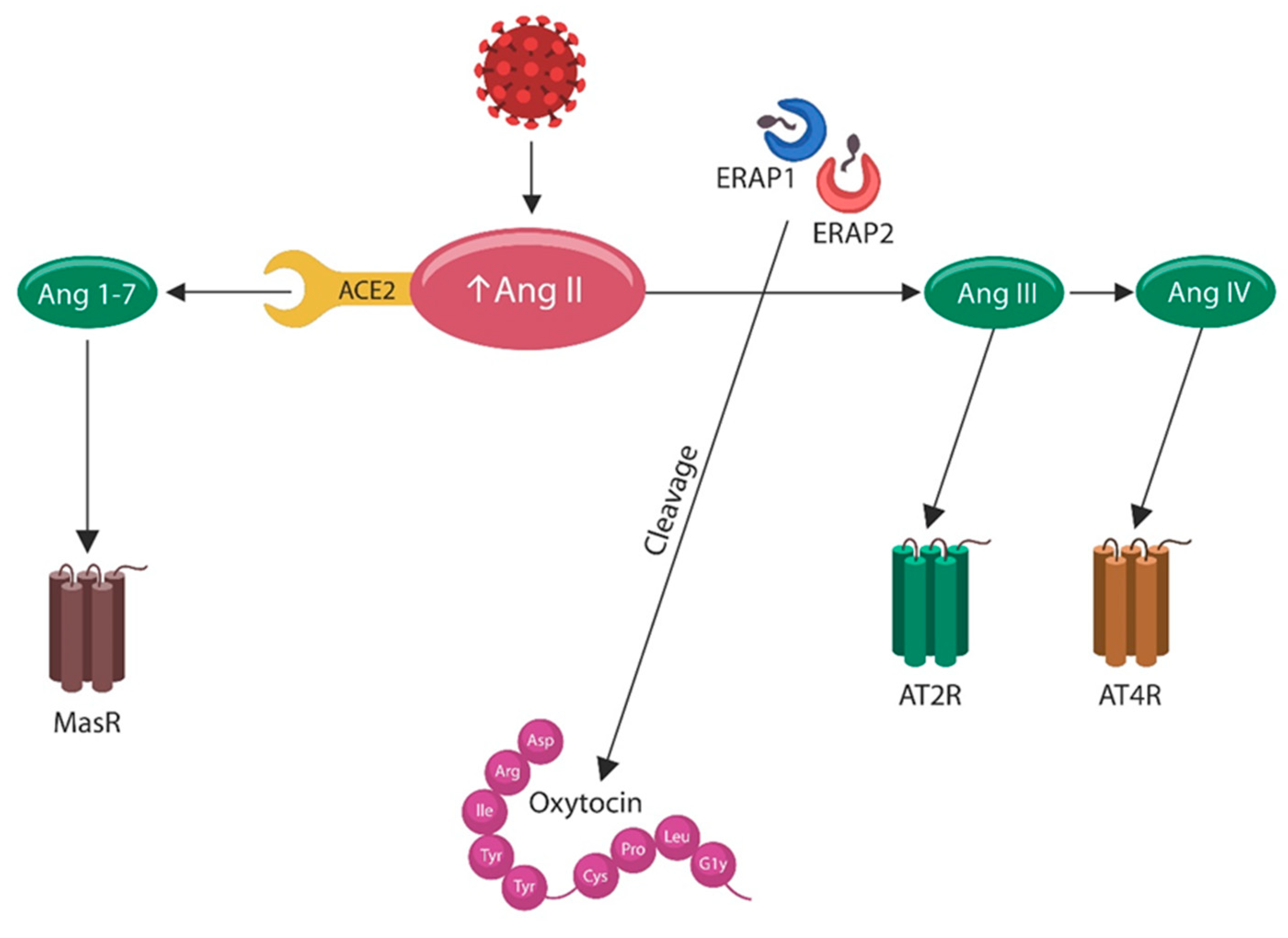
5. The Mystery of ERAP2
- A recombinant ERAP2 may be beneficial for the treatment of PTSD and CVD;
- ERAP2 may be a viable marker for PTSD vulnerability.
6. Potential Interventions
6.1. OXT
6.2. HIF-1α
6.3. ASIC1 Blockers
- -
- -
- C5B, a synthetic ASIC1 inhibitor, crosses the BBB and protects against neuronal apoptosis, indicating potential benefits for PTSD and CVD [216].
- -
- -
6.4. Lactylation
- -
- -
- Oxamate has been found helpful for both CVD and psychological stress, indicating potential PTSD benefits [232].
- -
- Quinoline 3-sulfonamides, is a novel LDH inhibitor that reverses aerobic glycolysis in cancer cells, and various quinoline derivatives have been tested as anticancer agents [233,234]. Quinolines have been known for their antidepressant and antipsychotic properties, as aripiprazole is derived from these compounds [235]. Several studies have demonstrated the beneficial effect of aripiprazole on PTSD, suggesting that these drugs should be further interrogated [236,237].
- -
- Galloflavin is a newer LDH inhibitor that, to our knowledge, has not been assessed for CVD or PTSD, but its pharmacological profile suggests potential efficacy.
6.5. Interleukin 7
6.6. Autophagy and UPR
6.7. Significance for the Field and Novel Targets
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tasdemir, N.; Lowe, S.W. Senescent cells spread the word: Non-cell autonomous propagation of cellular senescence. EMBO J. 2013, 32, 1975–1976. [Google Scholar] [CrossRef]
- Faget, D.V.; Ren, Q.; Stewart, S.A. Unmasking senescence: Context-dependent effects of SASP in cancer. Nat. Rev. Cancer 2019, 19, 439–453. [Google Scholar] [CrossRef]
- Wolf, E.J.; Schnurr, P.P. PTSD-Related Cardiovascular Disease and Accelerated Cellular Aging. Psychiatry Ann. 2016, 46, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Kuan, P.F.; Waszczuk, M.A.; Kotov, R.; Marsit, C.J.; Guffanti, G.; Gonzalez, A.; Yang, X.; Koenen, K.; Bromet, E.; Luft, B.J. An epigenome-wide DNA methylation study of PTSD and depression in World Trade Center responders. Transl. Psychiatry 2017, 7, e1158. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhang, Y. Reversing DNA methylation: Mechanisms, genomics, and biological functions. Cell 2014, 156, 45–68. [Google Scholar] [CrossRef]
- Katrinli, S.; Stevens, J.; Wani, A.H.; Lori, A.; Kilaru, V.; van Rooij, S.J.H.; Hinrichs, R.; Powers, A.; Gillespie, C.F.; Michopoulos, V.; et al. Evaluating the impact of trauma and PTSD on epigenetic prediction of lifespan and neural integrity. Neuropsychopharmacology 2020, 45, 1609–1616. [Google Scholar] [CrossRef]
- Li, D.; Wang, L.; Zhou, Z.; Song, L.; Chen, S.; Yang, Y.; Hu, Y.; Wang, Y.; Wu, S.; Tian, Y. Lifetime risk of cardiovascular disease and life expectancy with and without cardiovascular disease according to changes in metabolic syndrome status. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Hamadneh, L.; Al-Lakkis, L.; Alhusban, A.A.; Tarawneh, S.; Abu-Irmaileh, B.; Albustanji, S.; Al-Bawab, A.Q. Changes in Lactate Production, Lactate Dehydrogenase Genes Expression and DNA Methylation in Response to Tamoxifen Resistance Development in MCF-7 Cell Line. Genes 2021, 12, 777. [Google Scholar] [CrossRef]
- Cimmino, F.; Avitabile, M.; Lasorsa, V.A.; Montella, A.; Pezone, L.; Cantalupo, S.; Capasso, M. HIF-1 transcription activity: HIF1A driven response in normoxia and in hypoxia. BMC Med. Genet. 2019, 20, 37. [Google Scholar] [CrossRef]
- Welford, S.M.; Bedogni, B.; Gradin, K.; Poellinger, L.; Broome Powell, M.; Giaccia, A.J. HIF1alpha delays premature senescence through the activation of MIF. Genes Dev. 2006, 20, 3366–3371. [Google Scholar] [CrossRef]
- Hopkins, R.O.; Weaver, L.K. Acute psychosis associated with diving. Undersea Hyperb. Med. 2001, 28, 145–148. [Google Scholar] [PubMed]
- Chiner, E.; Arriero, J.M.; Signes-Costa, J.; Marco, J. Acute psychosis after CPAP treatment in a schizophrenic patient with sleep apnoea-hypopnoea syndrome. Eur. Respir. J. 2001, 17, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Burtscher, J.; Niedermeier, M.; Hüfner, K.; van den Burg, E.; Kopp, M.; Stoop, R.; Burtscher, M.; Gatterer, H.; Millet, G.P. The interplay of hypoxic and mental stress: Implications for anxiety and depressive disorders. Neurosci. Biobehav. Rev. 2022, 138, 104718. [Google Scholar] [CrossRef]
- Vijayarangan, A.; Deru, K.; Hebert, D. Hyperbaric oxygen for mTBI-associated PCS and PTSD: Pooled analysis of results from Department of Defense and other published studies. Undersea Hyperb. Med. 2019, 46, 353–383. [Google Scholar]
- Parr, N.J.; Anderson, J.; Veazie, S. Evidence. Brief: Hyperbaric Oxygen Therapy for Traumatic Brain Injury and/or Post-Traumatic Stress Disorder; Department of Veterans Affairs (US): Washington, DC, USA, 2021. [Google Scholar]
- Bloch, Y.; Applebaum, J.; Osher, Y.; Amar, S.; Azab, A.N.; Agam, G.; Belmaker, R.H.; Bersudsky, Y. Normobaric hyperoxia treatment of schizophrenia. J. Clin. Psychopharmacol. 2012, 32, 525–530. [Google Scholar] [CrossRef]
- Akman, M.; Belisario, D.C.; Salaroglio, I.C.; Kopecka, J.; Donadelli, M.; De Smaele, E.; Riganti, C. Hypoxia, endoplasmic reticulum stress and chemoresistance: Dangerous liaisons. J. Exp. Clin. Cancer Res. 2021, 40, 28. [Google Scholar] [CrossRef]
- Nevell, L.; Zhang, K.; Aiello, A.E.; Koenen, K.; Galea, S.; Soliven, R.; Zhang, C.; Wildman, D.E.; Uddin, M. Elevated systemic expression of ER stress related genes is associated with stress-related mental disorders in the Detroit Neighborhood Health Study. Psychoneuroendocrinology 2014, 43, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Kim, K.; Kim, J.H.; Park, Y. The Role of Endoplasmic Reticulum Stress in Cardiovascular Disease and Exercise. Int. J. Vasc. Med. 2017, 2017, 2049217. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Hu, Y.; Ruan, L.; Ji, Y.; Lou, Z. Role of endoplasmic reticulum stress in depression (Review). Mol. Med. Rep. 2019, 20, 4774–4780. [Google Scholar] [CrossRef]
- Lin, J.H.; Walter, P.; Yen, T.S. Endoplasmic reticulum stress in disease pathogenesis. Annu. Rev. Pathol. 2008, 3, 399–425. [Google Scholar] [CrossRef]
- Chipurupalli, S.; Kannan, E.; Tergaonkar, V.; D’Andrea, R.; Robinson, N. Hypoxia Induced ER Stress Response as an Adaptive Mechanism in Cancer. Int. J. Mol. Sci. 2019, 20, 749. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Kitakaze, M. ER stress in cardiovascular disease. J. Mol. Cell Cardiol. 2010, 48, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Seligowski, A.V.; Misganaw, B.; Duffy, L.A.; Ressler, K.J.; Guffanti, G. Leveraging Large-Scale Genetics of PTSD and Cardiovascular Disease to Demonstrate Robust Shared Risk and Improve Risk Prediction Accuracy. Am. J. Psychiatry 2022, 179, 814–823. [Google Scholar] [CrossRef]
- Pullamsetti, S.S.; Mamazhakypov, A.; Weissmann, N.; Seeger, W.; Savai, R. Hypoxia-inducible factor signaling in pulmonary hypertension. J. Clin. Invest. 2020, 130, 5638–5651. [Google Scholar] [CrossRef]
- Culley, M.K.; Chan, S.Y. Endothelial Senescence: A New Age in Pulmonary Hypertension. Circ. Res. 2022, 130, 928–941. [Google Scholar] [CrossRef]
- Alique, M.; Sánchez-López, E.; Bodega, G.; Giannarelli, C.; Carracedo, J.; Ramírez, R. Hypoxia-Inducible Factor-1α: The Master Regulator of Endothelial Cell Senescence in Vascular Aging. Cells 2020, 9, 195. [Google Scholar] [CrossRef]
- Bussotti, M.; Sommaruga, M. Anxiety and depression in patients with pulmonary hypertension: Impact and management challenges. Vasc. Health. Risk. Manag. 2018, 14, 349–360. [Google Scholar] [CrossRef] [PubMed]
- McDivitt, J.D.; Barstow, C. Cardiovascular Disease Update: Pulmonary Hypertension. FP Essent. 2017, 454, 24–28. [Google Scholar] [PubMed]
- Sabbatinelli, J.; Prattichizzo, F.; Olivieri, F.; Procopio, A.D.; Rippo, M.R.; Giuliani, A. Where Metabolism Meets Senescence: Focus on Endothelial Cells. Front. Physiol. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Eelen, G.; de Zeeuw, P.; Treps, L.; Harjes, U.; Wong, B.W.; Carmeliet, P. Endothelial Cell Metabolism. Physiol. Rev. 2018, 98, 3–58. [Google Scholar] [CrossRef]
- Fitzgerald, G.; Soro-Arnaiz, I.; De Bock, K. The Warburg Effect in Endothelial Cells and its Potential as an Anti-angiogenic Target in Cancer. Front. Cell Dev. Biol. 2018, 6, 100. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, J.M.; Martino, F.; Thum, T. Epigenetic modifications in cardiovascular disease. Basic Res. Cardiol. 2012, 107, 245. [Google Scholar] [CrossRef] [PubMed]
- Hermann, R.; Lay, D.; Wahl, P.; Roth, W.T.; Petrowski, K. Effects of psychosocial and physical stress on lactate and anxiety levels. Stress 2019, 22, 664–669. [Google Scholar] [CrossRef]
