
DNA Barcoding for Scorpion Species from New Valley Governorate in Egypt Reveals Different Degrees of Cryptic Speciation and Species Misnaming

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
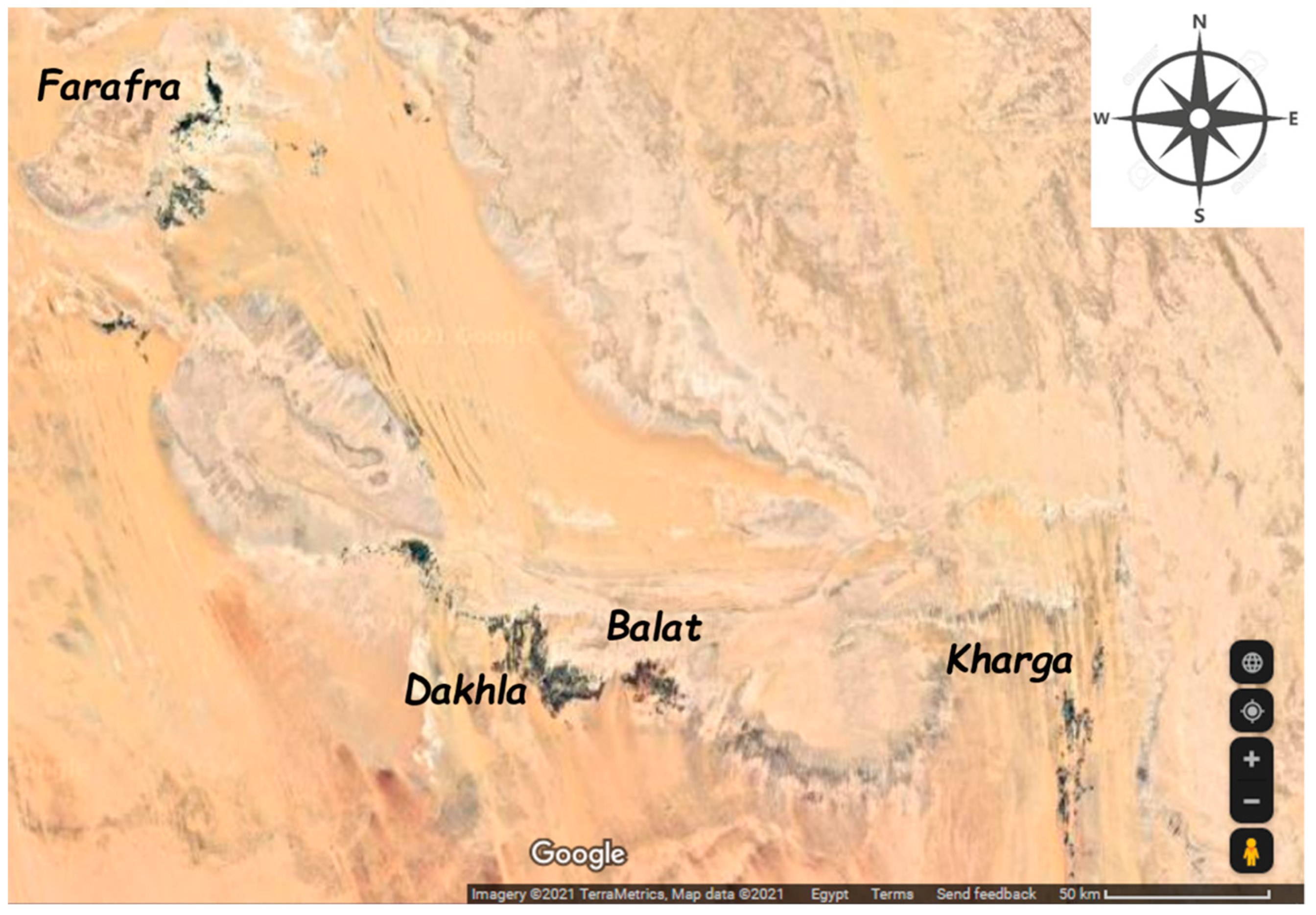
2.1. Sample Collection and Preservation
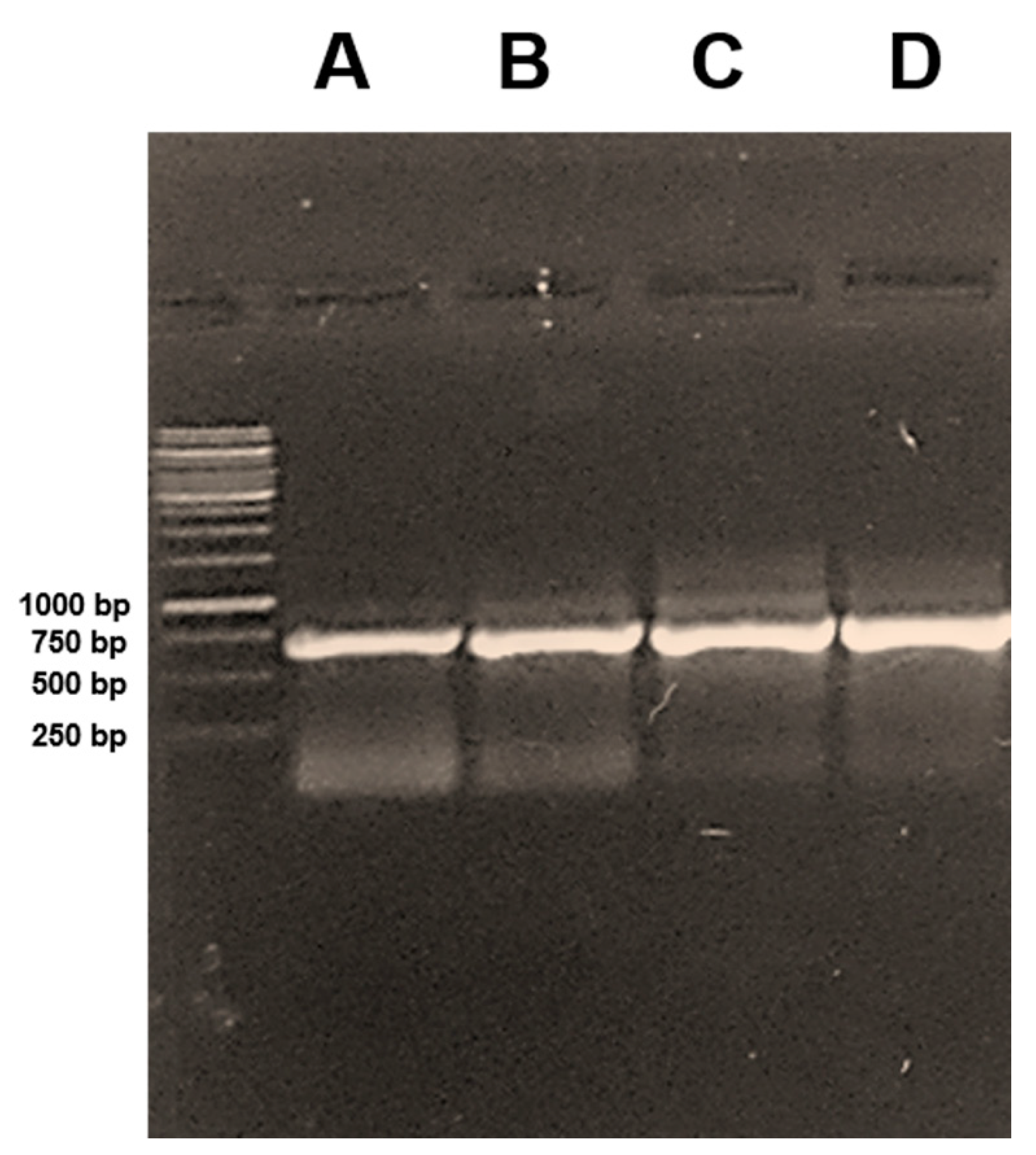
2.2. DNA Purification and COI-Based Polymerase Chain (PCR)
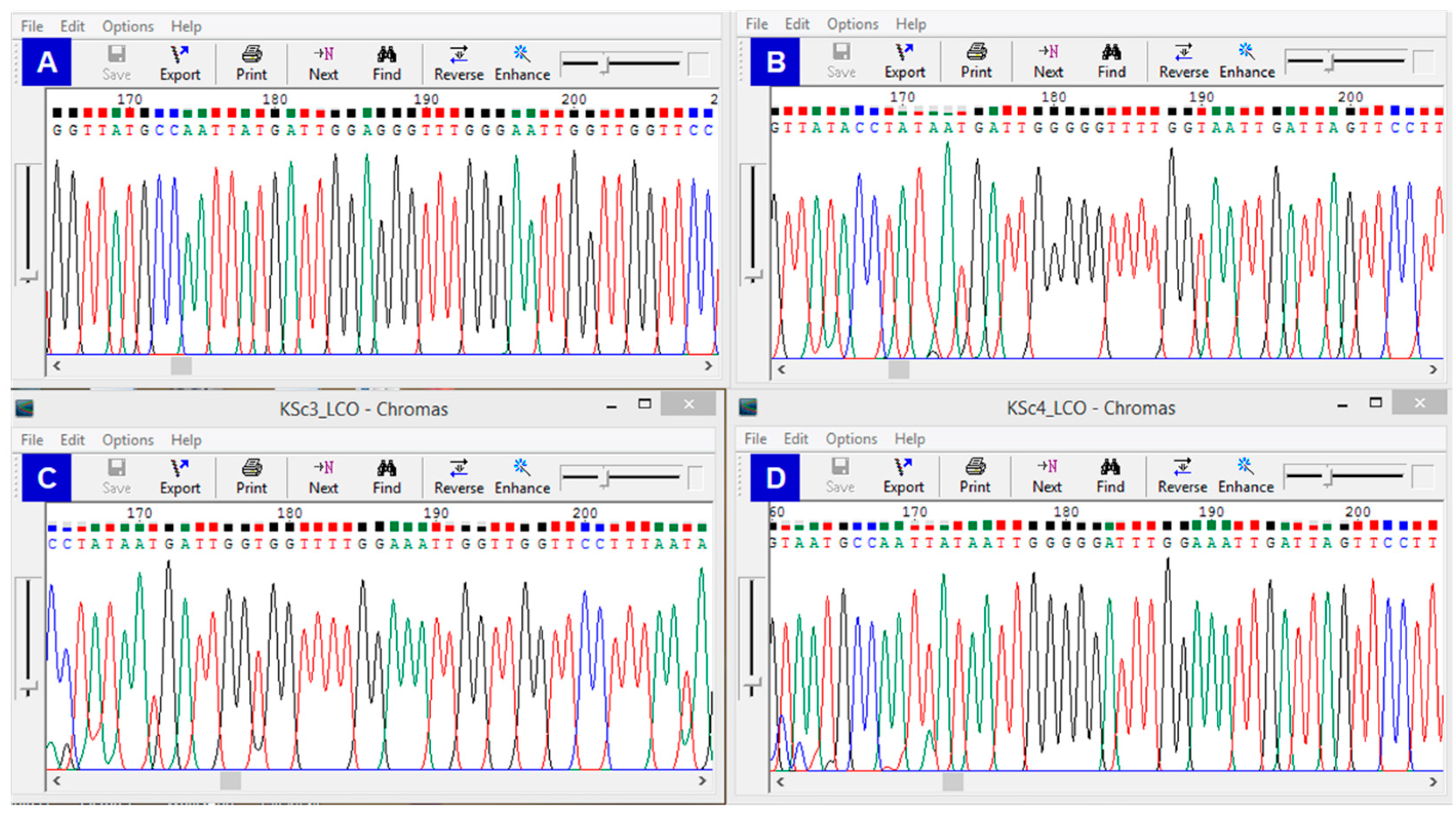
2.3. Genetic and Phylogenetic Analyses for New Valley Scorpions
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Nime, M.F.; Casanoves, F.; Mattoni, C.I. Scorpion diversity in two different habitats in the Arid Chaco, Argentina. J. Insect Conserv. 2014, 18, 373–384. [Google Scholar] [CrossRef]
- Polis, G.A.; Yamashita, T. The ecology and importance of predaceous arthropods in desert communities. In The Ecology of Desert Communities; Polis, G.A., Ed.; University of Arizona Press: Tucson, AZ, USA, 1991; Chapter 17; pp. 180–222. [Google Scholar]
