

DMSO Alleviates LPS-Induced Inflammatory Responses in RAW264.7 Macrophages by Inhibiting NF-κB and MAPK Activation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
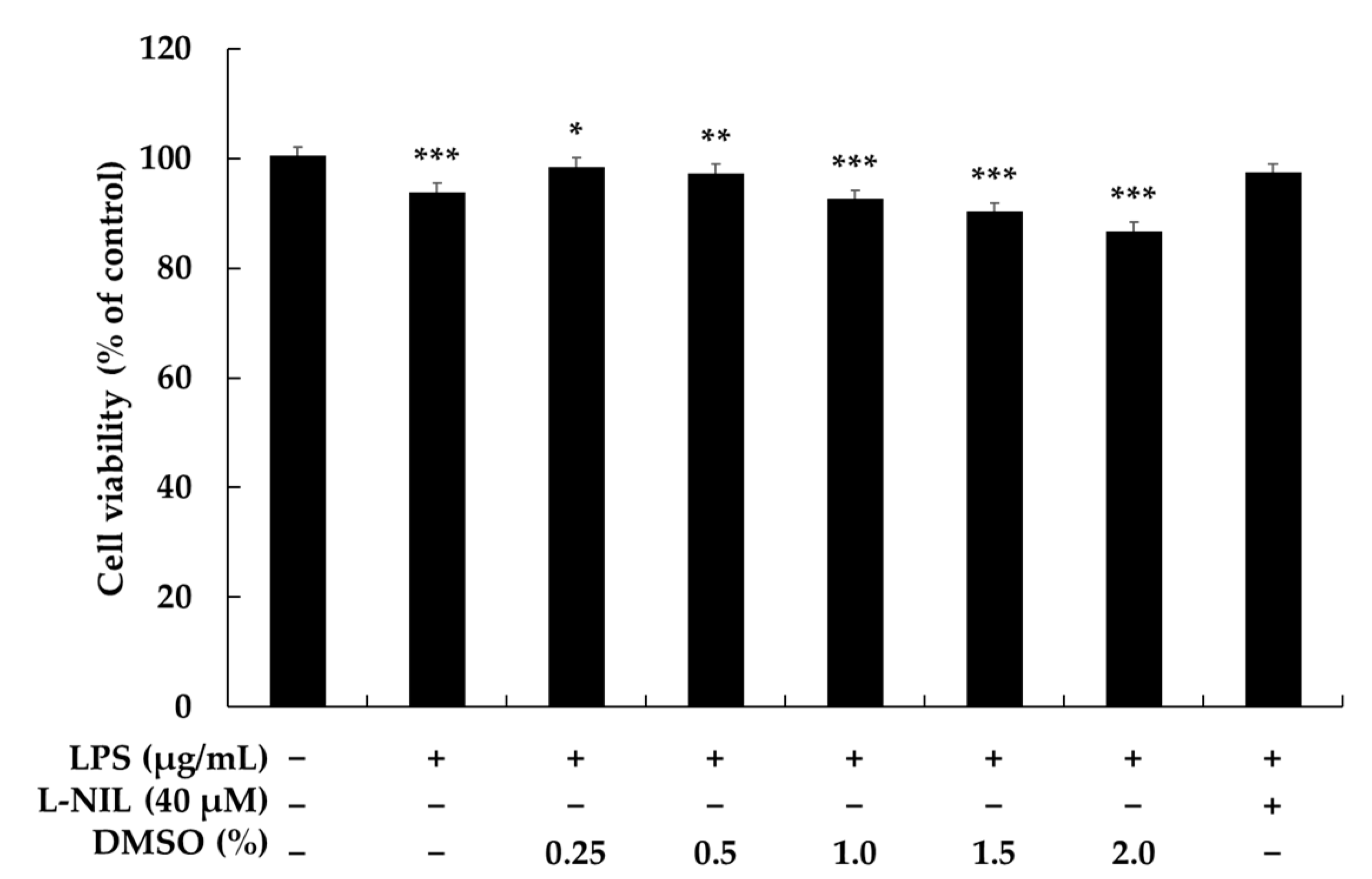
2.1. Effects of DMSO on Cell Viability
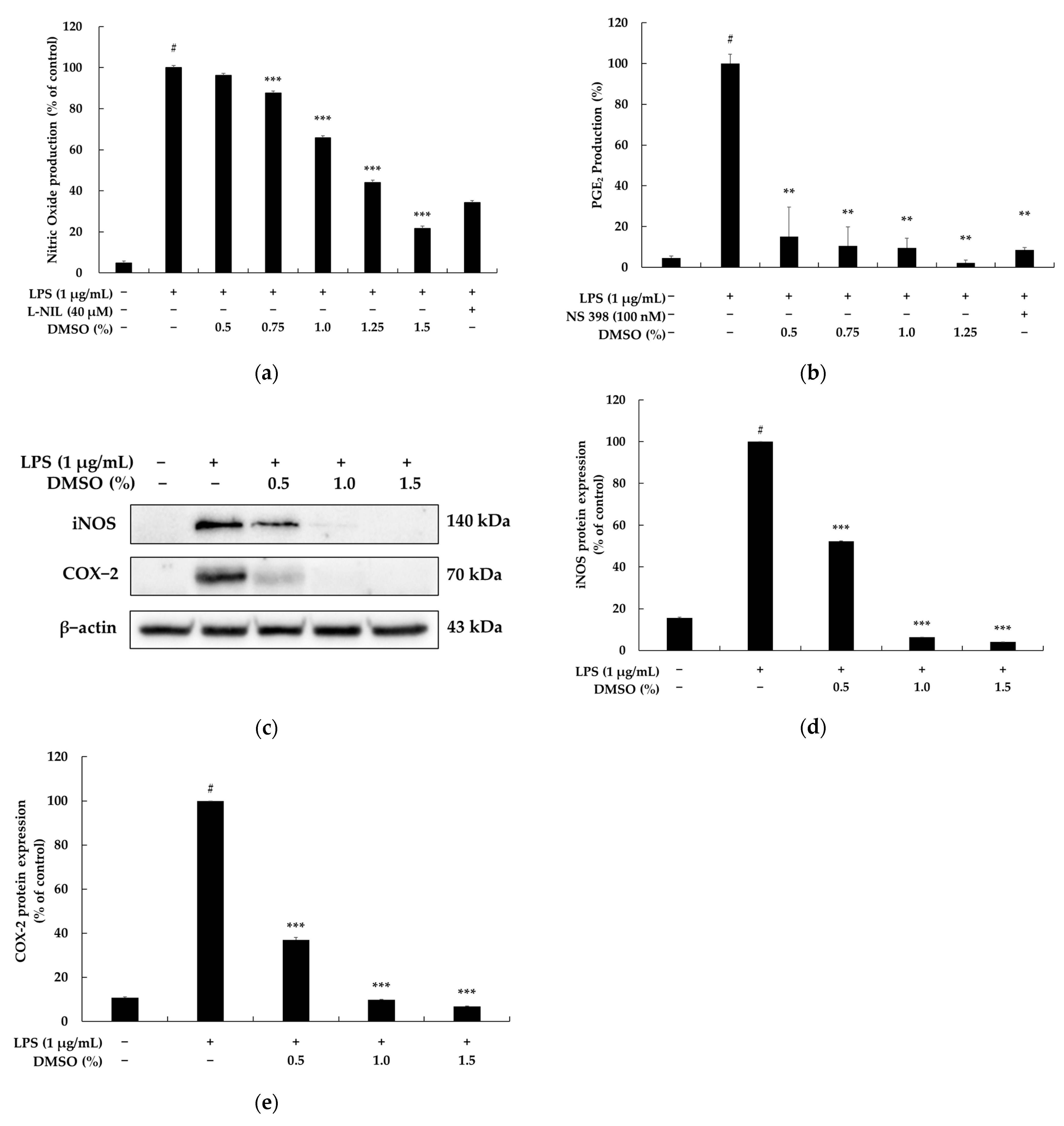
2.2. Effects of DMSO on NO Release, PGE2 Production, and Protein Expression in LPS-Stimulated RAW 264.7 Macrophages
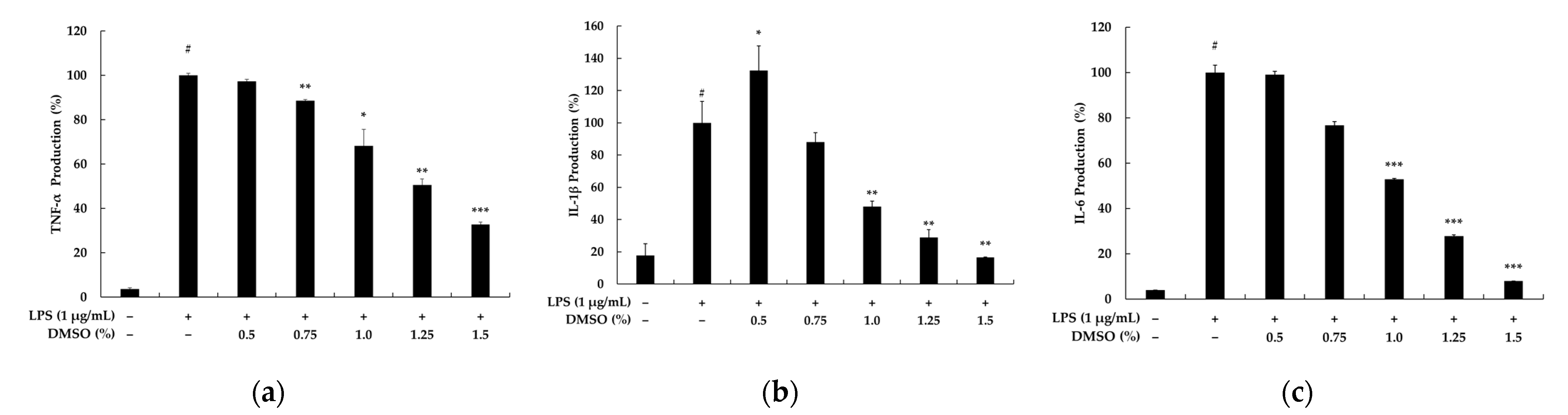
2.3. Effects of DMSO on Pro-Inflammatory Cytokine Secretion in LPS-Stimulated RAW 264.7 Cells
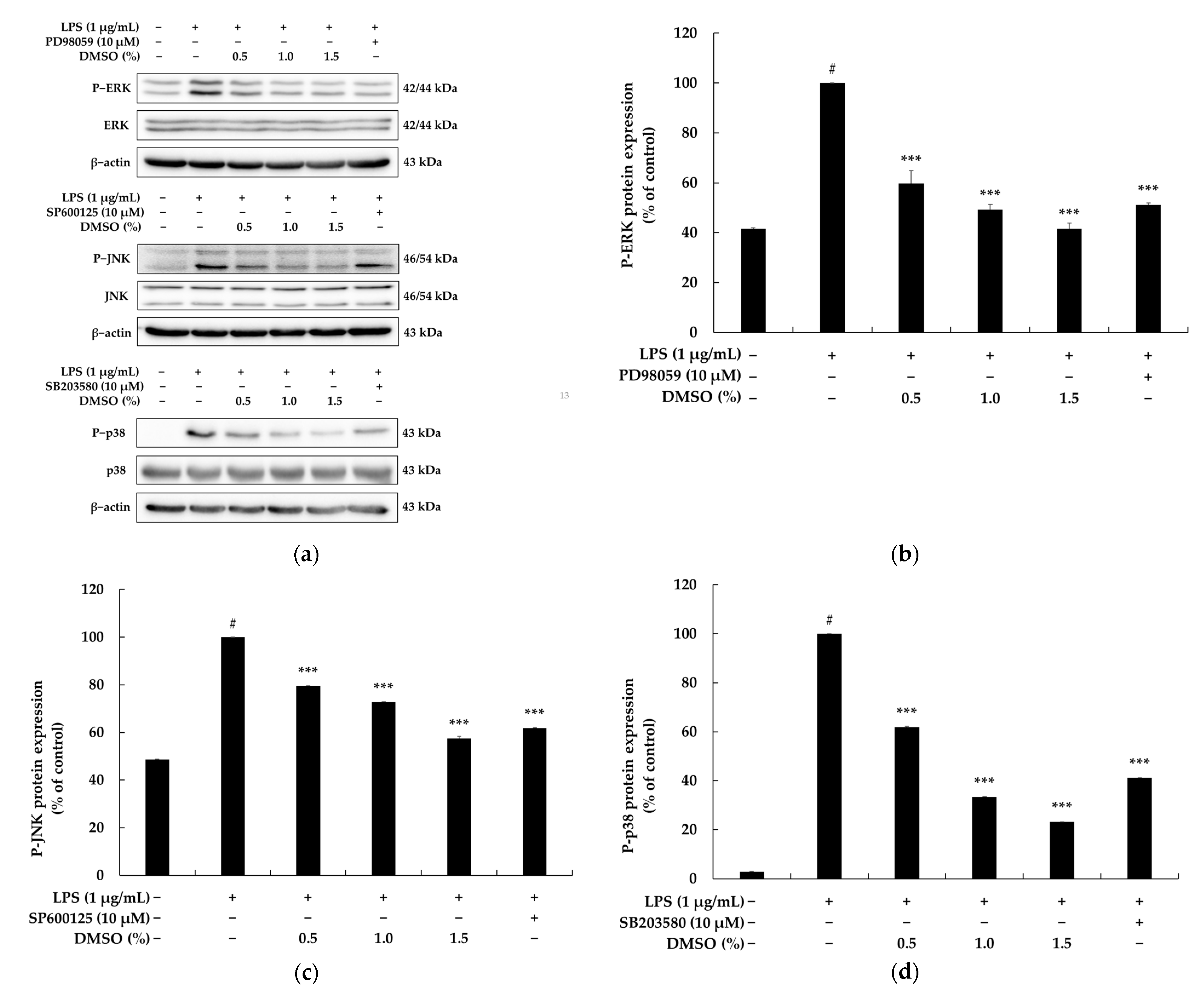
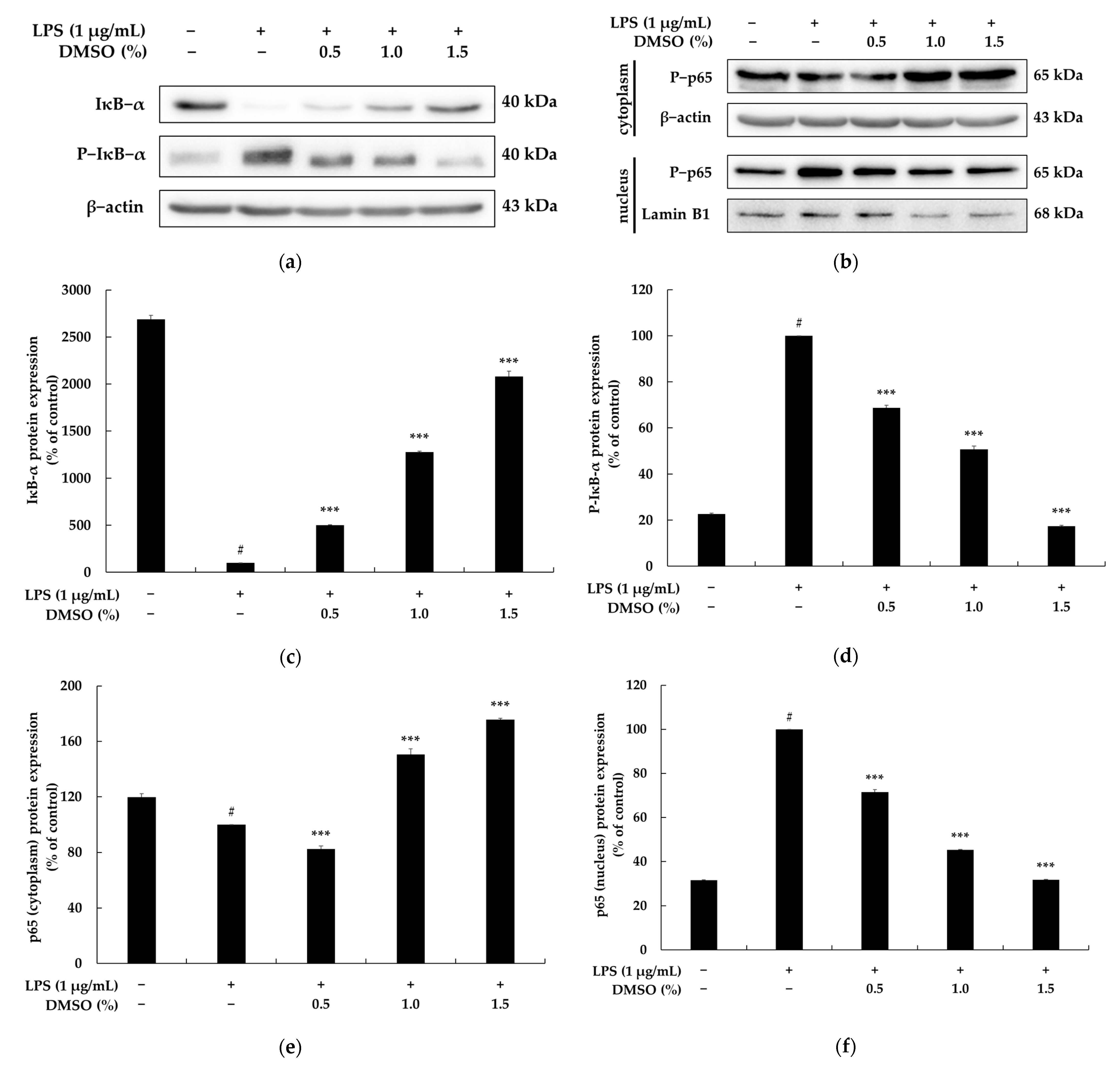
2.4. Effects of DMSO on the NF-κB and MAPK Signaling Pathways
3. Materials and Methods
3.1. Cell Culture
3.2. Cell Viability
3.3. Determination of NO Production
3.4. Determination of the PGE2, IL-1β, IL-6, and TNF-α Levels
3.5. Preparation of Nuclear and Cytoplasmic Extracts
3.6. Western Blotting
3.7. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Deol, P.; Yang, J.; Morisseau, C.; Hammock, B.D.; Sladek, F.M. Dimethyl Sulfoxide Decreases Levels of Oxylipin Diols in Mouse Liver. Front. Pharmacol. 2019, 10, 580. [Google Scholar] [CrossRef] [Green Version]