- Hagihara, H.; Shoji, H.; Otabi, H.; Toyoda, A.; Katoh, K.; Namihira, M.; Miyakawa, T. Protein lactylation induced by neural excitation. Cell Rep. 2021, 37, 109820. [Google Scholar] [CrossRef] [PubMed]
- Sara, J.D.S.; Toya, T.; Ahmad, A.; Clark, M.M.; Gilliam, W.P.; Lerman, L.O.; Lerman, A. Mental Stress and Its Effects on Vascular Health. Mayo. Clin. Proc. 2022, 97, 951–990. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. Endothelial Barrier and Its Abnormalities in Cardiovascular Disease. Front. Physiol. 2015, 6, 365. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Yang, Z.; Yu, Y.; Zhang, P. HIF1α lactylation enhances KIAA1199 transcription to promote angiogenesis and vasculogenic mimicry in prostate cancer. Int. J. Biol. Macromol. 2022, 222, 2225–2243. [Google Scholar] [CrossRef]
- Diekmann, F.; Legchenko, E.; Chouvarine, P.; Lichtinghagen, R.; Bertram, H.; Happel, C.M.; Hansmann, G. Circulating Interleukin-7 in Human Pulmonary Arterial Hypertension. Front. Cardiovasc. Med. 2021, 8, 794549. [Google Scholar] [CrossRef]
- Mori, K.; Nakaya, Y.; Sakamoto, S.; Hayabuchi, Y.; Matsuoka, S.; Kuroda, Y. Lactate-induced vascular relaxation in porcine coronary arteries is mediated by Ca2+-activated K+ channels. J. Mol. Cell Cardiol. 1998, 30, 349–356. [Google Scholar] [CrossRef]
- Duś, D.; Krawczenko, A.; Załecki, P.; Paprocka, M.; Wiedłocha, A.; Goupille, C.; Kieda, C. IL-7 receptor is present on human microvascular endothelial cells. Immunol. Lett. 2003, 86, 163–168. [Google Scholar] [CrossRef]
- Jacobs, S.R.; Michalek, R.D.; Rathmell, J.C. IL-7 is essential for homeostatic control of T cell metabolism in vivo. J. Immunol. 2010, 184, 3461–3469. [Google Scholar] [CrossRef]
- Shive, C.L.; Clagett, B.; McCausland, M.R.; Mudd, J.C.; Funderburg, N.T.; Freeman, M.L.; Younes, S.A.; Ferrari, B.M.; Rodriguez, B.; McComsey, G.A.; et al. Inflammation Perturbs the IL-7 Axis, Promoting Senescence and Exhaustion that Broadly Characterize Immune Failure in Treated HIV Infection. J. Acquir. Immune Defic. Syndr. 2016, 71, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.; Mendelsohn, A.; Larrick, J.W. Interleukin-7 and Immunosenescence. J. Immunol. Res. 2017, 2017, 4807853. [Google Scholar] [CrossRef]
- Ahlgrim, C.; Baumstark, M.W.; Roecker, K. Clarifying the Link Between the Blood Lactate Concentration and Cardiovascular Risk. Int. J. Sports Med. 2022, 43, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Unterluggauer, H.; Mazurek, S.; Lener, B.; Hütter, E.; Eigenbrodt, E.; Zwerschke, W.; Jansen-Dürr, P. Premature senescence of human endothelial cells induced by inhibition of glutaminase. Biogerontology 2008, 9, 247–259. [Google Scholar] [CrossRef]
- Nilsson, P.M. Early Vascular Aging in Hypertension. Front. Cardiovasc. Med. 2020, 7, 6. [Google Scholar] [CrossRef]
- Ramírez, R.; Ceprian, N.; Figuer, A.; Valera, G.; Bodega, G.; Alique, M.; Carracedo, J. Endothelial Senescence and the Chronic Vascular Diseases: Challenges and Therapeutic Opportunities in Atherosclerosis. J. Pers. Med. 2022, 12, 215. [Google Scholar] [CrossRef] [PubMed]
- Sher, L.D.; Geddie, H.; Olivier, L.; Cairns, M.; Truter, N.; Beselaar, L.; Essop, M.F. Chronic stress and endothelial dysfunction: Mechanisms, experimental challenges, and the way ahead. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H488–H506. [Google Scholar] [CrossRef] [PubMed]
- Tekin, D.; Dursun, A.D.; Xi, L. Hypoxia inducible factor 1 (HIF-1) and cardioprotection. Acta Pharmacol. Sin. 2010, 31, 1085–1094. [Google Scholar] [CrossRef]
- Mellon, S.H.; Bersani, F.S.; Lindqvist, D.; Hammamieh, R.; Donohue, D.; Dean, K.; Jett, M.; Yehuda, R.; Flory, J.; Reus, V.I.; et al. Metabolomic analysis of male combat veterans with post-traumatic stress disorder. PLoS ONE 2019, 14, e0213839. [Google Scholar] [CrossRef]
- Yang, J.; Liu, C.; Du, X.; Liu, M.; Ji, X.; Du, H.; Zhao, H. Hypoxia Inducible Factor 1α Plays a Key Role in Remote Ischemic Preconditioning Against Stroke by Modulating Inflammatory Responses in Rats. J. Am. Heart Assoc. 2018, 7, e007589. [Google Scholar] [CrossRef]
- Manukhina, E.B.; Tseilikman, V.E.; Karpenko, M.N.; Pestereva, N.S.; Tseilikman, O.B.; Komelkova, M.V.; Kondashevskaya, M.V.; Goryacheva, A.V.; Lapshin, M.S.; Platkovskii, P.O.; et al. Intermittent Hypoxic Conditioning Alleviates Post-Traumatic Stress Disorder-Induced Damage and Dysfunction of Rat Visceral Organs and Brain. Int. J. Mol. Sci. 2020, 21, 345. [Google Scholar] [CrossRef] [PubMed]
- Serebrovskaya, T.V.; Xi, L. Intermittent hypoxia training as non-pharmacologic therapy for cardiovascular diseases: Practical analysis on methods and equipment. Exp. Biol. Med. 2016, 241, 1708–1723. [Google Scholar] [CrossRef]
- Fahy, G.M.; Brooke, R.T.; Watson, J.P.; Good, Z.; Vasanawala, S.S.; Maecker, H.; Leipold, M.D.; Lin, D.T.S.; Kobor, M.S.; Horvath, S. Reversal of epigenetic aging and immunosenescent trends in humans. Aging Cell 2019, 18, e13028. [Google Scholar] [CrossRef] [PubMed]
- Rey, F.; Balsari, A.; Giallongo, T.; Ottolenghi, S.; Di Giulio, A.M.; Samaja, M.; Carelli, S. Erythropoietin as a Neuroprotective Molecule: An Overview of Its Therapeutic Potential in Neurodegenerative Diseases. ASN Neuro. 2019, 11, 1759091419871420. [Google Scholar] [CrossRef]
- Burger, D.; Xenocostas, A.; Feng, Q.P. Molecular basis of cardioprotection by erythropoietin. Curr. Mol. Pharmacol. 2009, 2, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Osborn, M.; Rustom, N.; Clarke, M.; Litteljohn, D.; Rudyk, C.; Anisman, H.; Hayley, S. Antidepressant-like effects of erythropoietin: A focus on behavioural and hippocampal processes. PLoS ONE 2013, 8, e72813. [Google Scholar] [CrossRef]
- Cantarelli, C.; Angeletti, A.; Cravedi, P. Erythropoietin, a multifaceted protein with innate and adaptive immune modulatory activity. Am. J. Transplant. 2019, 19, 2407–2414. [Google Scholar] [CrossRef]
- Sakanaka, M.; Wen, T.C.; Matsuda, S.; Masuda, S.; Morishita, E.; Nagao, M.; Sasaki, R. In vivo evidence that erythropoietin protects neurons from ischemic damage. Proc. Natl. Acad. Sci. USA 1998, 95, 4635–4640. [Google Scholar] [CrossRef]
- van der Meer, P.; Voors, A.A.; Lipsic, E.; van Gilst, W.H.; van Veldhuisen, D.J. Erythropoietin in cardiovascular diseases. Eur. Heart J. 2004, 25, 285–291. [Google Scholar] [CrossRef]
- Aiello, F.B.; Keller, J.R.; Klarmann, K.D.; Dranoff, G.; Mazzucchelli, R.; Durum, S.K. IL-7 induces myelopoiesis and erythropoiesis. J. Immunol. 2007, 178, 1553–1563. [Google Scholar] [CrossRef]
- Andrew, D.; Aspinall, R. Age-associated thymic atrophy is linked to a decline in IL-7 production. Exp. Gerontol. 2002, 37, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhang, D.; Wang, C.; Wu, Z.; Liang, C. The Pivotal Role of Thymus in Atherosclerosis Mediated by Immune and Inflammatory Response. Int. J. Med. Sci. 2018, 15, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, T.; Mizoi, Y.; Yamashita, A.; Yamada, M.; Yamamoto, Y.; Tatsuno, Y.; Nishi, K. Thymus of abused/neglected children. Forensic. Sci. Int. 1992, 53, 69–79. [Google Scholar] [CrossRef]
- Gruver, A.L.; Sempowski, G.D. Cytokines, leptin, and stress-induced thymic atrophy. J. Leukoc. Biol. 2008, 84, 915–923. [Google Scholar] [CrossRef]
- Odinokov, D.; Hamblin, M.R. Aging of lymphoid organs: Can photobiomodulation reverse age-associated thymic involution via stimulation of extrapineal melatonin synthesis and bone marrow stem cells? J. Biophotonics. 2018, 11, e201700282. [Google Scholar] [CrossRef] [PubMed]
- Geenen, V.; Legros, J.J.; Franchimont, P.; Baudrihaye, M.; Defresne, M.P.; Boniver, J. The neuroendocrine thymus: Coexistence of oxytocin and neurophysin in the human thymus. Science 1986, 232, 508–511. [Google Scholar] [CrossRef] [PubMed]
- Moll, U.M. Functional histology of the neuroendocrine thymus. Microsc. Res. Tech. 1997, 38, 300–310. [Google Scholar] [CrossRef]
- Tani-ichi, S.; Shimba, A.; Wagatsuma, K.; Miyachi, H.; Kitano, S.; Imai, K.; Hara, T.; Ikuta, K. Interleukin-7 receptor controls development and maturation of late stages of thymocyte subpopulations. Proc. Natl. Acad. Sci. USA 2013, 110, 612–617. [Google Scholar] [CrossRef] [PubMed]
- van Zuiden, M.; Frijling, J.L.; Nawijn, L.; Koch, S.B.J.; Goslings, J.C.; Luitse, J.S.; Biesheuvel, T.H.; Honig, A.; Veltman, D.J.; Olff, M. Intranasal Oxytocin to Prevent Posttraumatic Stress Disorder Symptoms: A Randomized Controlled Trial in Emergency Department Patients. Biol. Psychiatry 2017, 81, 1030–1040. [Google Scholar] [CrossRef]
- Petersson, M. Cardiovascular effects of oxytocin. Prog. Brain Res. 2002, 139, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Tirko, N.N.; Eyring, K.W.; Carcea, I.; Mitre, M.; Chao, M.V.; Froemke, R.C.; Tsien, R.W. Oxytocin Transforms Firing Mode of CA2 Hippocampal Neurons. Neuron 2018, 100, 593–608.e3. [Google Scholar] [CrossRef]