- Shehab, A.H.; Amr, Z.S.; Lindsell, J.A. Ecology and biology of scorpions in Palmyra, Syria. Turk. J. Zool. 2011, 35, 333–341. [Google Scholar]
- Lighton, J.; Brownell, P.; Joos, B.; Turner, R. Low metabolic rate in scorpions: Implications for population biomass and cannibalism. J. Exp. Biol. 2001, 204, 607–613. [Google Scholar] [CrossRef]
- Stockmann, R. Introduction to Scorpion Biology and Ecology. In Scorpion Venoms; Gopalakrishnakone, P., Ferroni Schwartz, E., Possani, L., Rodríguez de la Vega, R., Eds.; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Prendini, L. Scorpion diversity and distribution in southern Africa: Pattern and process. In African Biodiversity: Molecules, Organisms, Ecosystems; Huber, B.A., Sinclair, B.J., Lampe, K.-H., Eds.; Springer: Boston, MA, USA, 2005; pp. 25–68. [Google Scholar]
- Druce, D.; Hamer, M.; Slotow, R. Patterns of millipede (Diplopoda), centipede (Chilopoda) and scorpion (Scorpionida) diversity in savanna habitats within the Greater Makalali Conservancy, South Africa. Afr. Zool. 2007, 42, 204–215. [Google Scholar] [CrossRef]
- Foord, S.H.; Gelebe, V.; Prendini, L. Effects of aspect and altitude on scorpion diversity along an environmental gradient in the Soutpansberg, South Africa. J. Arid Environ. 2015, 113, 114–120. [Google Scholar] [CrossRef]
- Chippaux, J.-P.; Goyffon, M. Epidemiology of scorpionism: A global appraisal. Acta Trop. 2008, 107, 71–79. [Google Scholar] [CrossRef]
- Saleh, M.; Younes, M.; Badry, A.; Sarhan, M. Zoogeographical analysis of the Egyptian scorpion fauna. Al-Azhar Bull. Sci. 2017, 28, 1–14. [Google Scholar] [CrossRef]
- Badry, A.; Younes, M.; Sarhan, M.M.; Saleh, M. On the scorpion fauna of Egypt, with an identification key (Arachnida: Scorpiones). Zoöl. Middle East 2018, 64, 75–87. [Google Scholar] [CrossRef]
- Sarhan, M.; Badry, A.; Younes, M.; Saleh, M. Genetic diversity within Leiurus quinquestriatus (Scorpiones: Buthidae) populations in Egypt as inferred from 16S mDNA sequence analysis. Zoöl. Middle East 2020, 66, 269–276. [Google Scholar] [CrossRef]
- Kinzelbach, R.K.; Lutz, H. Stylopid larva from the Eocene—A spotlight on the phylogeny of the stylopids (Strepsiptera). Ann. Entomol. Soc. Am. 1985, 78, 600–602. [Google Scholar] [CrossRef]
- Moustafa, A.A. Studies on Scorpions in ST. Catherine and Wadi Feiran Regions, South Sinai. Master’s Thesis, Faculty of Science, Suez Canal University, Ismailia, Egypt, 1988. [Google Scholar]
- El-Hennawy, H.K. A catalogue of the scorpions described from the Arab countries (1758–1990) (Arachnida: Scorpionida). Serket 1992, 2, 95–123. [Google Scholar]
- El-Hennawy, H.K. Updated List of Scorpions of Egypt. Serket 2014, 1–4. Available online: http://hishamelhennawycv.blogspot.com.eg/p/blog-page_19.html (accessed on 1 July 2021).
