
An Unconventional Ligand for Scribble PDZ-4 Domain Mediates Its Interaction with Dusp26


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Constructs and Plasmids
2.2. Site-Specific Mutagenesis
2.3. Recombinant Protein Production and Purification
2.4. Pull-Down Assay
2.5. Affinity Chromatography
2.6. ELISA Assay
2.7. Molecular Docking Analysis
3. Results
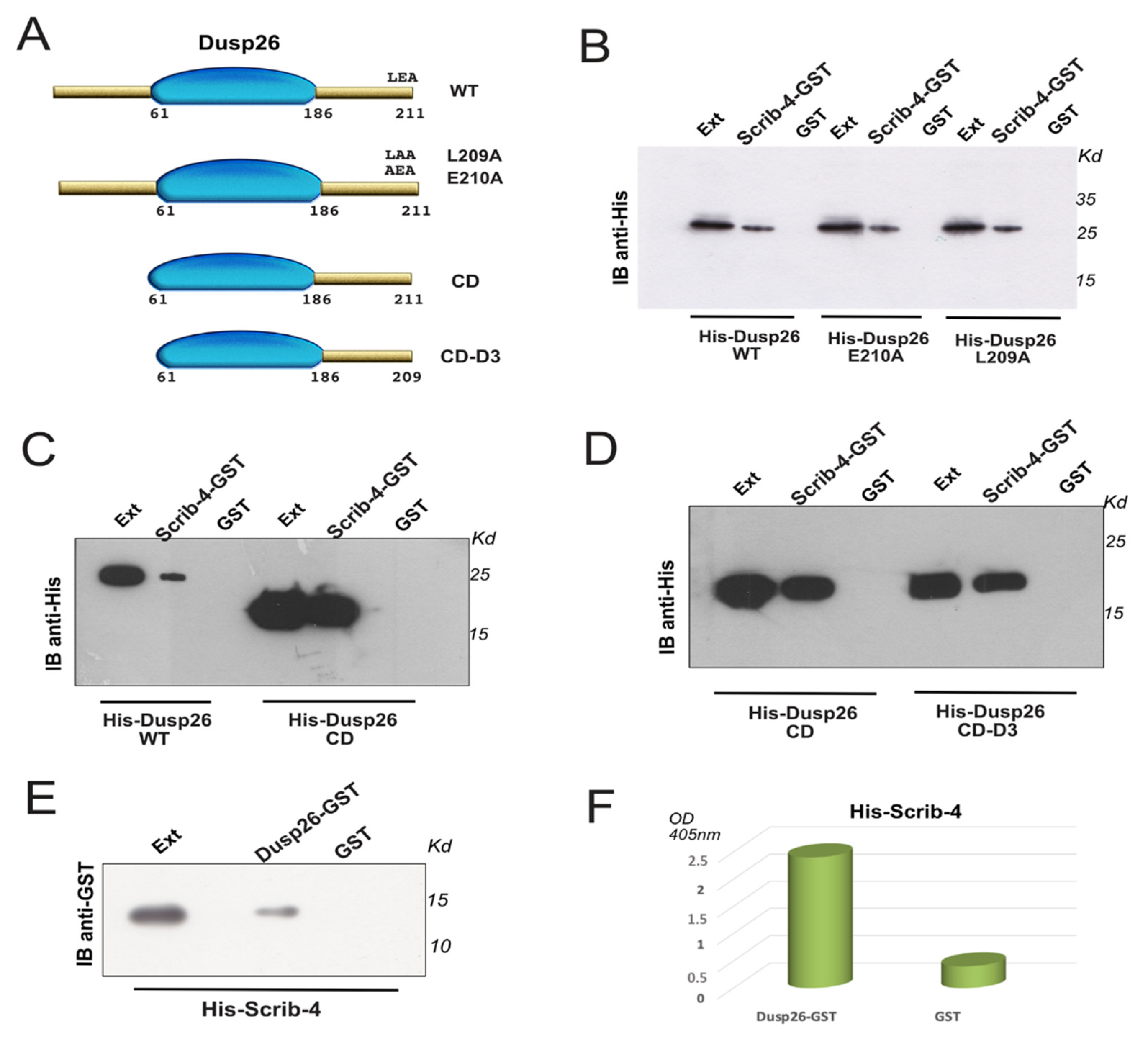
3.1. Scribble PDZ-4 Binds to an Internal Site of Dusp26
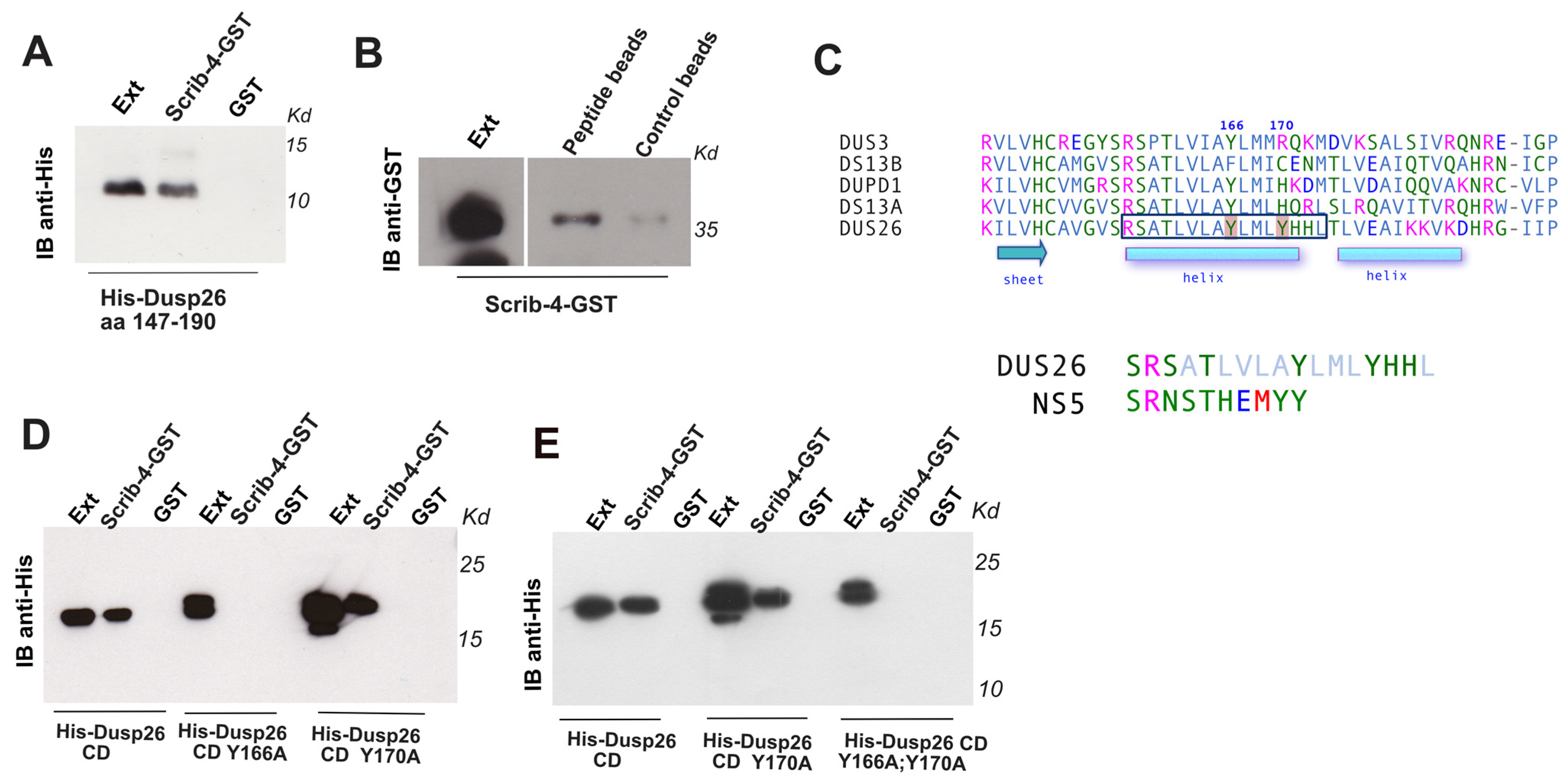
3.2. Mapping the Internal Region Binding Site
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pawson, T.; Raina, M.; Nash, P. Interaction domains: From simple binding events to complex cellular behavior. FEBS Lett. 2001, 513, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Zucconi, A.; Panni, S.; Paoluzi, S.; Castagnoli, L.; Dente, L.; Cesareni, G. Domain repertoires as a tool to derive protein recognition rules. FEBS Lett. 2000, 480, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Harris, B.Z.; Lim, W.A. Mechanism and role of PDZ domains in signaling complex assembly. J. Cell Sci. 2001, 114, 3219–3231. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Fuentes, E.J. Emerging themes in PDZ domain signaling: Structure, function, and inhibition. Int. Rev. Cell Mol. Biol. 2019, 343, 129–218. [Google Scholar]
- Christensen, N.R.; Čalyševa, J.; Fernandes, E.F.A.; Lüchow, S.; Clemmensen, L.S.; Haugaard-Kedström, L.M.; Strømgaard, K. PDZ Domains as Drug Targets. Adv. Ther. 2019, 2, 1800143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, D.A.; Lee, A.; Lewis, J.; Kim, E.; Sheng, M.; MacKinnon, R. Crystal Structures of a Complexed and Peptide-Free Membrane Protein–Binding Domain: Molecular Basis of Peptide Recognition by PDZ. Cell 1996, 85, 1067–1076. [Google Scholar] [CrossRef] [Green Version]
- Luck, K.; Charbonnier, S.; Travé, G. The emerging contribution of sequence context to the specificity of protein interactions mediated by PDZ domains. FEBS Lett. 2012, 586, 2648–2661. [Google Scholar] [CrossRef] [Green Version]
- Ivarsson, Y.; Arnold, R.; McLaughlin, M.; Nim, S.; Joshi, R.; Ray, D.; Liu, B.; Teyra, J.; Pawson, T.; Moffat, J.; et al. Large-scale interaction profiling of PDZ domains through proteomic peptide-phage display using human and viral phage peptidomes. Proc. Natl. Acad. Sci. USA 2014, 111, 2542–2547. [Google Scholar] [CrossRef] [Green Version]