- Jankowski, M.; Broderick, T.L.; Gutkowska, J. The Role of Oxytocin in Cardiovascular Protection. Front. Psychol. 2020, 11, 2139. [Google Scholar] [CrossRef] [PubMed]
- Goldman, M.B. The mechanism of life-threatening water imbalance in schizophrenia and its relationship to the underlying psychiatric illness. Brain Res. Rev. 2009, 61, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Genel, O.; Pariante, C.M.; Borsini, A. The role of AQP4 in the pathogenesis of depression, and possible related mechanisms. Brain Behav. Immun. 2021, 98, 366–377. [Google Scholar] [CrossRef]
- Rutkovskiy, A.; Stensløkken, K.O.; Mariero, L.H.; Skrbic, B.; Amiry-Moghaddam, M.; Hillestad, V.; Valen, G.; Perreault, M.C.; Ottersen, O.P.; Gullestad, L.; et al. Aquaporin-4 in the heart: Expression, regulation and functional role in ischemia. Basic Res. Cardiol. 2012, 107, 280. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.C.; Wu, L.; Qian, M.Z.; Cai, P.P.; Liu, Q.B.; Zhao, G.X.; Li, Z.X.; Wu, Z.Y. Variants of Interleukin-7/Interleukin-7 Receptor Alpha are Associated with Both Neuromyelitis Optica and Multiple Sclerosis Among Chinese Han Population in Southeastern China. Chin. Med. J. 2015, 128, 3062–3068. [Google Scholar] [CrossRef]
- Wei, F.; Song, J.; Zhang, C.; Lin, J.; Xue, R.; Shan, L.D.; Gong, S.; Zhang, G.X.; Qin, Z.H.; Xu, G.Y.; et al. Chronic stress impairs the aquaporin-4-mediated glymphatic transport through glucocorticoid signaling. Psychopharmacology 2019, 236, 1367–1384. [Google Scholar] [CrossRef]
- Russell, G.; Lightman, S. The human stress response. Nat. Rev. Endocrinol. 2019, 15, 525–534. [Google Scholar] [CrossRef]
- Godoy, L.D.; Rossignoli, M.T.; Delfino-Pereira, P.; Garcia-Cairasco, N.; de Lima Umeoka, E.H. A Comprehensive Overview on Stress Neurobiology: Basic Concepts and Clinical Implications. Front. Behav. Neurosci. 2018, 12, 127. [Google Scholar] [CrossRef]
- Schneiderman, N.; Ironson, G.; Siegel, S.D. Stress and health: Psychological, behavioral, and biological determinants. Annu. Rev. Clin. Psychol. 2005, 1, 607–628. [Google Scholar] [CrossRef]
- Muscatell, K.A.; Merritt, C.C.; Cohen, J.R.; Chang, L.; Lindquist, K.A. The Stressed Brain: Neural Underpinnings of Social Stress Processing in Humans. Curr. Top. Behav. Neurosci. 2022, 54, 373–392. [Google Scholar] [CrossRef] [PubMed]
- Bremner, J.D. Traumatic stress: Effects on the brain. Dialogues Clin. Neurosci. 2006, 8, 445–461. [Google Scholar] [CrossRef]
- Janak, P.H.; Tye, K.M. From circuits to behaviour in the amygdala. Nature 2015, 517, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Tawakol, A.; Ishai, A.; Takx, R.A.; Figueroa, A.L.; Ali, A.; Kaiser, Y.; Truong, Q.A.; Solomon, C.J.; Calcagno, C.; Mani, V.; et al. Relation between resting amygdalar activity and cardiovascular events: A longitudinal and cohort study. Lancet 2017, 389, 834–845. [Google Scholar] [CrossRef] [PubMed]
- Hostinar, C.E.; Sullivan, R.M.; Gunnar, M.R. Psychobiological mechanisms underlying the social buffering of the hypothalamic–pituitary–adrenocortical axis: A review of animal models and human studies across development. Psychol. Bull. 2014, 140, 256–282. [Google Scholar] [CrossRef] [PubMed]
- Saha, S. Role of the central nucleus of the amygdala in the control of blood pressure: Descending pathways to medullary cardiovascular nuclei. Clin. Exp. Pharmacol. Physiol. 2005, 32, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Wellman, L.L.; Forcelli, P.A.; Aguilar, B.L.; Malkova, L. Bidirectional Control of Social Behavior by Activity within Basolateral and Central Amygdala of Primates. J. Neurosci. 2016, 36, 8746–8756. [Google Scholar] [CrossRef]
- Jackson, K.L.; Palma-Rigo, K.; Nguyen-Huu, T.P.; Davern, P.J.; Head, G.A. Major contribution of the medial amygdala to hypertension in BPH/2J genetically hypertensive mice. Hypertension 2014, 63, 811–818. [Google Scholar] [CrossRef]
- Marinzalda, M.L.; Pérez, P.A.; Gargiulo, P.A.; Casarsa, B.S.; Bregonzio, C.; Baiardi, G. Fear-potentiated behaviour is modulated by central amygdala angiotensin II AT1 receptors stimulation. Biomed. Res. Int. 2014, 2014, 183248. [Google Scholar] [CrossRef]
- Wiaderkiewicz, J.; Speth, R.C.; Park, J.; Marvar, P.J. Angiotensin II Type 2 Receptor-Expressing Neurons in the Central Amygdala Influence Fear-Related Behavior. Biol. Psychiatry 2019, 86, 899–909. [Google Scholar] [CrossRef]
- Labuschagne, I.; Phan, K.L.; Wood, A.; Angstadt, M.; Chua, P.; Heinrichs, M.; Stout, J.C.; Nathan, P.J. Oxytocin attenuates amygdala reactivity to fear in generalized social anxiety disorder. Neuropsychopharmacol 2010, 35, 2403–2413. [Google Scholar] [CrossRef] [PubMed]
- Ebner, K.; Bosch, O.J.; Krömer, S.A.; Singewald, N.; Neumann, I.D. Release of Oxytocin in the Rat Central Amygdala Modulates Stress-Coping Behavior and the Release of Excitatory Amino Acids. Neuropsychopharmacology 2005, 30, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Thibonnier, M.; Conarty, D.M.; Preston, J.A.; Plesnicher, C.L.; Dweik, R.A.; Erzurum, S.C. Human vascular endothelial cells express oxytocin receptors. Endocrinology 1999, 140, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Ishunina, T.A.; Swaab, D.F. Vasopressin and oxytocin neurons of the human supraoptic and paraventricular nucleus: Size changes in relation to age and sex. J. Clin. Endocrinol. Metab. 1999, 84, 4637–4644. [Google Scholar] [CrossRef]
- Wang, P.; Wang, S.C.; Liu, X.; Jia, S.; Wang, X.; Li, T.; Yu, J.; Parpura, V.; Wang, Y.F. Neural Functions of Hypothalamic Oxytocin and its Regulation. ASN Neuro. 2022, 14, 17590914221100706. [Google Scholar] [CrossRef]
- Sambuco, N.; Bradley, M.M.; Lang, P.J. Trauma-related dysfunction in the fronto-striatal reward circuit. J. Affect. Disord. 2021, 287, 359–366. [Google Scholar] [CrossRef]
- Taghva, A.; Oluigbo, C.; Corrigan, J.; Rezai, A.R. Posttraumatic stress disorder: Neurocircuitry and implications for potential deep brain stimulation. Stereotact. Funct. Neurosurg. 2013, 91, 207–219. [Google Scholar] [CrossRef]
- Logue, M.W.; van Rooij, S.J.H.; Dennis, E.L.; Davis, S.L.; Hayes, J.P.; Stevens, J.S.; Densmore, M.; Haswell, C.C.; Ipser, J.; Koch, S.B.J.; et al. Smaller Hippocampal Volume in Posttraumatic Stress Disorder: A Multisite ENIGMA-PGC Study: Subcortical Volumetry Results from Posttraumatic Stress Disorder Consortia. Biol. Psychiatry 2018, 83, 244–253. [Google Scholar] [CrossRef]
- Sailer, U.; Robinson, S.; Fischmeister, F.P.; König, D.; Oppenauer, C.; Lueger-Schuster, B.; Moser, E.; Kryspin-Exner, I.; Bauer, H. Altered reward processing in the nucleus accumbens and mesial prefrontal cortex of patients with posttraumatic stress disorder. Neuropsychologia 2008, 46, 2836–2844. [Google Scholar] [CrossRef]
- Hu, B.; Boyle, C.A.; Lei, S. Oxytocin receptors excite lateral nucleus of central amygdala by phospholipase Cβ- and protein kinase C-dependent depression of inwardly rectifying K+ channels. J. Physiol. 2020, 598, 3501–3520. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, J.A.; Szu, J.I.; Binder, D.K. The role of aquaporin-4 in synaptic plasticity, memory and disease. Brain Res. Bull. 2018, 136, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, A.O.; Lodder, E.M.; Wilders, R. Aquaporin Channels in the Heart-Physiology and Pathophysiology. Int. J. Mol. Sci. 2019, 20, 2039. [Google Scholar] [CrossRef]
- Li, Y.K.; Wang, F.; Wang, W.; Luo, Y.; Wu, P.F.; Xiao, J.L.; Hu, Z.L.; Jin, Y.; Hu, G.; Chen, J.G. Aquaporin-4 deficiency impairs synaptic plasticity and associative fear memory in the lateral amygdala: Involvement of downregulation of glutamate transporter-1 expression. Neuropsychopharmacology 2012, 37, 1867–1878. [Google Scholar] [CrossRef]
- Kalsi, A.S.; Greenwood, K.; Wilkin, G.; Butt, A.M. Kir4.1 expression by astrocytes and oligodendrocytes in CNS white matter: A developmental study in the rat optic nerve. J. Anat. 2004, 204, 475–485. [Google Scholar] [CrossRef]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218, Erratum in: Trends. Biochem. Sci. 2016, 41, 287. [Google Scholar] [CrossRef]
- Lee, T.Y. Lactate: A multifunctional signaling molecule. Yeungnam Univ. J. Med. 2021, 38, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; et al. Metabolic regulation of gene expression by histone lactylation. Nature 2019, 574, 575–580. [Google Scholar] [CrossRef]
- Yang, K.; Fan, M.; Wang, X.; Xu, J.; Wang, Y.; Tu, F.; Gill, P.S.; Ha, T.; Liu, L.; Williams, D.L.; et al. Lactate promotes macrophage HMGB1 lactylation, acetylation, and exosomal release in polymicrobial sepsis. Cell Death Differ. 2022, 29, 133–146. [Google Scholar] [CrossRef]