- Teruel, M.; McKinney, C.; Balsa, A.; Pascual-Salcedo, D.; Rodriguez-Rodriguez, L.; Ortiz, A.M.; Gómez-Vaquero, C.; González-Gay, M.A.; Smith, M.; Witte, T.; et al. Association of CD247 polymorphisms with rheumatoid arthritis: A replication study and a meta-analysis. PLoS ONE 2013, 8, e68295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, P.D.; Ratnasingham, S.; De Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Biol. Sci. 2003, 270, S96–S99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheir, S.K.; Galal-Khallaf, A.; Mohamed, A.H.; Mohammed-Geba, K. Morphological and molecular clues for recording the first appearance of Artemia franciscana (Kellogg, 1906) in Egypt. Heliyon 2018, 4, e01110. [Google Scholar] [CrossRef] [PubMed]
- Mohammed-Geba, K.; Sheir, S.K.; Hamed, E.A.E.A.; Galal-Khallaf, A. Molecular and morphological signatures for extreme environmental adaptability of the invasive mussel Brachidontes pharaonis (Fischer, 1870). Mol. Cell. Probes 2020, 53, 101594. [Google Scholar] [CrossRef]
- Obuid-Allah, A.H.; El-Shimy, N.A.; Mahbob, M.A.; Ali, R.S. Survey and Morphological Studies on Scorpions Inhabiting New Valley Governorate, Egypt. Egypt. Acad. J. Biol. Sci. B Zoöl. 2020, 12, 227–238. [Google Scholar] [CrossRef]
- Said, S.; Obuid-Allah, A.H.; El-Shimy, N.A.; Mahbob, M.A.; Ali, R.S. Ultrastructure Study on The Exo-Morphology of Four Species of Scorpion Inhabiting New Valley, Egypt. Egypt. Acad. J. Biol. Sci. B Zoöl. 2021, 13, 17–29. [Google Scholar] [CrossRef]
- Amr, Z.S.; Baker, M.A.A.; Al-Saraireh, M.; Warrell, D.A. Scorpions and scorpion sting envenoming (scorpionism) in the Arab Countries of the Middle East. Toxicon 2020, 191, 83–103. [Google Scholar] [CrossRef]
- Mohammed-Geba, K.; Hassab El-Nabi, S.E.; El-Desoky, M.S. Development of cytochrome-c-oxidase 1 specific primers for genetic discrimination of the European eel Anguilla anguilla (Linnaeus, 1758). J. Biosci. Appl. Res. 2016, 2, 258–262. [Google Scholar] [CrossRef]
- Wasko, A.P.; Marlio, C.; Oliveira, C.; Forresli, F. Non-destructive genetic sampling in fish. An improved method for DNA extraction from fish fins and scales. Hereditas 2003, 138, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.E.; Abdel-Baseer, K.A.; Saad, K.; Hassan, A.F.; El-Houfey, A.A. Endocrinological and biochemical changes of scorpionism in children in Upper Egypt. Ther. Adv. Endocrinol. Metab. 2015, 6, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Saad, K.; El-Hamed, M.A.A.; Abo-Elela, M.G.M.; Ahmed, A.E.; Abdel-Baseer, K.A.; Aboul-Khair, M.D.; Metwalley, K.A.; El-Houfey, A.A.; Hasan, G.M.; El-Shareef, A.M. Neurologic Complications in Children with Scorpionism: A Retrospective Study in Upper Egypt. J. Child Neurol. 