- Songyang, Z.; Fanning, A.S.; Fu, C.; Xu, J.; Marfatia, S.M.; Chishti, A.H.; Crompton, A.; Chan, A.C.; Anderson, J.M.; Cantley, L.C. Recognition of Unique Carboxyl-Terminal Motifs by Distinct PDZ Domains. Science 1997, 275, 73–77. [Google Scholar] [CrossRef]
- Stricker, N.L.; Christopherson, K.S.; Yi, B.A.; Schatz, P.J.; Raab, R.W.; Dawes, G.; Bassett, D.E.; Bredt, D.S.; Li, M. PDZ domain of neuronal nitric oxide synthase recognizes novel C-terminal peptide sequences. Nat. Biotechnol. 1997, 15, 336–342. [Google Scholar] [CrossRef]
- Tonikian, R.; Zhang, Y.; Sazinsky, S.L.; Currell, B.; Yeh, J.-H.; Reva, B.; Held, H.A.; Appleton, B.A.; Evangelista, M.; Wu, Y.; et al. A Specificity Map for the PDZ Domain Family. PLOS Biol. 2008, 6, e239. [Google Scholar] [CrossRef] [Green Version]
- Hillier, B.J.; Christopherson, K.S.; Prehoda, K.E.; Bredt, D.S.; Lim, W.A. Unexpected modes of PDZ domain scaffolding revealed by structure of nNOS-syntrophin complex. Science 1999, 284, 812–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenfant, N.; Polanowska, J.; Bamps, S.; Omi, S.; Borg, J.-P.; Reboul, J. A genome-wide study of PDZ-domain interactions in C. elegans reveals a high frequency of non-canonical binding. BMC Genom. 2010, 11, 671. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; McAuley, M.M.; Lüchow, S.; Knapp, S.; Joerger, A.C.; Ivarsson, Y. Integrated analysis of Shank1 PDZ interactions with C-terminal and internal binding motifs. Curr. Res. Struct. Biol. 2021, 3, 41–50. [Google Scholar] [CrossRef]
- Zhang, Y.; Appleton, B.A.; Wiesmann, C.; Lau, T.; Costa, M.; Hannoush, R.N.; Sidhu, S.S. Inhibition of Wnt signaling by Dishevelled PDZ peptides. Nat. Chem. Biol. 2009, 5, 217–219. [Google Scholar] [CrossRef] [PubMed]
- Penkert, R.R.; DiVittorio, H.M.; Prehoda, K.E. Internal recognition through PDZ domain plasticity in the Par-6–Pals1 complex. Nat. Struct. Mol. Biol. 2004, 11, 1122–1127. [Google Scholar] [CrossRef] [PubMed]
- Sacco, F.; Boldt, K.; Calderone, A.; Panni, S.; Paoluzi, S.; Castagnoli, L.; Ueffing, M.; Cesareni, G. Combining affinity proteomics and network context to identify new phosphatase substrates and adapters in growth pathways. Front. Genet. 2014, 5, 115. [Google Scholar] [CrossRef] [Green Version]