- Liebowitz, M.R.; Gorman, J.M.; Fyer, A.J.; Levitt, M.; Dillon, D.; Levy, G.; Klein, D.F. Lactate Provocation of Panic Attacks: II. Biochemical and Physiological Findings. Arch. Gen. Psychiatry 1985, 42, 709–719. [Google Scholar] [CrossRef]
- Jensen, C.F.; Keller, T.W.; Peskind, E.R.; McFall, M.E.; Veith, R.C.; Martin, D.; Wilkinson, C.W.; Raskind, M.A. Behavioral and neuroendocrine responses to sodium lactate infusion in subjects with posttraumatic stress disorder. Am. J. Psychiatry 1997, 154, 266–268. [Google Scholar] [CrossRef]
- Matsushita, K.; Williams, E.K.; Mongraw-Chaffin, M.L.; Coresh, J.; Schmidt, M.I.; Brancati, F.L.; Hoogeveen, R.C.; Ballantyne, C.M.; Young, J.H. The association of plasma lactate with incident cardiovascular outcomes: The ARIC Study. Am. J. Epidemiol. 2013, 178, 401–409. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, M.; Li, L.; Chen, L. Involvement of the Warburg effect in non-tumor diseases processes. J. Cell Physiol. 2018, 233, 2839–2849. [Google Scholar] [CrossRef]
- Azoulay, I.S.; Qi, X.; Rozenfeld, M.; Liu, F.; Hu, Q.; Ben Kasus Nissim, T.; Stavsky, A.; Zhu, M.X.; Xu, T.L.; Sekler, I. ASIC1a senses lactate uptake to regulate metabolism in neurons. Redox. Biol. 2022, 51, 102253. [Google Scholar] [CrossRef]
- Chen, L.; Huang, L.; Gu, Y.; Cang, W.; Sun, P.; Xiang, Y. Lactate-Lactylation Hands between Metabolic Reprogramming and Immunosuppression. Int. J. Mol. Sci. 2022, 23, 11943. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.M.; Lone, A.; Betts, D.H.; Cumming, R.C. Lactate preconditioning promotes a HIF-1α-mediated metabolic shift from OXPHOS to glycolysis in normal human diploid fibroblasts. Sci. Rep. 2020, 10, 8388. [Google Scholar] [CrossRef]
- Semenza, G.L. Hypoxia-inducible factor 1 and cardiovascular disease. Annu. Rev. Physiol. 2014, 76, 39–56. [Google Scholar] [CrossRef]
- Pan, R.Y.; He, L.; Zhang, J.; Liu, X.; Liao, Y.; Gao, J.; Liao, Y.; Yan, Y.; Li, Q.; Zhou, X.; et al. Positive feedback regulation of microglial glucose metabolism by histone H4 lysine 12 lactylation in Alzheimer’s disease. Cell Metab. 2022, 34, 634–648.e6. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Hu, H.; Liu, M.; Zhou, T.; Cheng, X.; Huang, W.; Cao, L. The role and mechanism of histone lactylation in health and diseases. Front. Genet. 2022, 13, 949252. [Google Scholar] [CrossRef] [PubMed]
- Yuan, K.; Gong, Y.M.; Liu, L.; Sun, Y.K.; Tian, S.S.; Wang, Y.J.; Zhong, Y.; Zhang, A.Y.; Su, S.Z.; Liu, X.X.; et al. Prevalence of posttraumatic stress disorder after infectious disease pandemics in the twenty-first century, including COVID-19: A meta-analysis and systematic review. Mol. Psychiatry 2021, 26, 4982–4998. [Google Scholar] [CrossRef]
- Sfera, A.; Osorio, C.; Rahman, L.; Zapata-Martín Del Campo, C.M.; Maldonado, J.C.; Jafri, N.; Cummings, M.A.; Maurer, S.; Kozlakidis, Z. PTSD as an Endothelial Disease: Insights From COVID-19. Front. Cell Neurosci. 2021, 15, 770387. [Google Scholar] [CrossRef]
- Israelski, D.M.; Prentiss, D.E.; Lubega, S.; Balmas, G.; Garcia, P.; Muhammad, M.; Cummings, S.; Koopman, C. Psychiatric co-morbidity in vulnerable populations receiving primary care for HIV/AIDS. AIDS Care 2007, 19, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Bah, A.J.; James, P.B.; Bah, N.; Sesay, A.B.; Sevalie, S.; Kanu, J.S. Prevalence of anxiety, depression and post-traumatic stress disorder among Ebola survivors in northern Sierra Leone: A cross-sectional study. BMC Public Health 2020, 20, 1391. [Google Scholar] [CrossRef]
- Janiri, D.; Carfì, A.; Kotzalidis, G.D.; Bernabei, R.; Landi, F.; Sani, G.; Gemelli Against COVID-19 Post-Acute Care Study Group. Posttraumatic Stress Disorder in Patients After Severe COVID-19 Infection. JAMA Psychiatry 2021, 78, 567–569. [Google Scholar] [CrossRef]
- Icard, P.; Lincet, H.; Wu, Z.; Coquerel, A.; Forgez, P.; Alifano, M.; Fournel, L. The key role of Warburg effect in SARS-CoV-2 replication and associated inflammatory response. Biochimie 2021, 180, 169–177. [Google Scholar] [CrossRef]
- Quagliato, L.A.; Freire, R.C.; Nardi, A.E. The role of acid-sensitive ion channels in panic disorder: A systematic review of animal studies and meta-analysis of human studies. Transl. Psychiatry 2018, 8, 185. [Google Scholar] [CrossRef] [PubMed]
- Hollenbeck, A.; Weber, S.; Händschke, K.; Necke, M.; Opalka, B.; Dührsen, U.; Göthert, J.R. Activation of the Hypoxia-Inducible Factor Pathway Expands Early Thymic Progenitors. Blood 2014, 124, 2896. [Google Scholar] [CrossRef]
- Tayeri, K.; Asadollahi, K.; Madani, N.; Haghjooy Javanmard, S. Does COVID-19 Escalate Aging Process? A Possible Concern. Adv. Biomed. Res. 2022, 11, 106. [Google Scholar] [CrossRef]
- Cao, X.; Li, W.; Wang, T.; Ran, D.; Davalos, V.; Planas-Serra, L.; Pujol, A.; Esteller, M.; Wang, X.; Yu, H. Accelerated biological aging in COVID-19 patients. Nat. Commun. 2022, 13, 2135. [Google Scholar] [CrossRef]
- Sfera, A.; Osorio, C.; Jafri, N.; Diaz, E.L.; Campo Maldonado, J.E. Intoxication with Endogenous Angiotensin II: A COVID-19 Hypothesis. Front. Immunol. 2020, 11, 1472. [Google Scholar] [CrossRef]
- Lee, S.; Yu, Y.; Trimpert, J.; Benthani, F.; Mairhofer, M.; Richter-Pechanska, P.; Wyler, E.; Belenki, D.; Kaltenbrunner, S.; Pammer, M.; et al. Virus-induced senescence is a driver and therapeutic target in COVID-19. Nature 2021, 599, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Mattorre, B.; Tedeschi, V.; Paldino, G.; Fiorillo, M.T.; Paladini, F.; Sorrentino, R. The emerging multifunctional roles of ERAP1, ERAP2 and IRAP between antigen processing and renin-angiotensin system modulation. Front. Immunol. 2022, 13, 1002375. [Google Scholar] [CrossRef]
- Marie, C.; Pedard, M.; Quirié, A.; Tessier, A.; Garnier, P.; Totoson, P.; Demougeot, C. Brain-derived neurotrophic factor secreted by the cerebral endothelium: A new actor of brain function? J. Cereb. Blood Flow Metab. 2018, 38, 935–949. [Google Scholar] [CrossRef] [PubMed]
- Taugher, R.J.; Lu, Y.; Fan, R.; Ghobbeh, A.; Kreple, C.J.; Faraci, F.M.; Wemmie, J.A. ASIC1A in neurons is critical for fear-related behaviors. Genes Brain Behav. 2017, 16, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Yingjun, G.; Xun, Q. Acid-sensing ion channels under hypoxia. Channels 2013, 7, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Dienel, G.A. Brain Glucose Metabolism: Integration of Energetics with Function. Physiol. Rev. 2019, 99, 949–1045. [Google Scholar] [CrossRef]
- Hayashi, T. Conversion of psychological stress into cellular stress response: Roles of the sigma-1 receptor in the process. Psychiatry Clin. Neurosci. 2015, 69, 179–191. [Google Scholar] [CrossRef]
- Hetz, C. The unfolded protein response: Controlling cell fate decisions under ER stress and beyond. Nat. Rev. Mol. Cell. Biol. 2012, 13, 89–102. [Google Scholar] [CrossRef]
- Mahendra, A.N.; Putra, I.N.A.J. Hippocampal Endoplasmic Reticulum Stress: Novel Target in PTSD Pharmacotherapy? Biomed. Pharmacol. J. 2018, 11, 1269–1274. [Google Scholar] [CrossRef]
- Ghemrawi, R.; Khair, M. Endoplasmic Reticulum Stress and Unfolded Protein Response in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 6127. [Google Scholar] [CrossRef]
- Díaz-Hung, M.L.; Martínez, G.; Hetz, C. Emerging roles of the unfolded protein response (UPR) in the nervous system: A link with adaptive behavior to environmental stress? Int. Rev. Cell Mol. Biol. 2020, 350, 29–61. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, L.; Sopontammarak, B.; Menikdiwela, K.R.; Moustaid-Moussa, N. Endoplasmic Reticulum (ER) Stress in Part Mediates Effects of Angiotensin II in Pancreatic Beta Cells. Diabetes Metab. Syndr. Obes. 2020, 13, 2843–2853. [Google Scholar] [CrossRef]
- Cicalese, S.; Cooper, H.; Preston, K.; Kawai, T.; Scalia, R.; Eguchi, S. Angiotensin II-induced ER stress response contributes to profibrotic VSMC phenotype and subsequent hypertensive arterial stiffness. J. Hypertens. 2021, 39, e40. [Google Scholar] [CrossRef]
- Su, J.B. Different cross-talk sites between the renin-angiotensin and the kallikrein-kinin systems. J. Renin. Angiotensin Aldosterone Syst. 2014, 15, 319–328. [Google Scholar] [CrossRef]
- Uddin, M.S.; Tewari, D.; Sharma, G.; Kabir, M.T.; Barreto, G.E.; Bin-Jumah, M.N.; Perveen, A.; Abdel-Daim, M.M.; Ashraf, G.M. Molecular Mechanisms of ER Stress and UPR in the Pathogenesis of Alzheimer’s Disease. Mol. Neurobiol. 2020, 57, 2902–2919. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wang, X.; Gillette, T.G.; Deng, Y.; Wang, Z.V. Unfolded Protein Response as a Therapeutic Target in Cardiovascular Disease. Curr. Top. Med. Chem. 2019, 19, 1902–1917. [Google Scholar] [CrossRef]