2017, 32, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Bahloul, M.; Rekik, N.; Chabchoub, I.; Chaari, A.; Ksibi, H.; Kallel, H.; Damak, H.; Chaari, A.; Ben Hamida, C.; Chelly, H.; et al. Neurological complications secondary to severe scorpion envenomation. Med Sci. Monit. 2005, 11, CR196–CR202. [Google Scholar]
- Amin, R.; Faiz, A. Scorpion sting and envenomation. In Clinical Toxinology in Asia Pacific and Africa; Gopalakrishnakone, P., Faiz, A., Fernando, R., Gnanathasan, C., Habib, A., Yang, C.C., Eds.; Toxinology; Springer: Dordrecht, The Netherlands, 2014; Volume 2, pp. 455–482. [Google Scholar]
- Barros, R.M.; Pasquino, J.A.; Peixoto, L.R.; Targino, I.T.G.; Sousa, J.A.D.; Leite, R.D.S. Clinical and epidemiological aspects of scorpion stings in the northeast region of Brazil. Ciência Saúde Coletiva 2014, 19, 1275–1282. [Google Scholar] [CrossRef] [Green Version]
- Al-Asmari, A.K.; Al-Saif, A.A.; Abdo, N.M. Morphological identification of scorpion species from Jazan and Al-Medina Al-Munawara regions, Saudi Arabia. J. Venom. Anim. Toxins Incl. Trop. Dis. 2007, 13, 821–843. [Google Scholar] [CrossRef]
- Sousa, P.; Froufe, E.; Alves, P.C.; Harris, D.J. Genetic diversity within scorpions of the genus Buthus from the Iberian Peninsula: Mitochondrial DNA sequence data indicate additional distinct cryptic lineages. J. Arachnol. 2010, 38, 206–211. [Google Scholar] [CrossRef]
- Štundlová, J.; Šmíd, J.; Nguyen, P.; Šťáhlavský, F. Cryptic diversity and dynamic chromosome evolution in Alpine scorpions (Euscorpiidae: Euscorpius). Mol. Phylogenetic. Evolut. 2019, 134, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Omran, M.A.; McVean, A. Intraspecific variation in scorpion Leiurus quinquestriatus venom collected from Egypt (Sinai and Aswan deserts). J. Toxicol. Toxin Rev. 2000, 19, 247–264. [Google Scholar] [CrossRef]
- Gantenbein, B.; Largiader, C.R. The phylogeographicimportance of the Strait of Gibraltar as a gene flow barrier in terrestrial arthropods: A case study with the scorpion Buthus occitanus as model organism. Mol. Phylogenet. Evol. 2003, 28, 119–130. [Google Scholar] [CrossRef]
- Alqahtani, A.R.; Badry, A. Interspecific phylogenetic relationship among different species of the genus Buthacus (Scorpiones: Buthidae) inferred from 16S rRNA in Egypt and Saudi Arabia. Zool. Middle East 2020, 66, 178–185. [Google Scholar] [CrossRef]