- Elisia, I.; Nakamura, H.; Lam, V.; Hofs, E.; Cederberg, R.; Cait, J.; Hughes, M.R.; Lee, L.; Jia, W.; Adomat, H.H.; et al. DMSO Represses Inflammatory Cytokine Production from Human Blood Cells and Reduces Autoimmune Arthritis. PLoS ONE 2016, 11, e0152538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajighasemi, F.; Tajik, S. Assessment of Cytotoxicity of Dimethyl Sulfoxide in Human Hematopoietic Tumor Cell Lines. Iran. J. Blood Cancer 2017, 9, 48–53. [Google Scholar]
- de Abreu Costa, L.; Henrique Fernandes Ottoni, M.; Dos Santos, M.G.; Meireles, A.B.; Gomes de Almeida, V.; de Fátima Pereira, W.; Alves de Avelar-Freitas, B.; Eustáquio Alvim Brito-Melo, G. Dimethyl Sulfoxide (DMSO) Decreases Cell Proliferation and TNF-α, IFN-γ, and IL-2 Cytokines Production in Cultures of Peripheral Blood Lymphocytes. Molecules 2017, 22, 1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Park, J.B. Evaluation of the effects of dimethylsulphoxide on morphology, cellular viability, mRNA, and protein expression of stem cells culture in growth media. Biomed. Rep. 2017, 7, 291–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bini, R.; Olivero, G.; Trombetta, A.; Castagna, E.; Cotogni, P. Effects of