- Kim, P.; Scott, M.R.; Meador-Woodruff, J.H. Dysregulation of the unfolded protein response (UPR) in the dorsolateral prefrontal cortex in elderly patients with schizophrenia. Mol. Psychiatry 2021, 26, 1321–1331. [Google Scholar] [CrossRef]
- Marciniak, S.J.; Chambers, J.E.; Ron, D. Pharmacological targeting of endoplasmic reticulum stress in disease. Nat. Rev. Drug. Discov. 2022, 21, 115–140. [Google Scholar] [CrossRef] [PubMed]
- Suliman, M.; Schmidtke, M.W.; Greenberg, M.L. The Role of the UPR Pathway in the Pathophysiology and Treatment of Bipolar Disorder. Front. Cell Neurosci. 2021, 15, 735622. [Google Scholar] [CrossRef]
- Srinivasan, V.; Korhonen, L.; Lindholm, D. The Unfolded Protein Response and Autophagy as Drug Targets in Neuropsychiatric Disorders. Front. Cell Neurosci. 2020, 14, 554548. [Google Scholar] [CrossRef]
- Kudo, K.; Ohta, M.; Yang, L.; Wakasa, Y.; Takahashi, S.; Takaiwa, F. ER stress response induced by the production of human IL-7 in rice endosperm cells. Plant. Mol. Biol. 2013, 81, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Chehtane, M.; Khaled, A.R. Interleukin-7 mediates glucose utilization in lymphocytes through transcriptional regulation of the hexokinase II gene. Am. J. Physiol. Cell Physiol. 2010, 298, C1560–C1571. [Google Scholar] [CrossRef] [PubMed]
- Bekele, Y.; Sui, Y. Berzofsky JA. IL-7 in SARS-CoV-2 Infection and as a Potential Vaccine Adjuvant. Front. Immunol. 2021, 12, 737406. [Google Scholar] [CrossRef]
- Tang, C.; Goldsamt, L.; Meng, J.; Xiao, X.; Zhang, L.; Williams, A.B.; Wang, H. Global estimate of the prevalence of post-traumatic stress disorder among adults living with HIV: A systematic review and meta-analysis. BMJ Open 2020, 10, e032435. [Google Scholar] [CrossRef]
- Kaputu-Kalala-Malu, C.; Musalu, E.M.; Walker, T.; Ntumba-Tshitenge, O.; Ahuka-Mundeke, S. PTSD, depression and anxiety in Ebola virus disease survivors in Beni town, Democratic Republic of the Congo. BMC Psychiatry 2021, 21, 342. [Google Scholar] [CrossRef]
- Silva, S.L.; Albuquerque, A.S.; Matoso, P.; Charmeteau-de-Muylder, B.; Cheynier, R.; Ligeiro, D.; Abecasis, M.; Anjos, R.; Barata, J.T.; Victorino, R.M.; et al. IL-7-Induced Proliferation of Human Naive CD4 T-Cells Relies on Continued Thymic Activity. Front. Immunol. 2017, 8, 20. [Google Scholar] [CrossRef]
- Zheng, S.; Han, F.; Shi, Y.; Wen, L.; Han, D. Single-Prolonged-Stress-Induced Changes in Autophagy-Related Proteins Beclin-1, LC3, and p62 in the Medial Prefrontal Cortex of Rats with Post-traumatic Stress Disorder. J. Mol. Neurosci. 2017, 62, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Jiang, X.; Ding, Z.; Ming, J. Interleukin 7 inhibit autophagy via P53 regulated AMPK/mTOR signaling pathway in non-small cell lung cancer. Sci. Rep. 2022, 12, 11208. [Google Scholar] [CrossRef] [PubMed]
- Shehata, M.; Abdou, K.; Choko, K.; Matsuo, M.; Nishizono, H.; Inokuchi, K. Autophagy Enhances Memory Erasure through Synaptic Destabilization. J. Neurosci. 2018, 38, 3809–3822. [Google Scholar] [CrossRef]
- Abdellatif, M.; Ljubojevic-Holzer, S.; Madeo, F.; Sedej, S. Autophagy in cardiovascular health and disease. Prog. Mol. Biol. Transl. Sci. 2020, 172, 87–106. [Google Scholar] [CrossRef]
- Vranova, M.; Friess, M.C.; Haghayegh Jahromi, N.; Collado-Diaz, V.; Vallone, A.; Hagedorn, O.; Halin, C. Opposing roles of endothelial and leukocyte-expressed IL-7Rα in the regulation of psoriasis-like skin inflammation. Sci. Rep. 2019, 9, 11714. [Google Scholar] [CrossRef]
- Saric, T.; Chang, S.C.; Hattori, A.; York, I.A.; Markant, S.; Rock, K.L.; Tsujimoto, M.; Goldberg, A.L. An IFN-gamma-induced aminopeptidase in the ER, ERAP1, trims precursors to MHC class I-presented peptides. Nat. Immunol. 2002, 3, 1169–1176. [Google Scholar] [CrossRef]
- Zee, R.Y.L.; Rivera, A.; Inostroza, Y.; Ridker, P.M.; Chasman, D.I.; Romero, J.R. Gene Variation of Endoplasmic Reticulum Aminopeptidases 1 and 2, and Risk of Blood Pressure Progression and Incident Hypertension among 17,255 Initially Healthy Women. Int. J. Genom. 2018, 2018, 2308585. [Google Scholar] [CrossRef] [PubMed]
- Cochran, D.M.; Fallon, D.; Hill, M.; Frazier, J.A. The role of oxytocin in psychiatric disorders: A review of biological and therapeutic research findings. Harv. Rev. Psychiatry 2013, 21, 219–247. [Google Scholar] [CrossRef] [PubMed]
- Frijling, J.L. Preventing PTSD with oxytocin: Effects of oxytocin administration on fear neurocircuitry and PTSD symptom development in recently trauma-exposed individuals. Eur. J. Psychotraumatol. 2017, 8, 1302652. [Google Scholar] [CrossRef]
- Martínez-Martos, J.M.; Correa-Rodríguez, M.; Rus, A.; Molina, F.; Ramírez-Expósito, M.J.; Aguilar-Ferrandiz, M.E. Altered Serum Oxytocinase and Enkephalin-Degrading aminopeptidase activities in patients with Fibromyalgia. Biol. Res. Nurs. 2019, 21, 431–439. [Google Scholar] [CrossRef] [PubMed]
- López de Castro, J.A.; Alvarez-Navarro, C.; Brito, A.; Guasp, P.; Martín-Esteban, A.; Sanz-Bravo, A. Molecular and pathogenic effects of endoplasmic reticulum aminopeptidases ERAP1 and ERAP2 in MHC-i-associated inflammatory disorders: Towards a unifying view. Mol. Immunol. 2016, 77, 193–204. [Google Scholar] [CrossRef]
- Gabrielsen, I.; Viken, M.; Amundsen, S.S.; Helgeland, H.; Holm, K.; Flåm, S.T.; Lie, B.A. Autoimmune risk variants in ERAP2 are associated with gene-expression levels in thymus. Genes Immun. 2016, 17, 406–411. [Google Scholar] [CrossRef]
- Liew, J.; Lucas Williams, J.; Dobscha, S.; Barton, J.L. Posttraumatic stress disorder and correlates of disease activity among veterans with ankylosing spondylitis. Rheumatol. Int. 2017, 37, 1765–1769. [Google Scholar] [CrossRef]
- Qureshi, S.U.; Pyne, J.M.; Magruder, K.M.; Schulz, P.E.; Kunik, M.E. The link between post-traumatic stress disorder and physical comorbidities: A systematic review. Psychiatry Q. 2009, 80, 87–97. [Google Scholar] [CrossRef]
- Beyer, C.E.; Dwyer, J.M.; Platt, B.J.; Neal, S.; Luo, B.; Ling, H.P.; Lin, Q.; Mark, R.J.; Rosenzweig-Lipson, S.; Schechter, L.E. Angiotensin IV elevates oxytocin levels in the rat amygdala and produces anxiolytic-like activity through subsequent oxytocin receptor activation. Psychopharmacology 2010, 209, 303–311. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, S.; Tempora, P.; Lucarini, V.; Melaiu, O.; Gaspari, S.; Algeri, M.; Fruci, D. ERAP1 and ERAP2 Enzymes: A Protective Shield for RAS against COVID-19? Int. J. Mol. Sci. 2021, 22, 1705. [Google Scholar] [CrossRef] [PubMed]
- Saulle, I.; Vanetti, C.; Goglia, S.; Vicentini, C.; Tombetti, E.; Garziano, M.; Clerici, M.; Biasin, M. A New ERAP2/Iso3 Isoform Expression Is Triggered by Different Microbial Stimuli in Human Cells. Could It Play a Role in the Modulation of SARS-CoV-2 Infection? Cells 2020, 9, 1951. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.J.; Chen, J.; Villani, A.C.; Gate, R.E.; Subramaniam, M.; Bhangale, T.; Lee, M.N.; Raj, T.; Raychowdhury, R.; Li, W.; et al. Genetic analysis of isoform usage in the human anti-viral response reveals influenza-specific regulation of ERAP2 transcripts under balancing selection. Genome Res. 2018, 28, 1812–1825. [Google Scholar] [CrossRef]
- Gracia-Ramos, A.E.; Martin-Nares, E.; Hernández-Molina, G. New Onset of Autoimmune Diseases Following COVID-19 Diagnosis. Cells 2021, 10, 3592. [Google Scholar] [CrossRef]
- Liu, S.; Cao, D.; Shen, Y.; Li, Y.; Li, Y.; Shi, L.; Yu, J.; Li, C.; Zhang, X.; Sun, M.; et al. The ERAP gene is associated with HCV chronic infection in a Chinese Han population. Hum. Immunol. 2017, 78, 731–738. [Google Scholar] [CrossRef]
- Klunk, J.; Vilgalys, T.P.; Demeure, C.E. Evolution of immune genes is associated with the Black Death. Nature 2022, 611, 312–319. [Google Scholar] [CrossRef]
- Chang, Y.; Wei, W. Angiotensin II in inflammation, immunity and rheumatoid arthritis. Clin. Exp. Immunol. 2015, 179, 137–145. [Google Scholar] [CrossRef]
- Ferreira, L.C.; Gomes, C.E.M.; Duggal, P.; De Paula Holanda, I.; de Lima, A.S.; do Nascimento, P.R.P.; Jeronimo, S.M.B. Genetic association of ERAP1 and ERAP2 with eclampsia and preeclampsia in northeastern Brazilian women. Sci. Rep. 2021, 11, 6764. [Google Scholar] [CrossRef]
- Gelfand, J.M.; Wang, S.; Garshick, M.S. Autoimmune diseases and cardiovascular risk. Trends Mol. Med. 2022, 28, 1025–1027. [Google Scholar] [CrossRef]
- Song, H.; Fang, F.; Tomasson, G.; Arnberg, F.K.; Mataix-Cols, D.; Fernández de la Cruz, L.; Almqvist, C.; Fall, K.; Valdimarsdóttir, U.A. Association of Stress-Related Disorders with Subsequent Autoimmune Disease. JAMA 2018, 319, 2388–2400. [Google Scholar] [CrossRef]