- Luna-Ramirez, K.; Miller, A.D.; Rašić, G. Genetic and morphological analyses indicate that the Australian endemic scorpion Urodacus yaschenkoi (Scorpiones: Urodacidae) is a species complex. PeerJ 2017, 5, e2759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.S.; Liu, G.M.; Zhang, D.X.; Shi, C.M. Genetic analysis and ecological niche modelling delimit species boundary of the Przewalski’s scorpion (Scorpiones: Buthidae) in arid Asian inland. BioRxiv 2020, 652024. [Google Scholar] [CrossRef]
- Soltan-Alinejad, P.; Rafinejad, J.; Dabiri, F.; Onorati, P.; Terenius, O.; Chavshin, A.R. Molecular analysis of the mitochondrial markers COI, 12S rDNA and 16S rDNA for six species of Iranian scorpions. BMC Res. Notes 2021, 14, 40. [Google Scholar] [CrossRef]
- Lourenço, W.R. Scorpion incidents, misidentification cases and possible implications for the final interpretation of results. J. Venom. Anim. Toxins Incl. Trop. Dis. 2016, 22, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Hendrixson, B.E. Buthid scorpions of Saudi Arabia, with notes on other families (Scorpiones: Buthidae, Liochelidae, Scorpionidae). Fauna Arab. 2006, 21, 120. [Google Scholar]
- Lourenço, W.R. Why does the number of dangerous species of scorpions increase? The particular case of the genus Leiurus Ehrenberg (Buthidae) in Africa. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200041. [Google Scholar] [CrossRef] [PubMed]
- Sadine, S.E. New locality of Orthochirus innesi Simon, 1910 in Algeria (Scorpiones: Buthidae). Serket 2020, 17, 171–175. [Google Scholar]
- Husemann, M.; Schmitt, T.; Stathi, I.; Habel, J.C. Evolution and Radiation in the Scorpion Buthus elmoutaouakili Lourenço and Qi 2006 (Scorpiones: Buthidae) at the Foothills of the Atlas Mountains (North Africa). J. Hered. 2012, 103, 221–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kartavtsev, Y.P.; Rozhkovan, K.V.; Masalkova, N.A. Phylogeny based on two mtDNA genes (Co-1, Cyt-B) among Sculpins (Scorpaeniformes, Cottidae) and some other scorpionfish in the Russian Far East. Mitochondrial DNA Part A 2016, 27, 2225–2240. [Google Scholar] [CrossRef]
- Devi, S.R.; Thomas, A.; Rebijith, K.B.; Ramamurthy, V.V. Biology, morphology and molecular characterization of Sitophilus oryzae and S. zeamais (Coleoptera: Curculionidae). J. Stored Prod. Res. 2017, 73, 135–141. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BOLD Accession | Species | Country | GenBank Accession |
|---|---|---|---|
| GACO2126-19 | Leiurus quinquestriatus | Sudan | KX648423 |
| GACO2127-19 | KX648424 | ||
| GBCH7621-13 | JQ514258 | ||
| GBMIN116614-17 | KX648421 | ||
| GACO2125-19 | KX648422 | ||
| GBMNC71230-20 | MT636860 | ||
| GACO2124-19 | KX648420 | ||
| GBA14470-14 | Androctonus amoreuxi | Algeria | KJ538427 |
