1. Introduction
In the absence of systematic examinations of non-target effects of pesticides on key non-target insect species (such as are required by the EPA for non-target vertebrates), research on non-target insect toxicity of pesticides typically does not occur unless evidence of untoward effects is noted. The crisis with neonicotinoid insecticides and declines of pollinator populations [
1,
2,
3,
4] exemplify this pattern of problems being identified after they occur rather than being anticipated through research prior to pesticide release. Other issues, such as high sensitivity of insect predators and parasitoids to a pesticide, are potentially as ecologically disruptive as pollinator loss, but more difficult to recognize, because of the complexity of food webs and lack of direct measures of pesticide toxicities with insect natural enemies. Historically, the failure to consider impacts of insecticides on natural enemies has led to unintended consequences including the resurgence of pest populations (released from the impact of natural enemies) and the creation of secondary pest species [
5] (species that only reached pest status after natural enemies were eliminated by insecticide treatment).
Although the role of pesticides in the decline of vertebrate species is well known, are pesticides contributing to the decline and extinction of insect species? In our work on the ecology and conservation of the federally endangered Salt Creek tiger beetle,
Cicindela nevadica lincolniana, we became concerned that pesticide use might be contributing to the beetle’s decline. The most recent review of tiger beetle conservation mentions herbicide use in the context of habitat restoration, but the direct effects of herbicides and insecticides on tiger beetles are unexamined [
6].
Salt Creek tiger beetles lived in association with salt marshes of the eastern Great Plains, and now occur primarily in a single remnant salt marsh in the Little Salt Creek watershed, in Lancaster County, Nebraska. Tiger beetles are active predators as adults, but as larvae they are primarily sessile and live in burrows where they wait for prey to walk within striking distance. A complex of tiger beetle species occurs in association with salt marshes, and their burrows are typically located near water sources and subject to seasonal flooding [
7]. Given this life history, salt marsh tiger beetles, including
C. n. lincolniana, are potentially exposed to direct pesticide use associated with surrounding agricultural fields and indirect pesticide residues in water from agricultural and urban sources. Beyond this specific situation and species, we note that many insect predators within and outside of agricultural settings are predaceous as immatures.
Our original intention was to see if tiger beetle susceptibility is such that pesticides could be a factor in the decline of
C. n. lincolniana and a risk to insect predators generally. Another of the salt marsh tiger beetle species,
C. circumpicta, is not endangered and because it shares most life history requirements with
C. n. lincolniana [
8], we chose it as a suitable experimental surrogate for
C. n. lincolniana. Our objective was to examine two classes of insecticide (a pyrethroid, bifenthrin, and a neonicotinoid, imidacloprid) and one herbicide (glyphosate) based on their wide use, to see if tiger beetle susceptibility is such that pesticides could be a factor in the decline of
C. n. lincolniana, and a risk to insect predators generally. Here, we report not only that pyrethroids and neonicotinoids are potential factors in the decline of
C. n. lincolniana, but also that tiger beetle (and likely many predacious insect species) are susceptible to these chemicals at levels 100–1000 times lower than have been shown to impact insect pollinators.
2. Materials and Methods
2.1. Experimental Materials
Obtaining sufficient numbers of predaceous insects for dose–response experiments is often difficult and usually requires captive rearing; however, by field collecting pre-ovipositional females and rearing larvae from eggs, we were able to obtain sufficient larvae of the salt marsh tiger beetle species Cicindela circumpicta (a surrogate for the endangered C. n. lincolniana) to conduct dose–response studies with three pesticides. Our use of C. circumpicta in examining pesticide toxicity to non-target predators and the endangered C. n. lincolniana was based on opportunity (in obtaining adequate numbers), need (particularly in examining larval susceptibility), and suitability, because C. circumpicta is the largest of the Salt Marsh tiger beetles. In using a larger species, results are logically applicable to smaller species (absent any physiological differences).
Cicindela circumpicta adults were collected at Arbor Lake Wildlife Management Area, north of Lincoln, Nebraska. Adults were placed in containers containing Petri dishes filled with topsoil. Adults were fed pinhead crickets (Achetus domesticus) for a period of 1 week and then they were removed from the container. Cicindela circumpicta eggs hatched in the lab and were placed in individual plastic containers containing soil where they were maintained on a diet of wingless fruit flies (Drosophila melanogaster) until the bioassays were conducted (within 14 days of the first hatched larvae). Because the numbers of resulting first instars were limited, we modified our experimental designs to obtain maximum information. Larvae were only used once.