- Blake, M.K.; O’Connell, P.; Pepelyayeva, Y.; Godbehere, S.; Aldhamen, Y.A.; Amalfitano, A. ERAP1 is a critical regulator of inflammasome-mediated proinflammatory and ER stress responses. BMC Immunol. 2022, 23, 9. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.; Nakata, K.; Sagara, A.; Iwamoto, C.; Endo, S.; Matsuda, R.; Matsumoto, S.; Ikenaga, N.; Shindo, K.; Moriyama, T.; et al. ERAP2 is a novel target involved in autophagy and activation of pancreatic stellate cells via UPR signaling pathway. Pancreatology 2022, 22, 9–19. [Google Scholar] [CrossRef]
- Ren, J.; Bi, Y.; Sowers, J.R.; Hetz, C.; Zhang, Y. Endoplasmic reticulum stress and unfolded protein response in cardiovascular diseases. Nat. Rev. Cardiol. 2021, 18, 499–521. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Yuan, Y.; Zou, Q. The role and mechanism of gastrodin in the medial prefrontal cortex autophagy of PTSD rats. Int. J. Clin. Exp. Pathol. 2020, 13, 989–994. [Google Scholar] [PubMed]
- Martin-Du Pan, R.C. L’ocytocine: Hormone de l’amour, de la confiance et du lien conjugal et social [Oxytocin: The hormone of love, trust and social bond. Clinical use in autism and social phobia]. Rev. Med. Suisse 2012, 8, 627–630. (In French) [Google Scholar]
- Qi, J.; Han, W.Y.; Yang, J.Y.; Wang, L.H.; Dong, Y.X.; Wang, F.; Song, M.; Wu, C.F. Oxytocin regulates changes of extracellular glutamate and GABA levels induced by methamphetamine in the mouse brain. Addict. Biol. 2012, 17, 758–769. [Google Scholar] [CrossRef]
- Love, T.M. Oxytocin, motivation and the role of dopamine. Pharmacol. Biochem. Behav. 2014, 119, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Krimberg, J.S.; Lumertz, F.S.; Orso, R.; Viola, T.W.; de Almeida, R.M.M. Impact of social isolation on the oxytocinergic system: A systematic review and meta-analysis of rodent data. Neurosci. Biobehav. Rev. 2022, 134, 104549. [Google Scholar] [CrossRef]
- Engel, S.; Klusmann, H.; Laufer, S.; Pfeifer, A.C.; Ditzen, B.; van Zuiden, M.; Knaevelsrud, C.; Schumacher, S. Trauma exposure, posttraumatic stress disorder and oxytocin: A meta-analytic investigation of endogenous concentrations and receptor genotype. Neurosci. Biobehav. Rev. 2019, 107, 560–601. [Google Scholar] [CrossRef]
- Valtorta, N.K.; Kanaan, M.; Gilbody, S.; Hanratty, B. Loneliness, social isolation and risk of cardiovascular disease in the English Longitudinal Study of Ageing. Eur. J. Prev. Cardiol. 2018, 25, 1387–1396. [Google Scholar] [CrossRef] [PubMed]
- Golaszewski, N.M.; LaCroix, A.Z.; Godino, J.G.; Allison, M.A.; Manson, J.E.; King, J.J.; Weitlauf, J.C.; Bea, J.W.; Garcia, L.; Kroenke, C.H.; et al. Evaluation of Social Isolation, Loneliness, and Cardiovascular Disease Among Older Women in the US. JAMA Netw. Open 2022, 5, e2146461. [Google Scholar] [CrossRef]
- Wang, C.; Pan, R.; Wan, X.; Tan, Y.; Xu, L.; McIntyre, R.S.; Choo, F.N.; Tran, B.; Ho, R.; Sharma, V.K.; et al. A longitudinal study on the mental health of general population during the COVID-19 epidemic in China. Brain Behav. Immun. 2020, 87, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Gryksa, K.; Neumann, I.D. Consequences of pandemic-associated social restrictions: Role of social support and the oxytocin system. Psychoneuroendocrinology 2022, 135, 105601. [Google Scholar] [CrossRef] [PubMed]
- Erdman, S.E.; Poutahidis, T. Microbes and Oxytocin: Benefits for Host Physiology and Behavior. Int. Rev. Neurobiol. 2016, 131, 91–126. [Google Scholar] [CrossRef] [PubMed]
- Varian, B.J.; Poutahidis, T.; DiBenedictis, B.T.; Levkovich, T.; Ibrahim, Y.; Didyk, E.; Shikhman, L.; Cheung, H.K.; Hardas, A.; Ricciardi, C.E.; et al. Microbial lysate upregulates host oxytocin. Brain Behav. Immun. 2017, 61, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Kang, I.; Kondo, D.; Kim, J.; Lyoo, I.K.; Yurgelun-Todd, D.; Hwang, J.; Renshaw, P.F. Elevating the level of hypoxia inducible factor may be a new potential target for the treatment of depression. Med. Hypotheses 2021, 146, 110398. [Google Scholar] [CrossRef]
- Doenyas-Barak, K.; Catalogna, M.; Kutz, I.; Levi, G.; Hadanny, A.; Tal, S.; Daphna-Tekoha, S.; Sasson, E.; Shechter, Y.; Efrati, S. Hyperbaric oxygen therapy improves symptoms, brain’s microstructure and functionality in veterans with treatment resistant post-traumatic stress disorder: A prospective, randomized, controlled trial. PLoS ONE 2022, 17, e0264161. [Google Scholar] [CrossRef]
- Ding, F.S.; Cheng, X.; Zhao, T.; Zhao, Y.Q.; Zhang, G.B.; Wu, H.T.; Wu, K.W. Intermittent hypoxic preconditioning relieves fear and anxiety behavior in post-traumatic stress model mice. Sheng Li Xue Bao Acta Physiol. Sin. 2019, 71, 537–546. [Google Scholar]
- Aparci, M.; Kardesoglu, E.; Suleymanoglu, S.; Uzun, G.; Onem, Y.; Uz, O.; Kucukardali, Y.; Ozkan, S. Hyperbaric oxygen therapy improves myocardial diastolic function in diabetic patients. Tohoku. J. Exp. Med. 2008, 214, 281–289. [Google Scholar] [CrossRef]
- Hesse, J.; Groterath, W.; Owenier, C.; Steinhausen, J.; Ding, Z.; Steckel, B.; Czekelius, C.; Alter, C.; Marzoq, A.; Schrader, J. Normoxic induction of HIF-1α by adenosine-A2B R signaling in epicardial stromal cells formed after myocardial infarction. FASEB J. 2021, 35, e21517. [Google Scholar] [CrossRef]
- Gnad, T.; Navarro, G.; Lahesmaa, M.; Reverte-Salisa, L.; Copperi, F.; Cordomi, A.; Naumann, J.; Hochhäuser, A.; Haufs-Brusberg, S.; Wenzel, D.; et al. Adenosine/A2B Receptor Signaling Ameliorates the Effects of Aging and Counteracts Obesity. Cell Metab. 2020, 32, 56–70.e7. [Google Scholar] [CrossRef] [PubMed]
- Kotańska, M.; Dziubina, A.; Szafarz, M.; Mika, K.; Bednarski, M.; Nicosia, N.; Temirak, A.; Müller, C.E.; Kieć-Kononowicz, K. Preliminary Evidence of the Potent and Selective Adenosine A2B Receptor Antagonist PSB-603 in Reducing Obesity and Some of Its Associated Metabolic Disorders in Mice. Int. J. Mol. Sci. 2022, 23, 13439. [Google Scholar] [CrossRef]
- Fisher, J.W.; Brookins, J. Adenosine A(2A) and A(2B) receptor activation of erythropoietin production. Am. J. Physiol. Renal. Physiol. 2001, 281, F826–F832. [Google Scholar] [CrossRef] [PubMed]
- Sampath, D.; McWhirt, J.; Sathyanesan, M.; Newton, S.S. Carbamoylated erythropoietin produces antidepressant-like effects in male and female mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 96, 109754. [Google Scholar] [CrossRef] [PubMed]
- Stankowska, D.L.; Mueller, B.H., 2nd; Oku, H.; Ikeda, T.; Dibas, A. Neuroprotective effects of inhibitors of Acid-Sensing ion channels (ASICs) in optic nerve crush model in rodents. Curr. Eye. Res. 2018, 43, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zheng, L.; Yang, X.; Yao, S.; Wang, H.; An, J.; Jin, H.; Wen, G.; Tuo, B. Pathology and physiology of acid-sensitive ion channels in the digestive system (Review). Int. J. Mol. Med. 2022, 50, 94. [Google Scholar] [CrossRef]
- Dwyer, J.M.; Rizzo, S.J.; Neal, S.J.; Lin, Q.; Jow, F.; Arias, R.L.; Rosenzweig-Lipson, S.; Dunlop, J.; Beyer, C.E. Acid sensing ion channel (ASIC) inhibitors exhibit anxiolytic-like activity in preclinical pharmacological models. Psychopharmacology 2009, 203, 41–52. [Google Scholar] [CrossRef]
- Gu, Q.; Lee, L.Y. Acid-Sensing Ion Channels and Pain. Pharmaceuticals 2010, 3, 1411–1425. [Google Scholar] [CrossRef]
- Arias, R.L.; Sung, M.L.; Vasylyev, D.; Zhang, M.Y.; Albinson, K.; Kubek, K.; Kagan, N.; Beyer, C.; Lin, Q.; Dwyer, J.M.; et al. Amiloride is neuroprotective in an MPTP model of Parkinson’s disease. Neurobiol. Dis. 2008, 31, 334–341. [Google Scholar] [CrossRef]
- Ziemann, A.E.; Allen, J.E.; Dahdaleh, N.S.; Drebot, I.I.; Coryell, M.W.; Wunsch, A.M.; Lynch, C.M.; Faraci, F.M.; Howard, M.A., 3rd; Welsh, M.J.; et al. The amygdala is a chemosensor that detects carbon dioxide and acidosis to elicit fear behavior. Cell 2009, 139, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Smoller, J.W.; Gallagher, P.J.; Duncan, L.E.; McGrath, L.M.; Haddad, S.A.; Holmes, A.J.; Wolf, A.B.; Hilker, S.; Block, S.R.; Weill, S.; et al. The human ortholog of acid-sensing ion channel gene ASIC1a is associated with panic disorder and amygdala structure and function. Biol. Psychiatry 2014, 76, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Bhagatwala, J.; Harris, R.A.; Parikh, S.J.; Zhu, H.; Huang, Y.; Kotak, I.; Seigler, N.; Pierce, G.L.; Egan, B.M.; Dong, Y. Epithelial sodium channel inhibition by amiloride on blood pressure and cardiovascular disease risk in young prehypertensives. J. Clin. Hypertens. 2014, 16, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, M.; Rossignol, O.; Bachand, K.; D’Amato, F.R.; De Koninck, Y. Amiloride modulation of carbon dioxide hypersensitivity and thermal nociceptive hypersensitivity induced by interference with early maternal environment. J. Psychopharmacol. 2019, 33, 101–108. [Google Scholar] [CrossRef]