| GBA14469-14 | KJ538423 | ||
| GACO2547-19 | Morocco | KJ538294 | |
| GBA14726-14 | KJ538282 | ||
| GBA14727-14 | KJ538286 | ||
| GBA14731-14 | KJ538290 | ||
| GBA14732-14 | KJ538297 | ||
| GBA14733-14 | KJ538305 | ||
| GBA14738-14 | KJ538301 | ||
| GBA14739-14 | KJ538313 | ||
| GBA14740-14 | KJ538317 | ||
| GBA14741-14 | KJ538309 | ||
| GBA14750-14 | KJ538275 | ||
| GBA14751-14 | KJ538279 | ||
| GBA14752-14 | KJ538436 | ||
| GBA14756-14 | KJ538429 | ||
| GBA14757-14 | KJ538447 | ||
| GBA14758-14 | KJ538433 | ||
| GBA14765-14 | KJ538450 | ||
| GBA14997-14 | Tunisia | KJ538439 | |
| GBA14998-14 | KJ538443 | ||
| GBA16571-14 | Egypt | KJ538478 | |
| GBA16576-14 | KJ538492 | ||
| GBA16572-14 | KJ538480 | ||
| GBA16573-14 | KJ538483 | ||
| GBA16574-14 | KJ538486 | ||
| GBA16575-14 | KJ538489 | ||
| GBCH7634-13 | Orthochirus innesi | Origin_Unknown | JQ514244 |
| GBMNC71229-20 | Buthacus arenicola | Origin_Unknown | MT636861 |
| GBMNB48836-20 | Buthacus macrocentrus | Iraq | MT229838 |
| GBA16582-14 | Buthacus sp. | Egypt | KF548116 |
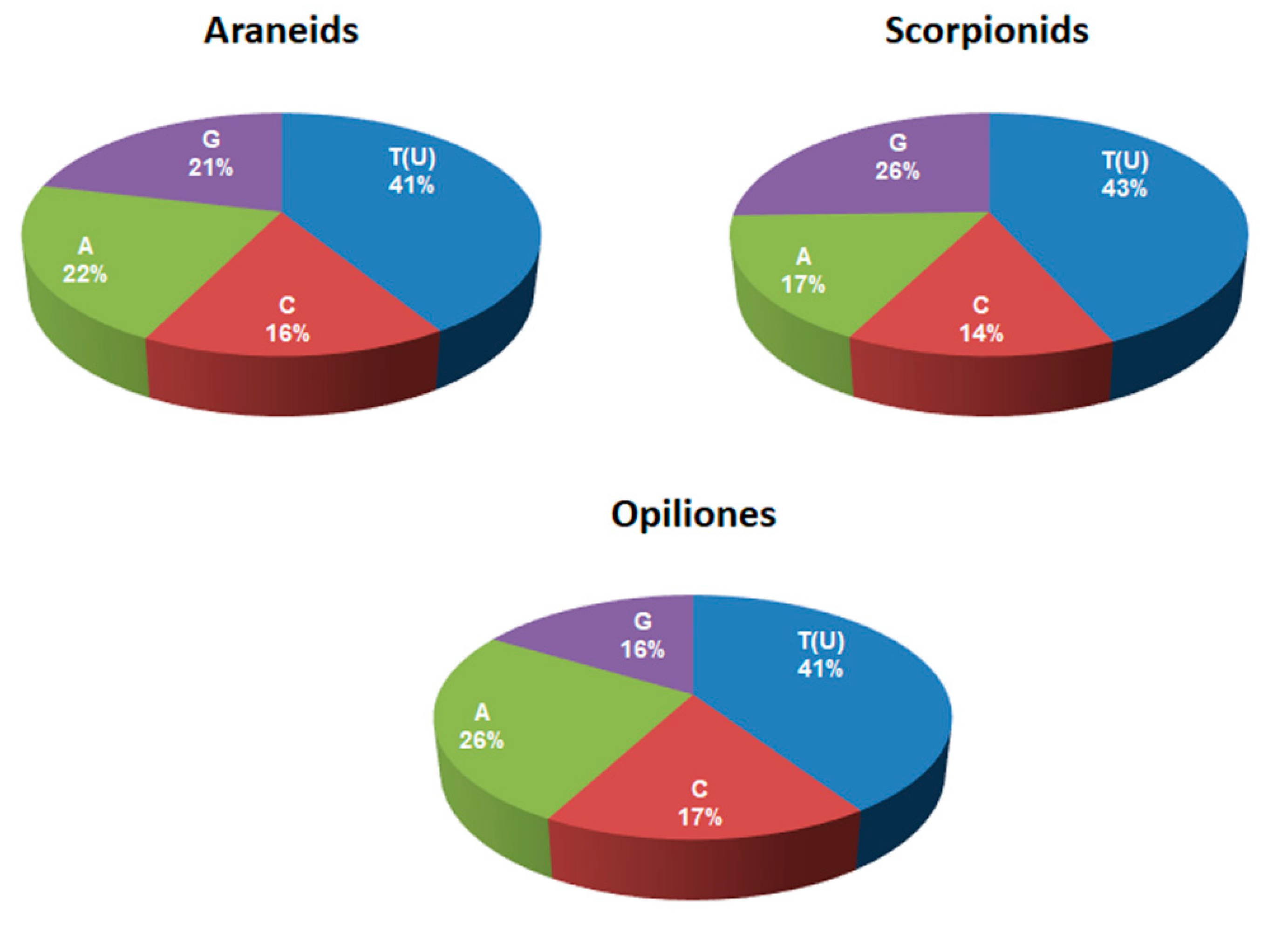
| T(U) % | C % | A % | G % | |
|---|---|---|---|---|
| Scorpiones | 43 | 14 | 17 * | 25 * |
| Aranea | 41 | 16 | 21 * | 21 * |
| Opiliones | 41 | 17 | 26 * | 16 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammed-Geba, K.; Obuid-Allah, A.H.; El-Shimy, N.A.; Mahbob, M.A.E.-M.; Ali, R.S.; Said, S.M. DNA Barcoding for Scorpion Species from New Valley Governorate in Egypt Reveals Different Degrees of Cryptic Speciation and Species Misnaming. Conservation 2021, 1, 228-240. https://doi.org/10.3390/conservation1030018
Mohammed-Geba K, Obuid-Allah AH, El-Shimy NA, Mahbob MAE-M, Ali RS, Said SM. DNA Barcoding for Scorpion Species from New Valley Governorate in Egypt Reveals Different Degrees of Cryptic Speciation and Species Misnaming. Conservation. 2021; 1(3):228-240. https://doi.org/10.3390/conservation1030018
Chicago/Turabian StyleMohammed-Geba, Khaled, Ahmad Hamed Obuid-Allah, Naser Abdellatif El-Shimy, Mohamed Abd El-Moez Mahbob, Rouwaida Saadawy Ali, and Shimaa Mohamed Said. 2021. "DNA Barcoding for Scorpion Species from New Valley Governorate in Egypt Reveals Different Degrees of Cryptic Speciation and Species Misnaming" Conservation 1, no. 3: 228-240. https://doi.org/10.3390/conservation1030018
APA StyleMohammed-Geba, K., Obuid-Allah, A. H., El-Shimy, N. A., Mahbob, M. A. E. -M., Ali, R. S., & Said, S. M. (2021). DNA Barcoding for Scorpion Species from New Valley Governorate in Egypt Reveals Different Degrees of Cryptic Speciation and Species Misnaming. Conservation, 1(3), 228-240. https://doi.org/10.3390/conservation1030018