- Luck, K.; Fournane, S.; Kieffer, B.; Masson, M.; Nominé, Y.; Travé, G. Putting into Practice Domain-Linear Motif Interaction Predictions for Exploration of Protein Networks. PLoS ONE 2011, 6, e25376. [Google Scholar] [CrossRef]
- Lim, K.; Godde, N.; Humbert, P.O.; Kvansakul, M. Structural basis for the differential interaction of Scribble PDZ domains with the guanine nucleotide exchange factor β-PIX. J. Biol. Chem. 2017, 292, 20425–20436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takizawa, S.; Nagasaka, K.; Nakagawa, S.; Yano, T.; Nakagawa, K.; Yasugi, T.; Takeuchi, T.; Kanda, T.; Huibregtse, J.M.; Akiyama, T.; et al. Human scribble, a novel tumor suppressor identified as a target of high-risk HPV E6 for ubiquitin-mediated degradation, interacts with adenomatous polyposis coli. Genes Cells 2006, 11, 453–464. [Google Scholar] [CrossRef]
- Janezic, E.M.; Harris, D.-A.; Dinh, D.; Lee, K.-S.; Stewart, A.; Hinds, T.R.; Hsu, P.L.; Zheng, N.; Hague, C. Scribble co-operatively binds multiple α1D-adrenergic receptor C-terminal PDZ ligands. Sci. Rep. 2019, 9, 14073. [Google Scholar] [CrossRef] [Green Version]
- Werme, K.; Wigerius, M.; Johansson, M. Tick-borne encephalitis virus NS5 associates with membrane protein scribble and impairs interferon-stimulated JAK-STAT signalling. Cell. Microbiol. 2008, 10, 696–712. [Google Scholar] [CrossRef]
- Richier, L.; Williton, K.; Clattenburg, L.; Colwill, K.; O’Brien, M.; Tsang, C.; Kolar, A.; Zinck, N.; Metalnikov, P.; Trimble, W.S.; et al. NOS1AP Associates with Scribble and Regulates Dendritic Spine Development. J. Neurosci. 2010, 30, 4796–4805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melik, W.; Ellencrona, K.; Wigerius, M.; Hedström, C.; Elväng, A.; Johansson, M. Two PDZ binding motifs within NS5 have roles in Tick-borne encephalitis virus replication. Virus Res. 2012, 169, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Zundert, G.; Rodrigues, J.; Trellet, M.; Schmitz, C.; Kastritis, P.; Karaca, E.; Melquiond, A.; van Dijk, M.; de Vries, S.; Bonvin, A. The HADDOCK2.2 Web Server: User-Friendly Integrative Modeling of Biomolecular Complexes. J. Mol. Biol. 2015, 428, 720–725. [Google Scholar] [CrossRef] [Green Version]
