


Styrene Production in Genetically Engineered Escherichia coli in a Two-Phase Culture

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Plasmid Construction
2.2. Culture Conditions
2.3. Analytical Methods
2.4. Analytical Methods for Compounds
3. Results and Discussion
3.1. Screening for the Optimized Gene Encoding PAL
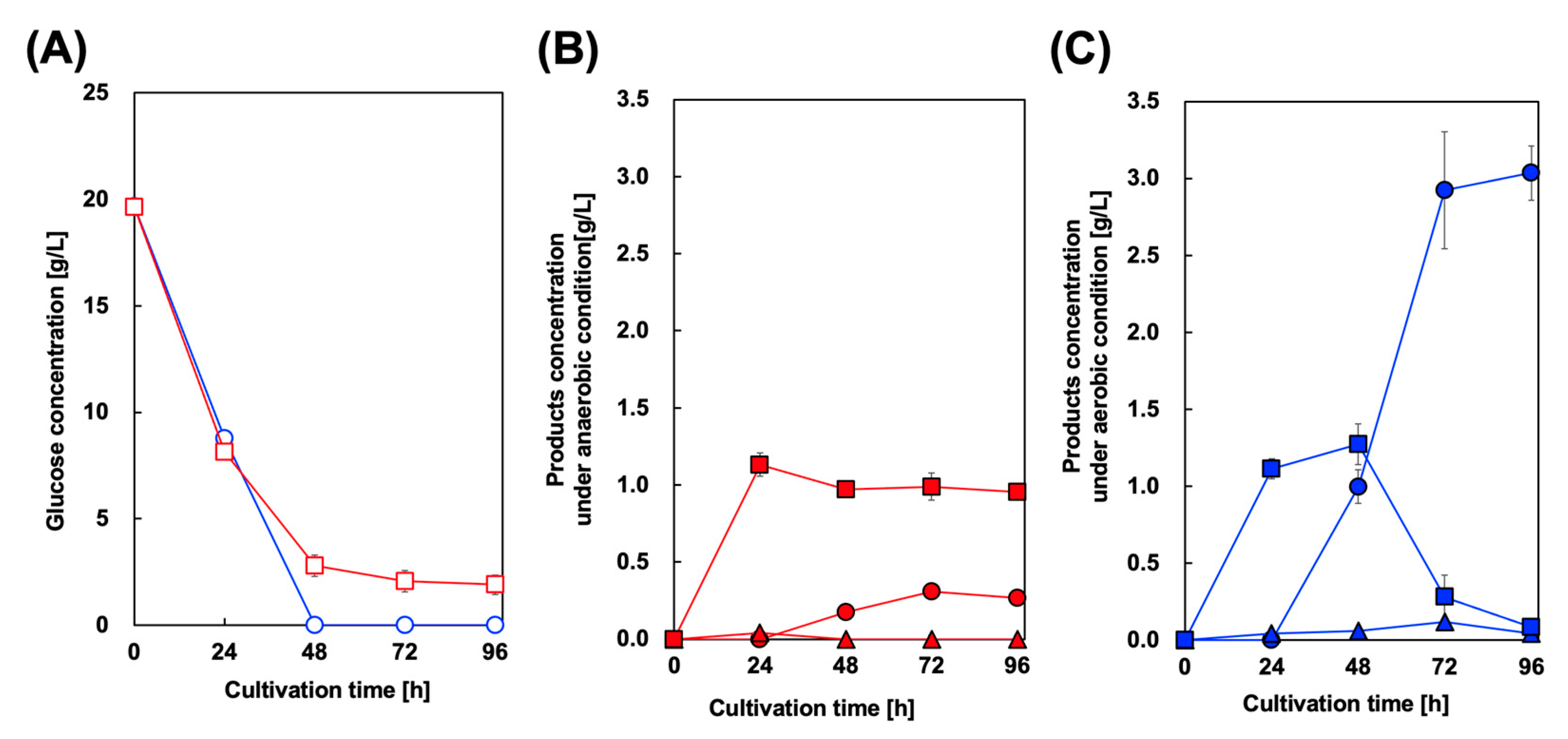
3.2. Styrene Production Using Single-Phase Batch Culture with E. coli Co-Expressing PAL and FDC
3.3. Two-Phase Batch Cultivation of Styrene-Producing E. coli Using OA
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Cruz-Morales, P.; Zargar, A.; Belcher, M.S.; Pang, B.; Englund, E.; Dan, Q.; Yin, K.; Keasling, J.D. Biofuels for a Sustainable Future. Cell 2021, 184, 1636–1647. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, X.; Wang, J.; Yuan, X.; Jiang, X.; Wang, Y.; Zhong, C.; Xu, D.; Gu, T.; Wang, F. Bacterial Biofilms as Platforms Engineered for Diverse Applications. Biotechnol. Adv. 2022, 57, 107932. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Du, N.; Shen, R.; He, T.; Xi, J.; Tan, J.; Bian, G.; Yang, Y.; Liu, T.; Tan, W.; et al. Redox Signaling-Driven Modulation of Microbial Biosynthesis and Biocatalysis. Nat. Commun. 2023, 14, 6800–6812. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, L.; Liu, P.; Yuan, Q.; Ma, C.; Wang, W.; Zhang, C.; Ma, H.; Zeng, A. Design, Evaluation, and Implementation of Synthetic Isopentyldiol Pathways in Escherichia coli. ACS Synth. Biol. 2023, 12, 3381–3392. [Google Scholar] [CrossRef]
- Madhavan, A.; Arun, K.B.; Sindhu, R.; Nair, B.G.; Pandey, A.; Awasthi, M.K.; Szakacs, G.; Binod, P. Design and Genome Engineering of Microbial Cell Factories for Efficient Conversion of Lignocellulose to Fuel. Bioresour. Technol. 2023, 370, 128555. [Google Scholar] [CrossRef]
- Xie, Y.; Erşan, S.; Guan, X.; Wang, J.; Sha, J.; Xu, S.; Wohlschlegel, J.A.; Park, J.O.; Liu, C. Unexpected Metabolic Rewiring of CO2 Fixation in H2-Mediated Materials-Biology Hybrids. Proc. Natl. Acad. Sci. USA 2023, 120, e2308373120. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, J.; Zhang, Y.; Chen, L.; Ouyang, Z.; Liao, D.; Zhao, F.; Han, S. Characterization of the Enzyme Kinetics of EMP and HMP Pathway in Corynebacterium glutamicum: Reference for Modeling Metabolic Networks. Front. Bioeng. Biotechnol. 2023, 11, 1296880. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, J.; Gao, P.; Yan, W.; Shen, J.; Luo, X.; Keasling, J.D. Development of an Efficient Yeast Platform for Cannabigerolic Acid Biosynthesis. Metab. Eng. 2023, 80, 232–240. [Google Scholar] [CrossRef]
- Sharan, S.K.; Thomason, L.C.; Kuznetsov, S.G.; Court, D.L. Recombineering: A Homologous Recombination-Based Method of Genetic Engineering. Nat. Protoc. 2009, 4, 206–223. [Google Scholar] [CrossRef]