dimethyl sulfoxide, pyrrolidine dithiocarbamate, and methylprednisolone on nuclear factor-kappaB and heat shock protein 70 in a rat model of hemorrhagic shock. J. Trauma Acute Care Surg. 2008, 64, 1048–1054. [Google Scholar] [CrossRef]
- Kelly, K.A.; Hill, M.R.; Youkhana, K.; Wanker, F.; Gimble, J.M. Dimethyl sulfoxide modulates NF-kappa B and cytokine activation in lipopolysaccharide-treated murine macrophages. Infect. Immun. 1994, 62, 3122–3128. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Qiu, W.; Ao, X.; Li, W.; He, X.; Ao, L.; Hu, X.; Li, Z.; Zhu, M.; Luo, D.; et al. Low-concentration DMSO accelerates skin wound healing by Akt/mTOR-mediated cell proliferation and migration in diabetic mice. Br. J. Pharmacol. 2020, 177, 3327–3341. [Google Scholar] [CrossRef]
- Kang, J.K.; Kang, H.K.; Hyun, C.G. Anti-Inflammatory Effects of Spiramycin in LPS-Activated RAW 264.7 Macrophages. Molecules 2022, 27, 3202. [Google Scholar] [CrossRef]
- Kang, J.K.; Chung, Y.C.; Hyun, C.G. Anti-Inflammatory Effects of 6-Methylcoumarin in LPS-Stimulated RAW 264.7 Macrophages via Regulation of MAPK and NF-κB Signaling Pathways. Molecules 2021, 26, 5351. [Google Scholar] [CrossRef]