- De Vries, S.J.; Bonvin, A.M.J.J. CPORT: A Consensus Interface Predictor and Its Performance in Prediction-Driven Docking with HADDOCK. PLoS ONE 2011, 6, e17695. [Google Scholar] [CrossRef] [Green Version]
- Méndez, R.; Leplae, R.; De Maria, L.; Wodak, S. Assessment of blind predictions of protein-protein interactions: Current status of docking methods. Proteins: Struct. Funct. Bioinform. 2003, 52, 51–67. [Google Scholar] [CrossRef]
- Xue, L.C.; Rodrigues, J.P.; Kastritis, P.L.; Bonvin, A.M.; Vangone, A. PRODIGY: A web server for predicting the binding affinity of protein–protein complexes. Bioinformatics 2016, 32, 3676–3678. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Lokareddy, R.K.; Bhardwaj, A.; Cingolani, G. Atomic Structure of Dual-Specificity Phosphatase 26, a Novel p53 Phosphatase. Biochemistry 2013, 52, 938–948. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E.; Stoker, A. A Review of DUSP26: Structure, Regulation and Relevance in Human Disease. Int. J. Mol. Sci. 2021, 22, 776. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Mivechi, N.F. Association and Regulation of Heat Shock Transcription Factor 4b with both Extracellular Signal-Regulated Kinase Mitogen-Activated Protein Kinase and Dual-Specificity Tyrosine Phosphatase DUSP26. Mol. Cell. Biol. 2006, 26, 3282–3294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanuma, N.; Nomura, M.; Ikeda, M.; Kasugai, I.; Tsubaki, Y.; Takagaki, K.; Kawamura, T.; Yamashita, Y.; Sato, I.; Sato, M.; et al. Protein phosphatase Dusp26 associates with KIF3 motor and promotes N-cadherin-mediated cell–cell adhesion. Oncogene 2008, 28, 752–761. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Lee, H.-J.; Oh, Y.; Choi, S.-G.; Hong, S.-H.; Kim, H.-J.; Lee, S.-Y.; Choi, J.-W.; Hwang, D.S.; Kim, K.-S.; et al. The DUSP26 phosphatase activator adenylate kinase 2 regulates FADD phosphorylation and cell growth. Nat. Commun. 2014, 5, 3351. [Google Scholar] [CrossRef]
- Sacco, F.; Gherardini, P.; Paoluzi, S.; Saez-Rodriguez, J.; Helmer-Citterich, M.; Ragnini, A.; Castagnoli, L.; Cesareni, G. Mapping the human phosphatome on growth pathways. Mol. Syst. Biol. 2012, 8, 603. [Google Scholar] [CrossRef]