2.2. Bioassays
Bioassays were conducted within two weeks of hatching. Three different pesticides were chosen for the experiments based on their wide usage and modes of action. Bifenthrin (2-methylbiphenyl-3-ylmethyl (Z)-(1RS,3RS)-3-(2-chloro-3,3,3-trifluoroprop-l-enyl)- 2,2 dimethylcyclopropanecarboxylate) is a pyrethroid insecticide and axionic excitoxin used for urban and agricultural pests [
9]. Imidacloprid (1-(6-chloro-3-pyridylmethyl)-N-nitroimidazolidin-2-ylideneamine) is a neonicotinoid insecticide that attacks post-synaptic nicotinic acetylcholine receptors [
9]. Imidacloprid is used as a systemic insecticide, taken up by plants to poison herbivorous insects, and as a topical insecticide used on domestic animals against insect ectoparasites. Glyphosate (N(phosphonomethyl)glycine) is a herbicide and inhibits EPSP synthase in plants, resulting in fatal reductions in aromatic amino acids [
9]. Glyphosate is one of the most widely used pesticides in the world.
Three controls were used to determine if the pesticide carrier (acetone or distilled water) had any influence on mortality. Thirty C. circumpicta larvae were placed in control scintillation vials (1 larva/vial): 10 larvae in empty vials, 10 larvae in vials with distilled water, and 10 larvae vials treated with acetone. The larvae were checked for mortality after 48 h, and the survival rate was 100% in all controls at all times.
Serial dilutions of pesticides were made from technical grade (98% a.i.) pesticides. Bifenthrin and imidacloprid dilutions were prepared from starting solutions of 1.0 mg chemical/1 mL acetone, and glyphosate dilutions were prepared from a starting solution of 1.0 mg chemical/1 mL distilled water. Serial dilutions (treatments) ranged from 0.98 × 10−4 g pesticide/L carrier to 0.98 × 10−13 g pesticide/L carrier.
Each vial received 1 mL of solution (pesticide or control), vials were capped and placed horizontally on a vial roller for 10 min. Vials were then placed upright and uncapped so acetone or water would evaporate. When evaporation was completed, one first stage C. circumpicta larva was placed in each vial, and vials were sealed. Vials were laid on their side and placed on the counter at room temp (21 °C). Larvae were observed for signs of toxicity at 24 and 48 h, with specific observations recorded as Alive (A) = active movement forward; Movement (M) = movement forward with tapping stimulus; Dead (D) = no movement. Death was confirmed by removing larvae from vials and observing them for roughly one hour.
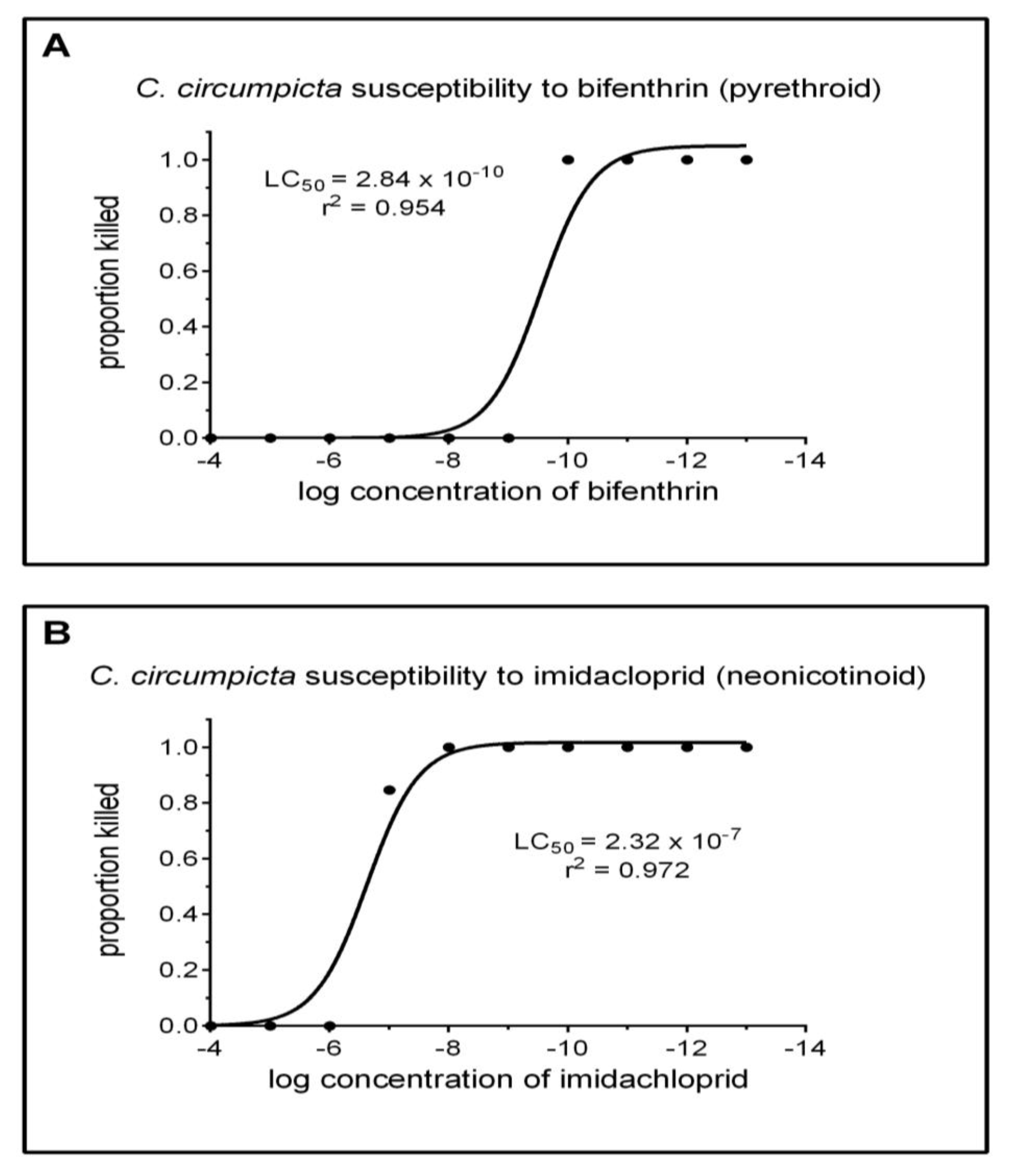