- Jeong, J.; Cho, N.; Jung, D.; Bang, D. Genome-Scale Genetic Engineering in Escherichia coli. Biotechnol. Adv. 2013, 31, 804–810. [Google Scholar] [CrossRef]
- Yan, M.Y.; Yan, H.Q.; Ren, G.X.; Zhao, J.P.; Guo, X.P.; Sun, Y.C. CRISPR-Cas12a-Assisted Recombineering in Bacteria. Appl. Environ. Microbiol. 2017, 83, e00947-17. [Google Scholar] [CrossRef]
- Pontrelli, S.; Chiu, T.Y.; Lan, E.I.; Chen, F.Y.; Chang, P.; Liao, J.C. Escherichia coli as a Host for Metabolic Engineering. Metab. Eng. 2018, 50, 16–46. [Google Scholar] [CrossRef]
- Tong, Y.; Jørgensen, T.S.; Whitford, C.M.; Weber, T.; Lee, S.Y. A Versatile Genetic Engineering Toolkit for E. coli Based on CRISPR-Prime Editing. Nat. Commun. 2021, 12, 5206–5216. [Google Scholar] [CrossRef]
- Cen, X.; Dong, Y.; Liu, D.; Chen, Z. New Pathways and Metabolic Engineering Strategies for Microbial Synthesis of Diols. Curr. Opin. Biotechnol. 2022, 78, 102845. [Google Scholar] [CrossRef]
- Baez, A.; Cho, K.M.; Liao, J.C. High-Flux Isobutanol Production Using Engineered Escherichia coli: A Bioreactor with in Situ Product Removal. Appl. Microbiol. Biotechnol. 2011, 90, 1681–1690. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, G.M.; Atsumi, S. Isobutyraldehyde Production from Escherichia coli by Removing Aldehyde Reductase Activity. Microb. Cell. Fact. 2012, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.W.; Cho, J.S.; Lee, S.Y. Microbial Production of Methyl Anthranilate, a Grape Flavor Compound. Proc. Natl. Acad. Sci. USA 2019, 116, 10749–10756. [Google Scholar] [CrossRef] [PubMed]
- Combes, J.; Imatoukene, N.; Couvreur, J.; Godon, B.; Brunissen, F.; Fojcik, C.; Allais, F.; Lopez, M. Intensification of p-Coumaric Acid Heterologous Production Using Extractive Biphasic Fermentation. Bioresour. Technol. 2021, 337, 125436. [Google Scholar] [CrossRef]
- Luo, Z.W.; Lee, S.Y. Biotransformation of p-Xylene to Terephthalic Acid Using Engineered Escherichia coli. Nat. Commun. 2017, 8, 15689. [Google Scholar] [CrossRef]
- Mori, Y.; Noda, S.; Shirai, T.; Kondo, A. Direct 1,3-Butadiene Biosynthesis in Escherichia coli via a Tailored Ferulic Acid Decarboxylase Mutant. Nat. Commun. 2021, 12, 2195. [Google Scholar] [CrossRef]
- Acquavella, J.F.; Leonard, R.C. Review of the Epidemiology of 1,3-Butadiene and Chloroprene. Chem. Biol. Interact. 2001, 135–136, 43–52. [Google Scholar] [CrossRef]
- McKenna, R.; Nielsen, D.R. Styrene Biosynthesis from Glucose by Engineered E. coli. Metab. Eng. 2011, 13, 544–554. [Google Scholar] [CrossRef] [PubMed]