- Cuppen, E.; Gerrits, H.; Pepers, B.; Wieringa, B.; Hendriks, W. PDZ Motifs in PTP-BL and RIL Bind to Internal Protein Segments in the LIM Domain Protein RIL. Mol. Biol. Cell 1998, 9, 671–683. [Google Scholar] [CrossRef] [Green Version]
- Kock, G.; Dicks, M.; Yip, K.T.; Kohl, B.; Pütz, S.; Heumann, R.; Erdmann, K.S.; Stoll, R. Molecular Basis of Class III Ligand Recognition by PDZ3 in Murine Protein Tyrosine Phosphatase PTPN13. J. Mol. Biol. 2018, 430, 4275–4292. [Google Scholar] [CrossRef]
- Chen, K.-E.; Lin, S.-Y.; Wu, M.-J.; Ho, M.-R.; Santhanam, A.; Chou, C.-C.; Meng, T.-C.; Wang, A.H.-J. Reciprocal allosteric regulation of p38γ and PTPN3 involves a PDZ domain–modulated complex formation. Sci. Signal. 2014, 7, ra98. [Google Scholar] [CrossRef]
- Iuliano, R.; Trapasso, F.; Samà, I.; Le Pera, I.; Martelli, M.L.; Lembo, F.; Santoro, M.; Viglietto, G.; Chiariotti, L.; Fusco, A. Rat protein tyrosine phosphatase η physically interacts with the PDZ domains of syntenin. FEBS Lett. 2001, 500, 41–44. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, Y.; Morales, F.C.; Kreimann, E.L.; Georgescu, M.-M. PTEN tumor suppressor associates with NHERF proteins to attenuate PDGF receptor signaling. EMBO J. 2006, 25, 910–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Yang, H.; Liu, J.; Schmidt, M.D.; Gao, T. Scribble-mediated membrane targeting of PHLPP1 is required for its negative regulation of Akt. EMBO Rep. 2011, 12, 818–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagasaka, K.; Seiki, T.; Yamashita, A.; Massimi, P.; Subbaiah, V.K.; Thomas, M.; Kranjec, C.; Kawana, K.; Nakagawa, S.; Yano, T.; et al. A Novel Interaction between hScrib and PP1γ Downregulates ERK Signaling and Suppresses Oncogene-Induced Cell Transformation. PLoS ONE 2013, 8, e53752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dow, L.E.; Elsum, I.A.; King, C.L.; Kinross, K.M.; Richardson, H.E.; Humbert, P.O. Loss of human Scribble cooperates with H-Ras to promote cell invasion through deregulation of MAPK signalling. Oncogene 2008, 27, 5988–6001. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Huang, C.; Wu, J.; Li, J.; Hu, T.; Wang, Z.; Zhang, H.; Shao, Y.; Fu, Z. SCRIB Promotes Proliferation and Metastasis by Targeting Hippo/YAP Signalling in Colorectal Cancer. Front. Cell Dev. Biol. 2021, 9, 656359. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.; Rahmouni, S.; Williams, S.; Van Stipdonk, M.; Jaroszewski, L.; Godzik, A.; Abraham, R.T.; Schoenberger, S.P.; Mustelin, T. Tyrosine phosphorylation of VHR phosphatase by ZAP-70. Nat. Immunol. 2003, 4, 44–48. [Google Scholar] [CrossRef]
- Hornbeck, P.V.; Zhang, B.; Murray, B.; Kornhauser, J.M.; Latham, V.; Skrzypek, E. PhosphoSitePlus, 2014: Mutations, PTMs and recalibrations. Nucleic Acids Res. 2014, 43, D512–D520. [Google Scholar] [CrossRef] [Green Version]
- Jeffrey, K.L.; Camps, M.; Rommel, C.; Mackay, C.R. Targeting dual-specificity phosphatases: Manipulating MAP kinase signalling and immune responses. Nat. Rev. Drug Discov. 2007, 6, 391–403. [Google Scholar] [CrossRef]
- Chen, J.; Zeng, Y.; Wu, R.; Xuan, Y.; Jiang, M.; Teng, H. Corrigendum: Decreased DUSP26 Expression Promotes Malignant Behavior in Glioblastoma Cells via Deregulation of MAPK and Akt Signaling Pathway. Front. Oncol. 2021, 11, 676647. [Google Scholar] [CrossRef]
- Wang, Y.; Han, D.; Zhou, T.; Chen, C.; Cao, H.; Zhang, J.Z.; Ma, N.; Liu, C.; Song, M.; Shi, J.; et al. DUSP26 induces aortic valve calcification by antagonizing MDM2-mediated ubiquitination of DPP4 in human valvular interstitial cells. Eur. Hear. J. 2021, 42, 2935–2951. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallo, R.; De Sensi, E.; Storino, F.; Panni, S. An Unconventional Ligand for Scribble PDZ-4 Domain Mediates Its Interaction with Dusp26. BioChem 2022, 2, 83-92. https://doi.org/10.3390/biochem2010006
Gallo R, De Sensi E, Storino F, Panni S. An Unconventional Ligand for Scribble PDZ-4 Domain Mediates Its Interaction with Dusp26. BioChem. 2022; 2(1):83-92. https://doi.org/10.3390/biochem2010006
Chicago/Turabian StyleGallo, Raffaella, Erika De Sensi, Francesca Storino, and Simona Panni. 2022. "An Unconventional Ligand for Scribble PDZ-4 Domain Mediates Its Interaction with Dusp26" BioChem 2, no. 1: 83-92. https://doi.org/10.3390/biochem2010006
APA StyleGallo, R., De Sensi, E., Storino, F., & Panni, S. (2022). An Unconventional Ligand for Scribble PDZ-4 Domain Mediates Its Interaction with Dusp26. BioChem, 2(1), 83-92. https://doi.org/10.3390/biochem2010006