- Qi, X.; Lu, J.F.; Huang, Z.Y.; Liu, Y.J.; Cai, L.B.; Wen, X.L.; Song, X.L.; Xiong, J.; Sun, P.Y.; Zhang, H.; et al. Pharmacological Validation of ASIC1a as a Druggable Target for Neuroprotection in Cerebral Ischemia Using an Intravenously Available Small Molecule Inhibitor. Front. Pharmacol. 2022, 13, 849498. [Google Scholar] [CrossRef]
- Osmakov, D.I.; Koshelev, S.G.; Belozerova, O.A.; Kublitski, V.S.; Andreev, Y.A.; Grishin, E.V.; Kozlov, S.A. The Biological Activity of the Sevanol and Its Analogues. Bioorg. Khim. 2015, 41, 606–611, In Russian. [Google Scholar] [CrossRef]
- Belozerova, O.A.; Osmakov, D.I.; Vladimirov, A.; Koshelev, S.G.; Chugunov, A.O.; Andreev, Y.A.; Palikov, V.A.; Palikova, Y.A.; Shaykhutdinova, E.R.; Gvozd, A.N.; et al. Sevanol and Its Analogues: Chemical Synthesis, Biological Effects and Molecular Docking. Pharmaceuticals 2020, 13, 163. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.L.; Cheng, C.F.; Sun, W.H.; Wong, C.W.; Chen, C.C. Targeting ASIC3 for pain, anxiety, and insulin resistance. Pharmacol. Ther. 2012, 134, 127–138. [Google Scholar] [CrossRef]
- Yan, X.G.; Li, W.G.; Qi, X.; Zhu, J.J.; Huang, C.; Han, S.L.; Jiang, Q.; Xu, T.L.; Liu, J.H. Subtype-selective inhibition of acid-sensing ion channel 3 by a natural flavonoid. CNS Neurosci. Ther. 2019, 25, 47–56. [Google Scholar] [CrossRef]
- Gu, J.; Zhou, J.; Chen, Q.; Xu, X.; Gao, J.; Li, X.; Shao, Q.; Zhou, B.; Zhou, H.; Wei, S.; et al. Tumor metabolite lactate promotes tumorigenesis by modulating MOESIN lactylation and enhancing TGF-β signaling in regulatory T cells. Cell Rep. 2022, 39, 110986. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, A.; Mitsuhara, Y.; Orimoto, A.; Nakayasu, Y.; Tsunoda, S.; Obana, M.; Maeda, M.; Nakayama, H.; Yoshioka, Y.; Tsutsumi, Y.; et al. Moesin is activated in cardiomyocytes in experimental autoimmune myocarditis and mediates cytoskeletal reorganization with protrusion formation. Am. J. Physiol. Heart. Circ. Physiol. 2016, 311, H476–H486. [Google Scholar] [CrossRef]
- Freymuth, P.S.; Fitzsimons, H.L. The ERM protein Moesin is essential for neuronal morphogenesis and long-term memory in Drosophila. Mol. Brain 2017, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Swartz, C.M.; Breen, K.J. Multiple muscle enzyme release with psychiatric illness. J. Nerv. Ment. Dis. 1990, 178, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Ma, Y.; Guo, W.; Lu, J.; Li, X.; Wu, J.; Qin, P.; Zhu, C.; Zhang, Q. Serum Level of Lactate Dehydrogenase is Associated with Cardiovascular Disease Risk as Determined by the Framingham Risk Score and Arterial Stiffness in a Health-Examined Population in China. Int. J. Gen. Med. 2022, 15, 11–17. [Google Scholar] [CrossRef]
- Ivanova, T.M.; Belova, T.I. Effect of emotional stress on lactate dehydrogenase isozyme spectrum in the rat reticular formation. Bull. Exp. Biol. Med. 1986, 102, 1331–1334. [Google Scholar] [CrossRef]
- Kato, A.; Sakakibara, H.; Tsuboi, H.; Tatsumi, A.; Akimoto, M.; Shimoi, K.; Ohashi, N. Depressive symptoms of female nursing staff working in stressful environments and their association with serum creatine kinase and lactate dehydrogenase—A preliminary study. BioPsychoSocial Med. 2014, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Padoli, P.; Suwito, J.; Hariyanto, T. Self-Affirmation Reduces the Anxiety, LDH and Troponin I in the Clients with Coronary Heart Disease (CHD). J. Ners. 2020, 14, 310–315. [Google Scholar] [CrossRef]
- Higashikawa, F.; Kanno, K.; Ogata, A.; Sugiyama, M. Reduction of fatigue and anger-hostility by the oral administration of 5-aminolevulinic acid phosphate: A randomized, double-blind, placebo-controlled, parallel study. Sci. Rep. 2020, 10, 16004. [Google Scholar] [CrossRef]
- Perez, M.; Shintani, T.; Rodriguez, B.; Davis, J.; Harrigan, R. The Role of 5-Aminolevulinic Acid (5-ALA) and Sleep. Int. J. Clin. Med. 2013, 4, 1–7. [Google Scholar] [CrossRef]
- Hagisawa, K.; Ayaori, M.; Ikewaki, K.; Nakajima, M.; Morimoto, Y. 5-Aminolevulinic Acid Attenuates Atherosclerotic Plaque Progression in Low-Density Lipoprotein Receptor-Deficient Mice by Heme Oxygenase-1 Induction. Circ. Rep. 2019, 2, 60–68. [Google Scholar] [CrossRef]
- Mohammed Abdul, K.S.; Jovanović, S.; Du, Q.; Sukhodub, A.; Jovanović, A. Mild hypoxia in vivo regulates cardioprotective SUR2A: A role for Akt and LDH. Biochim. Biophys. Acta 2015, 1852, 709–719. [Google Scholar] [CrossRef]
- Jafary, F.; Ganjalikhany, M.R.; Moradi, A.; Hemati, M.; Jafari, S. Novel Peptide Inhibitors for Lactate Dehydrogenase A (LDHA): A Survey to Inhibit LDHA Activity via Disruption of Protein-Protein Interaction. Sci. Rep. 2019, 9, 4686. [Google Scholar] [CrossRef] [PubMed]
- Matada, B.S.; Pattanashettar, R.; Yernale, N.G. A comprehensive review on the biological interest of quinoline and its derivatives. Bioorg. Med. Chem. 2021, 32, 115973. [Google Scholar] [CrossRef] [PubMed]
- Zajdel, P.; Marciniec, K.; Maślankiewicz, A.; Grychowska, K.; Satała, G.; Duszyńska, B.; Lenda, T.; Siwek, A.; Nowak, G.; Partyka, A.; et al. Antidepressant and antipsychotic activity of new quinoline- and isoquinoline-sulfonamide analogs of aripiprazole targeting serotonin 5-HT1A/5-HT2A/5-HT7 and dopamine D2/D3 receptors. Eur. J. Med. Chem. 2013, 60, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Britnell, S.R.; Jackson, A.D.; Brown, J.N.; Capehart, B.P. Aripiprazole for Post-traumatic Stress Disorder: A Systematic Review. Clin. Neuropharmacol. 2017, 40, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.D.; Fikretoglu, D.; Liu, A.; McIntosh, D. Aripiprazole augmentation in the treatment of military-related PTSD with major depression: A retrospective chart review. BMC Psychiatry 2011, 11, 86. [Google Scholar] [CrossRef]
- Pan, L.; Feng, F.; Wu, J.; Fan, S.; Han, J.; Wang, S.; Yang, L.; Liu, W.; Wang, C.; Xu, K. Demethylzeylasteral targets lactate by inhibiting histone lactylation to suppress the tumorigenicity of liver cancer stem cells. Pharmacol. Res. 2022, 181, 106270. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Dong, Y.; Jin, X.; Zhang, C.; Zhang, T.; Zhao, J.; Shi, J.; Li, J. The novel and potent anti-depressive action of triptolide and its influences on hippocampal neuroinflammation in a rat model of depression comorbidity of chronic pain. Brain Behav. Immun. 2017, 64, 180–194. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cao, Y.; Zhang, W.; Bergmeier, S.; Qian, Y.; Akbar, H.; Colvin, R.; Ding, J.; Tong, L.; Wu, S.; et al. A small-molecule inhibitor of glucose transporter 1 downregulates glycolysis, induces cell-cycle arrest, and inhibits cancer cell growth in vitro and in vivo. Mol. Cancer. Ther. 2012, 11, 1672–1682. [Google Scholar] [CrossRef] [PubMed]
- Koch, H.; Weber, Y.G. The glucose transporter type 1 (Glut1) syndromes. Epilepsy. Behav. 2019, 91, 90–93. [Google Scholar] [CrossRef]
- Gulbins, A.; Schumacher, F.; Becker, K.A.; Wilker, B.; Soddemann, M.; Boldrin, F.; Müller, C.P.; Edwards, M.J.; Goodman, M.; Caldwell, C.C.; et al. Antidepressants act by inducing autophagy controlled by sphingomyelin-ceramide. Mol. Psychiatry 2018, 23, 2324–2346. [Google Scholar] [CrossRef] [PubMed]
- Gassen, N.C.; Rein, T. Is There a Role of Autophagy in Depression and Antidepressant Action? Front. Psychiatry 2019, 10, 337. [Google Scholar] [CrossRef] [PubMed]
- Alcocer-Gómez, E.; Casas-Barquero, N.; Williams, M.R.; Romero-Guillena, S.L.; Cañadas-Lozano, D.; Bullón, P.; Sánchez-Alcazar, J.A.; Navarro-Pando, J.M.; Cordero, M.D. Antidepressants induce autophagy dependent-NLRP3-inflammasome inhibition in Major depressive disorder. Pharmacol. Res. 2017, 121, 114–121. [Google Scholar] [CrossRef]
- Lyu, D.; Wang, F.; Zhang, M.; Yang, W.; Huang, H.; Huang, Q.; Wu, C.; Qian, N.; Wang, M.; Zhang, H.; et al. Ketamine induces rapid antidepressant effects via the autophagy-NLRP3 inflammasome pathway. Psychopharmacology 2022, 239, 3201–3212. [Google Scholar] [CrossRef]
- Hill, L.D.; Hilliard, D.D.; York, T.P.; Srinivas, S.; Kusanovic, J.P.; Gomez, R.; Strauss, J.F. Fetal ERAP2 variation is associated with preeclampsia in African Americans in a case-control study. BMC Med. Genet. 2011, 12, 64. [Google Scholar] [CrossRef] [PubMed]
- Wolfarth, A.A.; Dhar, S.; Goon, J.B.; Ezeanya, U.I.; Ferrando-Martínez, S.; Lee, B.H. Advancements of Common Gamma-Chain Family Cytokines in Cancer Immunotherapy. Immune. Netw. 2022, 22, e5. [Google Scholar] [CrossRef]
- Lee, S.W.; Choi, D.; Heo, M.; Shin, E.C.; Park, S.H.; Kim, S.J.; Oh, Y.K.; Lee, B.H.; Yang, S.H.; Sung, Y.C.; et al. hIL-7-hyFc, A Long-Acting IL-7, Increased Absolute Lymphocyte Count in Healthy Subjects. Clin. Transl. Sci. 2020, 13, 1161–1169. [Google Scholar] [CrossRef]