- Han, S.; Gao, H.; Chen, S.; Wang, Q.; Li, X.; Du, L.J.; Li, J.; Luo, Y.Y.; Li, J.X.; Zhao, L.C.; et al. Procyanidin A1 Alleviates Inflammatory Response induced by LPS through NF-κB, MAPK, and Nrf2/HO-1 Pathways in RAW264.7 cells. Sci. Rep. 2019, 9, 15087. [Google Scholar] [CrossRef] [Green Version]
- Silva, E.J.; Herrera, D.R.; Almeida, J.F.; Ferraz, C.C.; Gomes, B.P.; Zaia, A.A. Evaluation of cytotoxicity and up-regulation of gelatinases in fibroblast cells by three root repair materials. Int. Endod. J. 2012, 45, 815–820. [Google Scholar] [CrossRef]
- Kloesch, B.; Liszt, M.; Broell, J.; Steiner, G. Dimethyl sulphoxide and dimethyl sulphone are potent inhibitors of IL-6 and IL-8 expression in the human chondrocyte cell line C-28/I2. Life Sci. 2011, 89, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Hollebeeck, S.; Raas, T.; Piront, N.; Schneider, Y.J.; Toussaint, O.; Larondelle, Y.; During, A. Dimethyl sulfoxide (DMSO) attenuates the inflammatory response in the in vitro intestinal Caco-2 cell model. Toxicol. Lett. 2011, 206, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Wedner, H.J.; Bass, G. Induction of the tyrosine phosphorylation of a 66 kd soluble protein by DMSO in human peripheral blood T lymphocytes. Biochem. Biophys. Res. Commun. 1986, 140, 743–749. [Google Scholar] [CrossRef]