- McKenna, R.; Thompson, B.; Pugh, S.; Nielsen, D.R. Rational and Combinatorial Approaches to Engineering Styrene Production by Saccharomyces cerevisiae. Microb. Cell. Fact. 2014, 13, 123. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, R.; Noda, S.; Tanaka, T.; Kondo, A. Styrene Production from a Biomass-Derived Carbon Source Using a Coculture System of Phenylalanine Ammonia Lyase and Phenylacrylic Acid Decarboxylase-Expressing Streptomyces lividans Transformants. J. Biosci. Bioeng. 2016, 122, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; McKenna, R.; Rover, M.R.; Wen, Z.; Jarboe, L.R. Production of Biorenewable Styrene: Utilization of Biomass-Derived Sugars and Insights into Toxicity. J. Ind. Microbiol. Biotechnol. 2016, 43, 595–604. [Google Scholar] [CrossRef]
- Liu, C.; Men, X.; Chen, H.; Li, M.; Ding, Z.; Chen, G.; Wang, F.; Liu, H.; Wang, Q.; Zhu, Y.; et al. Systematic Optimization of Styrene Biosynthesis in Escherichia coli BL21(DE3). Biotechnol. Biofuels 2018, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Bang, H.B.; Lee, Y.H.; Jeong, K.J. Enhanced Styrene Production by Engineered Escherichia coli and in Situ Product Recovery (ISPR) with an Organic Solvent. Microb. Cell. Fact. 2019, 18, 79. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Liu, R.; Foster, K.E.O.; Choudhury, A.; Cook, S.; Cameron, J.C.; Srubar, W.V., III; Gill, R.T. Genome Engineering of E. coli for Improved Styrene Production. Metab. Eng. 2020, 57, 74–84. [Google Scholar] [CrossRef]
- Williamson, J.J.; Bahrin, N.; Hardiman, E.M.; Bugg, T.D.H. Production of Substituted Styrene Bioproducts from Lignin and Lignocellulose Using Engineered Pseudomonas putida KT2440. Biotechnol. J. 2020, 15, e1900571. [Google Scholar] [CrossRef]
- Webb, J.P.; Paiva, A.C.; Rossoni, L.; Alstrom-Moore, A.; Springthorpe, V.; Vaud, S.; Yeh, V.; Minde, D.P.; Langer, S.; Walker, H.; et al. Multi-Omic Based Production Strain Improvement (MOBpsi) for Bio-Manufacturing of Toxic Chemicals. Metab. Eng. 2022, 72, 133–149. [Google Scholar] [CrossRef]
- Noda, S.; Shirai, T.; Oyama, S.; Kondo, A. Metabolic Design of a Platform Escherichia coli Strain Producing Various Chorismate Derivatives. Metab. Eng. 2016, 33, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Noda, S.; Shirai, T.; Mori, Y.; Oyama, S.; Kondo, A. Engineering a Synthetic Pathway for Maleate in Escherichia coli. Nat. Commun. 2017, 8, 1153. [Google Scholar] [CrossRef] [PubMed]
- Noda, S.; Mori, Y.; Fujiwara, R.; Shirai, T.; Tanaka, T.; Kondo, A. Reprogramming the Escherichia coli Pyruvate-Forming Reaction Towards Chorismate Derivative Production. Metab. Eng. 2021, 67, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Noda, S.; Miyazaki, T.; Miyoshi, T.; Miyake, M.; Okai, N.; Tanaka, T.; Ogino, C.; Kondo, A. Cinnamic Acid Production Using Streptomyces lividans Expressing Phenylalanine Ammonia-Lyase. J. Ind. Microbiol. Biotechnol. 2011, 38, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Gottardi, M.; Knudsen, J.D.; Prado, L.; Oreb, M.; Branduardi, P.; Boles, E. De novo Biosynthesis of Trans-Cinnamic Acid Derivatives in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2017, 101, 4883–4893. [Google Scholar] [CrossRef]