Two sets of bioassays were conducted. Initially, each pesticide was examined in 12 treatments (10 pesticide concentrations treatments, 1 control treatment with carrier (acetone or water, and 1 untreated control) with 3 replications. Observations were made over 48 h. Based on these observations, a second round of bioassays was conducted for bifenthrin and imidacloprid. Treatments were identified that were near the LC50 from the first bioassay, and concentrations near these points were examined with greater resolution (by using 10 ten replications). For bifenthrin, these concentrations were 0.98 × 10−10 g pesticide/L carrier to 0.98 × 10−13 g pesticide/L carrier; for imidacloprid, concentrations were 0.98 × 10−6 g pesticide/L carrier to 0.98 × 10−9 g pesticide/L carrier. No additional bioassays were conducted with glyphosate because no mortality occurred in the first bioassay.
After bioassays, all surviving larvae were placed in plastic cups filled with topsoil. After larvae made initial burrows, they were fed a diet of fruit flies. Observations were made and recorded when burrows were open and closed. All larvae that were alive at the end of the bioassay developed into 2nd instars after a 4-month period.
2.3. Analysis
The experimental design included replicated doses varying by orders of magnitude and replicated untreated controls. Data from both bioassays were pooled by pesticide and were analyzed by non-linear regression, specifically to determine dose–response relationships. Because responses were scored as either alive or dead, these were summarized by comparing all replications versus the controls; the analysis was based on the proportion dead. The specific form of the model used was:
where
a = minimum value,
b = maximum value,
LogLC50 = log of the lethal concentration for 50% of the test subjects,
x = log of the dose, and
y = the percentage of individuals killed at a given dose (see GraphPad Prism Curve Fitting Guide, Equation: log(agonist vs. response) for more details). The regression was conducted with GraphPad Prism 6.0 software, and the regression was constrained by setting
b = >0 and <100. Because no individuals died in the glyphosate treatments, no analysis was necessary for that treatment.
We obtained 250 first stage larval C. circumpicta for our bioassays, so all subsequent experimental design decisions reflected this limitation. No larvae died in any of our controls (empty, acetone, or distilled water), indicating handling was not a cause of death. Similarly, no larval deaths occurred in the controls of any bioassays.