- Kloverpris, H.; Fomsgaard, A.; Handley, A.; Ackland, J.; Sullivan, M.; Goulder, P. Dimethyl sulfoxide (DMSO) exposure to human peripheral blood mononuclear cells (PBMCs) abolish T cell responses only in high concentrations and following coincubation for more than two hours. J. Immunol. Methods 2010, 356, 70–78. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, W.; Zheng, F.; Yu, H.; Wei, K. Xanthatin Alleviates LPS-Induced Inflammatory Response in RAW264.7 Macrophages by Inhibiting NF-κB, MAPK and STATs Activation. Molecules 2022, 27, 4603. [Google Scholar] [CrossRef]
- So, B.R.; Bach, T.T.; Paik, J.H.; Jung, S.K. Kmeria duperreana (Pierre) Dandy Extract Suppresses LPS-Induced iNOS and NO via Regulation of NF-κB Pathways and p38 in Murin Macrophage RAW 264.7 Cells. Prev. Nutr. Food Sci. 2020, 25, 166–172. [Google Scholar] [CrossRef]
- Wu, J.; Liu, B.; Mao, W.; Feng, S.; Yao, Y.; Bai, F.; Shen, Y.; Guleng, A.; Jirigala, B.; Cao, J. Prostaglandin E2 Regulates Activation of Mouse Peritoneal Macrophages by Staphylococcus aureus through Toll-Like Receptor 2, Toll-like Receptor 4, and NLRP3 Inflammasome Signaling. J. Innate Immun. 2020, 12, 154–169. [Google Scholar] [CrossRef]