- Kukil, K.; Lindberg, P. Expression of Phenylalanine Ammonia-Lyases in Synechocystis sp. PCC 6803 and Subsequent Improvements in the Sustainable Production of Phenylpropanoids. Microb. Cell. Fact. 2022, 21, 8. [Google Scholar] [CrossRef]
- Pant, S.R.; Irigoyen, S.; Liu, J.; Bedre, R.; Christensen, S.A.; Schmelz, E.A.; Sedbrook, J.C.; Scholthof, K.B.G.; Mandadi, K.K. Brachypodium Phenylalanine Ammonia Lyase (PAL) Promotes Antiviral Defense Against the Panicum Mosaic Virus and Its Satellites. mBio 2021, 12, e03518-20. [Google Scholar] [CrossRef]
- Bang, H.B.; Lee, K.; Lee, Y.J.; Jeong, K.J. High-Level Production of trans-Cinnamic Acid by Fed-Batch Cultivation of Escherichia coli. Process. Biochem. 2018, 68, 30–36. [Google Scholar] [CrossRef]
- Anvari, M.; Pahlavanzadeh, H.; Vasheghani-Farahani, E.; Khayati, G. In Situ Recovery of 2,3-Butanediol from Fermentation by Liquid-Liquid Extraction. J. Ind. Microbiol. Biotechnol. 2009, 36, 313–317. [Google Scholar] [CrossRef]
- Ku, J.T.; Simanjuntak, W.; Lan, E.I. Renewable Synthesis of n-Butyraldehyde from Glucose Using Engineered Escherichia coli. Biotechnol. Biofuels 2017, 10, 291. [Google Scholar] [CrossRef]
- Contreras-Dávila, C.A.; Zuidema, N.; Buisman, C.J.N.; Strik, D.P.B.T.B. The Reactor Microbiome Enriches Vegetable Oil with n-Caproate and n-Caprylate for Potential Functionalized Feed Additive Production via Extractive Lactate-Based Chain Elongation. Biotechnol. Biofuels 2021, 14, 232. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.J.; Gong, G.; Ahn, J.H.; Ko, J.K.; Lee, S.M.; Um, Y. Effective Hexanol Production from Carbon Monoxide Using Extractive Fermentation with Clostridium carboxidivorans P7. Bioresour. Technol. 2023, 367, 128201. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Genotype | Source or Reference |
|---|---|---|
| NovaBlue | endA1 hsdR17(rK12−mK12+) supE44 thi-I gyrA96 relA1 lac recA1/F’ | Novagen |
| [proAB+ lacIq ZΔM15::Tn10(Tetr)]; used for gene cloning | ||
| ATCC 31882 (defined as CFT0) | L-phenylalanine-overproducing strain | American Type Culture Collection |
| CFT3 | ATCC31882 ptsHI::PA1lacO-1-Glk-GalPΔpykFΔpykA | [31] |
| CFT3A2 | CFT3 harboring pZE12-AtPAL2 | This study |
| CFT3B1 | CFT3 harboring pZE12-BdPAL1 | This study |
| CFT3B2 | CFT3 harboring pZE12-BdPAL2 | This study |
| CFT3B6 | CFT3 harboring pZE12-BdPAL6 | This study |
| CFT3B8 | CFT3 harboring pZE12-BdPAL8 | This study |