4. Discussion
Regarding our original objective, the association of pesticides and endangered species usually focuses on historical examples with birds, chlorinated hydrocarbon insecticides, and bioaccumulation. Results—here using C. circumpicta as a model—indicate that tiger beetles are highly sensitive to pesticide concentrations that can and do occur in the environment. The Salt Creek tiger beetle over the past 25 years has undergone dramatic reductions in numbers and distributions, to the point that it now exists in only 200–300 individuals and is federally listed as endangered. Many former habitats were potentially exposed to pyrethroid and neonicotinoid insecticides, so we cannot exclude them as factors in decline of the beetle, along with habitat loss through urbanization, light pollution, and climate change.
Of specific concern from these data is the susceptibility of tiger beetles to neonicotinoids. While the LC
50 determined for
C. circumpicta to imidacloprid was in the tenth of a ppm range—rather than tenth of a ppb as with bifenthrin—our results indicate
C. circumpicta is 1000 times more sensitive than larval cocindelids [
13]. Because the cicindelids are close relatives to carabids, the high sensitivity of larval
C. circumpicta noted here would likely apply to many carabid species. Studies on carabid sensitivity to neonicotinoids have focused on environmental exposure (predators and prey) rather than direct toxicity (e.g., [
14,
15]), but declines in carabid numbers have been noted. Most of the current attention, debate, and research on neonicotinoids is focused on pollinator species, but the direct susceptibility data for
C. circumpicta indicates the potential for significant mortality to insect predators at concentrations orders of magnitude less than with effects noted for pollinators [
4].
Setting aside the specific toxicity of the neonicotinoids, their wide-spread use as systemic insecticides runs counter to a core principle of pest management (and common sense): avoid unnecessary selection pressures on pests and natural enemies [
5]. In the U.S., the use of neonicotinoid seed treatments has tripled in the last decade and has grown at an alarming rate in the Midwest [
16]. It might be argued that application as a systemic insecticide (inside a plant) mitigates exposure of non-target natural enemies, but data from run-off studies show that these compounds occur in the environment at toxic levels to non-target species. Specifically, based on both stream samples from agriculture-intensive areas [
16] and samples taken across habitats [
17], measured neonicotinoids concentrations from agricultural runoff routinely exceed the LC
50 reported here.
Although it can be extraordinarily difficult to assign cause and effect in ecological systems, examples of drastic declines in carabids have been documented, such as half of all carabid species in the United Kingdom showing a 30% loss in numbers from 1994–2008 [
18]. Whether or not such declines are associated with neonicotinoid use remains to be seen, but the high sensitivity of
C. circumpicta to imidacloprid (as an index of potential sensitivity of predaceous beetle larvae in general) strongly suggests that the ecological impact of the neonicotinoids extends well beyond pollinator concerns.
Our results with C. circumpicta point to a need for results to be replicated in the future with more tests and other pesticidal compounds, both within and across pesticide classes. Additionally, other predaceous insects, including other cicindelid species, should be examined. To date, no single model insect predator species has been identified for pesticide testing, but if responses of C. circumpicta are consistent with other predators, and if artificial colonies can be established, it could be a suitable candidate. Long generation times (e.g., 1 gen/year with C. circumpicta) and lower fecundity compared to herbivorous species do present potential limitations with C. circumpicta and many other predaceous insects. Consequently, we think it is appropriate to think of replication numbers in testing predaceous insects in terms of numbers used with vertebrate models rather than numbers typical with insect models. For instance, here we had a total n of 250 from field collecting adult C. circumpicta and rearing larvae, which is small compared with most herbivore insect testing, but robust compared with most vertebrate species testing.
Regarding potential risk to endangered species, our results with C. circumpicta are concerning relative to the conservation of the endangered C. nevadica lincolniana. Larvae of C. nevadica lincolniana live in burrows routinely exposed to water with agricultural runoff. Of the two species, C. circumpicta is substantially larger than C. nevadica lincolniana; thus, based on mass, there is no reason to suppose that C. nevadica lincolniana would show any greater resistance to pesticide toxicity than C. circumpicta. While habitat loss has been the greatest contributor to the decline of C. nevadica lincolniana, degradation of the remaining habitat through pesticide exposure represents a continuing risk.






{kind=link}