- Parameswaran, N.; Patial, S. Tumor necrosis factor-α signaling in macrophages. Crit. Rev. Eukaryot. Gene Expr. 2010, 20, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peroval, M.Y.; Boyd, A.C.; Young, J.R.; Smith, A.L. A critical role for MAPK signalling pathways in the transcriptional regulation of toll like receptors. PLoS ONE 2013, 8, e51243. [Google Scholar] [CrossRef] [Green Version]
- Laird, M.H.; Rhee, S.H.; Perkins, D.J.; Medvedev, A.E.; Piao, W.; Fenton, M.J.; Vogel, S.N. TLR4/MyD88/PI3K interactions regulate TLR4 signaling. J. Leukoc. Biol. 2009, 85, 966–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Invest. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orabona, C.; Orecchini, E.; Volpi, C.; Bacaloni, F.; Panfili, E.; Pagano, C.; Perioli, L.; Belladonna, M.L. Crocus sativus L. Petal Extract Inhibits Inflammation and Osteoclastogenesis in RAW 264.7 Cell Model. Pharmaceutics 2022, 14, 1290. [Google Scholar] [CrossRef]
- Zhao, W.R.; Shi, W.T.; Zhang, J.; Zhang, K.Y.; Qing, Y.; Tang, J.Y.; Chen, X.L.; Zhou, Z.Y. Tribulus terrestris L. Extract Protects against Lipopolysaccharide-Induced Inflammation in RAW 264.7 Macrophage and Zebrafish via Inhibition of Akt/MAPKs and NF-κB/iNOS-NO Signaling Pathways. Evid.-Based Complement. Altern. Med. 2021, 2021, 6628561. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, H.; Kang, J.-K.; Ahn, K.J.; Hyun, C.-G. DMSO Alleviates LPS-Induced Inflammatory Responses in RAW264.7 Macrophages by Inhibiting NF-κB and MAPK Activation. BioChem 2023, 3, 91-101. https://doi.org/10.3390/biochem3020007
Han H, Kang J-K, Ahn KJ, Hyun C-G. DMSO Alleviates LPS-Induced Inflammatory Responses in RAW264.7 Macrophages by Inhibiting NF-κB and MAPK Activation. BioChem. 2023; 3(2):91-101. https://doi.org/10.3390/biochem3020007
Chicago/Turabian StyleHan, Hyunju, Jin-Kyu Kang, Keun Jae Ahn, and Chang-Gu Hyun. 2023. "DMSO Alleviates LPS-Induced Inflammatory Responses in RAW264.7 Macrophages by Inhibiting NF-κB and MAPK Activation" BioChem 3, no. 2: 91-101. https://doi.org/10.3390/biochem3020007
APA StyleHan, H., Kang, J. -K., Ahn, K. J., & Hyun, C. -G. (2023). DMSO Alleviates LPS-Induced Inflammatory Responses in RAW264.7 Macrophages by Inhibiting NF-κB and MAPK Activation. BioChem, 3(2), 91-101. https://doi.org/10.3390/biochem3020007