- Yang, Y.; An, J.; Weng, N.P. Telomerase is involved in IL-7-mediated differential survival of naive and memory CD4+ T cells. J. Immunol. 2008, 180, 3775–3781. [Google Scholar] [CrossRef]
- Li, H.; Hong, W.; Jin, X.; Li, G.; Zhou, G.; Fan, L. The aryl hydrocarbon receptor is a novel negative regulator of interleukin-17-mediated signaling and inflammation in vitro. FEBS Lett. 2019, 593, 952–961. [Google Scholar] [CrossRef]
- Rothhammer, V.; Quintana, F.J. The aryl hydrocarbon receptor: An environmental sensor integrating immune responses in health and disease. Nat. Rev. Immunol. 2019, 19, 184–197. [Google Scholar] [CrossRef]
- Cole, J.W.; Xu, H. Aryl Hydrocarbon Receptor Repressor Methylation: A Link Between Smoking and Atherosclerosis. Circ. Cardiovasc. Genet. 2015, 8, 640–642. [Google Scholar] [CrossRef]
- Smith, A.K.; Ratanatharathorn, A.; Maihofer, A.X.; Naviaux, R.K.; Aiello, A.E.; Amstadter, A.B.; Ashley-Koch, A.E.; Baker, D.G.; Beckham, J.C.; Boks, M.P.; et al. Epigenome-wide meta-analysis of PTSD across 10 military and civilian cohorts identifies methylation changes in AHRR. Nat. Commun. 2020, 11, 5965. [Google Scholar] [CrossRef]
- Sakai, M.; Yu, Z.; Hirayama, R.; Nakasato, M.; Kikuchi, Y.; Ono, C.; Komatsu, H.; Nakanishi, M.; Yoshii, H.; Stellwagen, D.; et al. Deficient Autophagy in Microglia Aggravates Repeated Social Defeat Stress-Induced Social Avoidance. Neural. Plast. 2022, 2022, 7503553. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Thompson, M.D.; Cohen, R.A.; Tong, X. Autophagy and oxidative stress in cardiovascular diseases. Biochim. Biophys. Acta. 2015, 1852, 243–251. [Google Scholar] [CrossRef]
- Lee, B.; Sur, B.; Yeom, M.; Shim, I.; Lee, H.; Hahm, D.H. Gastrodin reversed the traumatic stress-induced depressed-like symptoms in rats. J. Nat. Med. 2016, 70, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, L.; Yu, J.; Asare, P.F.; Zhao, Y.Q. Gastrodin Reduces Blood Pressure by Intervening with RAAS and PPARγ in SHRs. Evid. Based. Complement. Alternat. Med. 2015, 2015, 828427. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Yang, P.; Xie, L.; Pu, Y.; Guo, J.; Jiao, J.; Sun, L.; Lu, D. Gastrodin induces lysosomal biogenesis and autophagy to prevent the formation of foam cells via AMPK-FoxO1-TFEB signalling axis. J. Cell Mol. Med. 2021, 25, 5769–5781. [Google Scholar] [CrossRef]
- Fenizia, C.; Galbiati, S.; Vanetti, C.; Vago, R.; Clerici, M.; Tacchetti, C.; Daniele, T. SARS-CoV-2 Entry: At the Crossroads of CD147 and ACE2. Cells 2021, 10, 1434. [Google Scholar] [CrossRef]
- van Ooij, C. Basigin opens the door to malaria. Nat. Rev. Microbiol. 2012, 10, 3. [Google Scholar] [CrossRef]
- Hashimoto, T.; Hussien, R.; Brooks, G.A. Colocalization of MCT1, CD147, and LDH in mitochondrial inner membrane of L6 muscle cells: Evidence of a mitochondrial lactate oxidation complex. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E1237–E1244. [Google Scholar] [CrossRef]
- Wang, P.; Wang, Z.; Yan, Y.; Xiao, L.; Tian, W.; Qu, M.; Meng, A.; Sun, F.; Li, G.; Dong, J. Psychological Stress Up-Regulates CD147 Expression Through Beta-Arrestin1/ERK to Promote Proliferation and Invasiveness of Glioma Cells. Front. Oncol. 2020, 10, 571181. [Google Scholar] [CrossRef]
- Seizer, P.; Gawaz, M.; May, A.E. Cyclophilin A and EMMPRIN (CD147) in cardiovascular diseases. Cardiovasc. Res. 2014, 102, 17–23. [Google Scholar] [CrossRef]
- Chen, R.; Wang, K.; Feng, Z.; Zhang, M.Y.; Wu, J.; Geng, J.J.; Chen, Z.N. CD147 deficiency in T cells prevents thymic involution by inhibiting the EMT process in TECs in the presence of TGFβ. Cell Mol. Immunol. 2021, 18, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Ke, X.; Fei, F.; Chen, Y.; Xu, L.; Zhang, Z.; Huang, Q.; Zhang, H.; Yang, H.; Chen, Z.; Xing, J. Hypoxia upregulates CD147 through a combined effect of HIF-1α and Sp1 to promote glycolysis and tumor progression in epithelial solid tumors. Carcinogenesis 2012, 33, 1598–1607. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, A.G. Anaesthetic and other treatments of shell shock: World War I and beyond. J. R Army Med. Corps. 2012, 158, 29–33. [Google Scholar] [CrossRef]
- Du, W.; Ren, L.; Hamblin, M.H.; Fan, Y. Endothelial Cell Glucose Metabolism and Angiogenesis. Biomedicines 2021, 9, 147. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Zhang, R.; Xu, Z.; Ke, Y.; Sun, R.; Yang, H.; Zheng, L.T. Early glycolytic reprogramming controls microglial inflammatory activation. J. Neuroinflamm. 2021, 18, 129. [Google Scholar] [CrossRef] [PubMed]
- Mason, S. Lactate Shuttles in Neuroenergetics-Homeostasis, Allostasis and Beyond. Front. Neurosci. 2017, 11, 43. [Google Scholar] [CrossRef]
- Goyal, M.S.; Vlassenko, A.G.; Blazey, T.M.; Su, Y.; Couture, L.E.; Durbin, T.J.; Bateman, R.J.; Benzinger, T.L.; Morris, J.C.; Raichle, M.E. Loss of Brain Aerobic Glycolysis in Normal Human Aging. Cell Metab. 2017, 26, 353–360.e3. [Google Scholar] [CrossRef]
- Hughes, K.C.; Shin, L.M. Functional neuroimaging studies of post-traumatic stress disorder. Expert Rev. Neurother. 2011, 11, 275–285. [Google Scholar] [CrossRef]
- Michopoulos, V.; Vester, A.; Neigh, G. Posttraumatic stress disorder: A metabolic disorder in disguise? Exp. Neurol. 2016, 284, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Caughron, B.; Young, M.R.I. Posttraumatic Stress Disorder: An Immunological Disorder? Front. Psychiatry 2017, 8, 222. [Google Scholar] [CrossRef]
- Boscarino, J.A. Posttraumatic stress disorder and physical illness: Results from clinical and epidemiologic studies. Ann. N. Y. Acad. Sci. 2004, 1032, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Wróbel-Biedrawa, D.; Grabowska, K.; Galanty, A.; Sobolewska, D.; Podolak, I. A Flavonoid on the Brain: Quercetin as a Potential Therapeutic Agent in Central Nervous System Disorders. Life 2022, 12, 591. [Google Scholar] [CrossRef] [PubMed]
- Dookun, E.; Passos, J.F.; Arthur, H.M.; Richardson, G.D. Therapeutic Potential of Senolytics in Cardiovascular Disease. Cardiovasc. Drugs Ther. 2022, 36, 187–196. [Google Scholar] [CrossRef]
- Barile, E.; De, S.K.; Pellecchia, M. PDK1 inhibitors. Pharm. Pat. Anal. 2012, 1, 145–163. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IL-7 Signaling | PTSD/CVD | References (Adapted from) |
| Upregulate Warburg effect | Upregulate Warburg effect | [153] |
| Lower SARS-CoV-2 risk | Higher SARS-CoV-2 risk | [154,155,156] |
| Thymic activation | Thymic autophagy | [65,157] |
| Upregulated autophagy | Downregulated autophagy | [158,159,160,161] |
| Lower endothelial/immune senescence | Increased endothelial/immune senescence | [44,162] |
| Therapeutic Agent | Mechanism | References (Adapted from) |
| OXT | ||
| Intranasal OXT | Substitution therapy | |
| Lactobacillus reuteri | Synthesizes OXT | [196,197] |
| HIF-1α | ||
| Intermittent hypoxia | Upregulates HIF-1α | [8,9,10] |
| Hyperbaric oxygen | Lowers hypoxia | [11,12,13,14,15,16] |
| A2B blockers | ||
| PSB-603 | Decrease obesity and aging | [203,204] |
| ASIC blockers | ||
| Amyloride | Potassium-sparing diuretic | [214,215] |
| C5B | Neuroprotective | [216] |
| Sevanol | ASIC1 and ASIC3 inhibitor | [219,220] |
| Epigallocatechin gallate | ASIC3 inhibitor | |
| LDH inhibitors | ||
| 5-ALA | Improves mood, fatigue, sleep | [229,230,231] |
| Quinoline 3-sulfonamides | Reverses glycolysis | [233,234] |
| Galloflavin | Inhibits lactate production; decreases ATP synthesis | [217] |
| Autophagy activators | ||
| Antidepressants/ketamine | Increase autophagic markers LC3II/I | [242,243,244,245] |
| Recombinant ERAP2 | Lower ER stress | [246] |
| Lactylation inhibitors | ||
| Demethylzeylasteral (DML) | Lowers depression, atherosclerosis, pain | [238] |
| IL-7 | ||
| IL-7 recombinant GX-17 | Prevents cellular senescence | [247,248] |
| Thymic activators | ||
| Photobiomodulation | Restores thymic function | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozlakidis, Z.; Shi, P.; Abarbanel, G.; Klein, C.; Sfera, A. Recent Developments in Protein Lactylation in PTSD and CVD: Novel Strategies and Targets. BioTech 2023, 12, 38. https://doi.org/10.3390/biotech12020038
Kozlakidis Z, Shi P, Abarbanel G, Klein C, Sfera A. Recent Developments in Protein Lactylation in PTSD and CVD: Novel Strategies and Targets. BioTech. 2023; 12(2):38. https://doi.org/10.3390/biotech12020038
Chicago/Turabian StyleKozlakidis, Zisis, Patricia Shi, Ganna Abarbanel, Carolina Klein, and Adonis Sfera. 2023. "Recent Developments in Protein Lactylation in PTSD and CVD: Novel Strategies and Targets" BioTech 12, no. 2: 38. https://doi.org/10.3390/biotech12020038
APA StyleKozlakidis, Z., Shi, P., Abarbanel, G., Klein, C., & Sfera, A. (2023). Recent Developments in Protein Lactylation in PTSD and CVD: Novel Strategies and Targets. BioTech, 12(2), 38. https://doi.org/10.3390/biotech12020038