| CFT3A2FD | CFT3A2 harboring pSAK-FDC1 | This study |
| Plasmids | ||
| pZE12-Ptrc | Ptrc, colE1 ori, Ampr | [33] |
| pSAK | PAlacO-1, SC101 ori, Cmr | [32] |
| pSAK-Ptrc | Ptrc, SC101 ori, Cmr | This study |
| pZE12-AtPAL2 | pZE12-Ptrc containing AtPAL2 from A. thaliana | This study |
| pZE12-BdPAL1 | pZE12-Ptrc containing BdPAL1 from B. distachyon | This study |
| pZE12-BdPAL2 | pZE12-Ptrc containing BdPAL2 from B. distachyon | This study |
| pZE12-BdPAL6 | pZE12-Ptrc containing BdPAL6 from B. distachyon | This study |
| pZE12-BdPAL8 | pZE12-Ptrc containing BdPAL8 from B. distachyon | This study |
| pSAK-FDC1 | pSAK-Ptrc containing FDC1 from S. cerevisiae | This study |
| Host | Genotype | Pmax (g L−1) | Yield (mol mol−1) | Substrate | Reference |
|---|---|---|---|---|---|
| E. coli CFT3 | aroG39, aroF394, PheA101, pheO352, tyrR366 tyrA4, trpE401, lacY5, malT384, thi-1 ptsHI::PA1lacO-1-glk-galPΔpykFΔpykA | 3.01 | 0.27 | Glucose | This study |
| E. coli NST74 | aroH367, tyrR366, tna-2, lacY5, aroF394(fbr), malT384, pheA101(fbr), pheO352, aroG397(fbr) | 0.26 | N.E. | Glucose | [22] |
| S. cerevisiae 22A75D | MATa his3Δ0 leu2Δ0 met15Δ0 ura3Δ0 aro10Δ::aro4K229L | 0.029 | 0.0025 | Glucose | [23] |
| S. lividans 1326 (coculture) | Wild-type strain | 0.029 | N.E. | Glucose | [24] |
| E. coli NST74 | aroH367, tyrR366, tna-2, lacY5, aroF394(fbr), malT384, pheA101(fbr), pheO352, aroG397(fbr) | 0.251 | N.E. | Pyrolytic sugars | [25] |
| E. coli BL21(DE3) | dcm ompT hsdS(rB− mB−) gal | 0.35 | N.E. | Glucose | [26] |
| E. coli YHP05 | F− l− rph-1 INV(rrnD, rrnE)Δcrr ΔtyrR ΔtrpE ΔtyrA ΔpykA | 1.7 | N.E. | Glucose | [27] |
| E. coli NST74 | aroH367, tyrR366, tna-2, lacY5, aroF394(fbr), malT384, pheA101(fbr), pheO352, aroG397(fbr) | 0.345 | N.E. | Glucose | [28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noda, S.; Fujiwara, R.; Mori, Y.; Dainin, M.; Shirai, T.; Kondo, A. Styrene Production in Genetically Engineered Escherichia coli in a Two-Phase Culture. BioTech 2024, 13, 2. https://doi.org/10.3390/biotech13010002
Noda S, Fujiwara R, Mori Y, Dainin M, Shirai T, Kondo A. Styrene Production in Genetically Engineered Escherichia coli in a Two-Phase Culture. BioTech. 2024; 13(1):2. https://doi.org/10.3390/biotech13010002
Chicago/Turabian StyleNoda, Shuhei, Ryosuke Fujiwara, Yutaro Mori, Mayumi Dainin, Tomokazu Shirai, and Akihiko Kondo. 2024. "Styrene Production in Genetically Engineered Escherichia coli in a Two-Phase Culture" BioTech 13, no. 1: 2. https://doi.org/10.3390/biotech13010002
APA StyleNoda, S., Fujiwara, R., Mori, Y., Dainin, M., Shirai, T., & Kondo, A. (2024). Styrene Production in Genetically Engineered Escherichia coli in a Two-Phase Culture. BioTech, 13(1), 2. https://doi.org/10.3390/biotech13010002