
Enhancing Bayesian Approaches in the Cognitive and Neural Sciences via Complex Dynamical Systems Theory



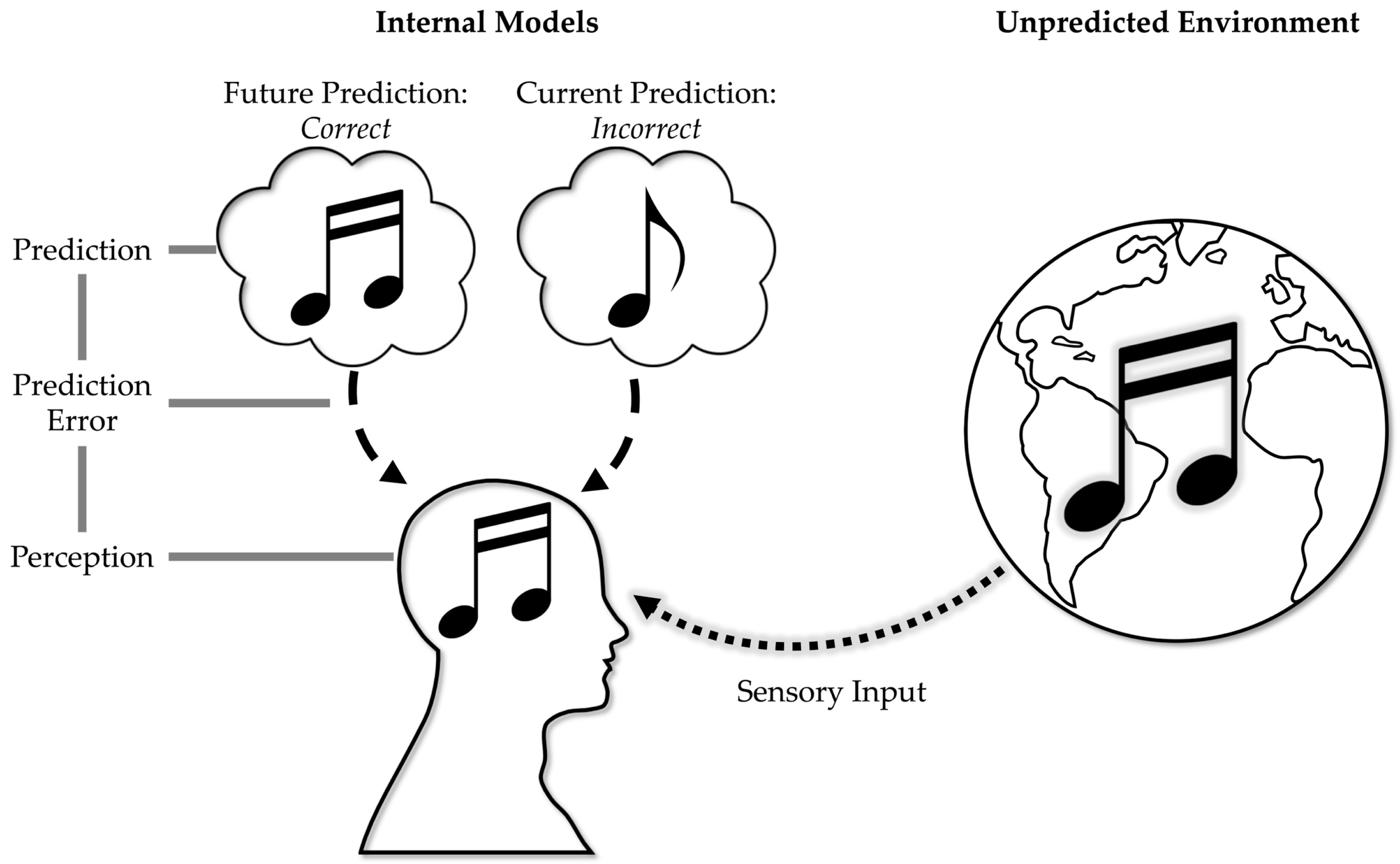{kind=link}
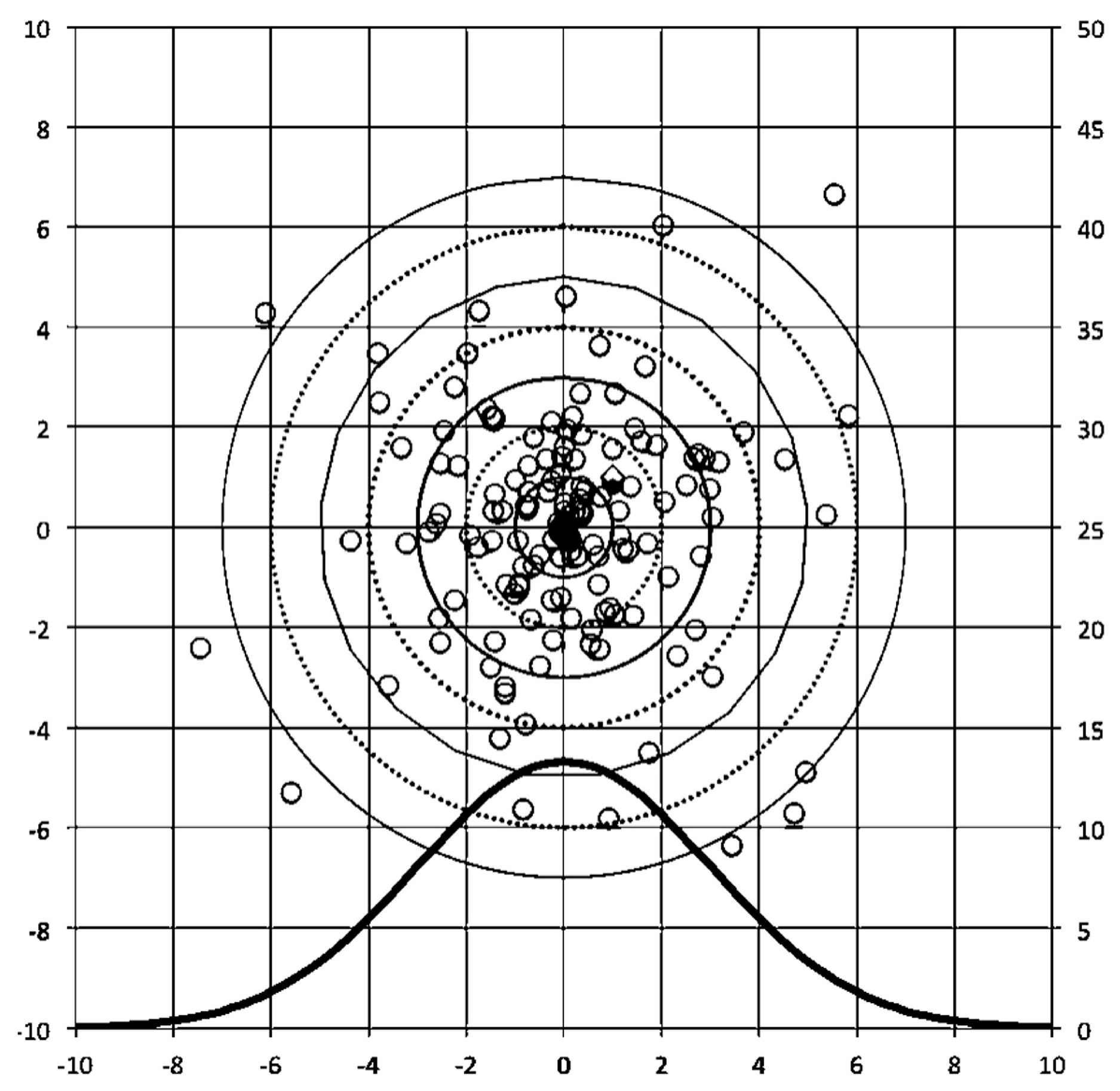{kind=link}
{kind=link}
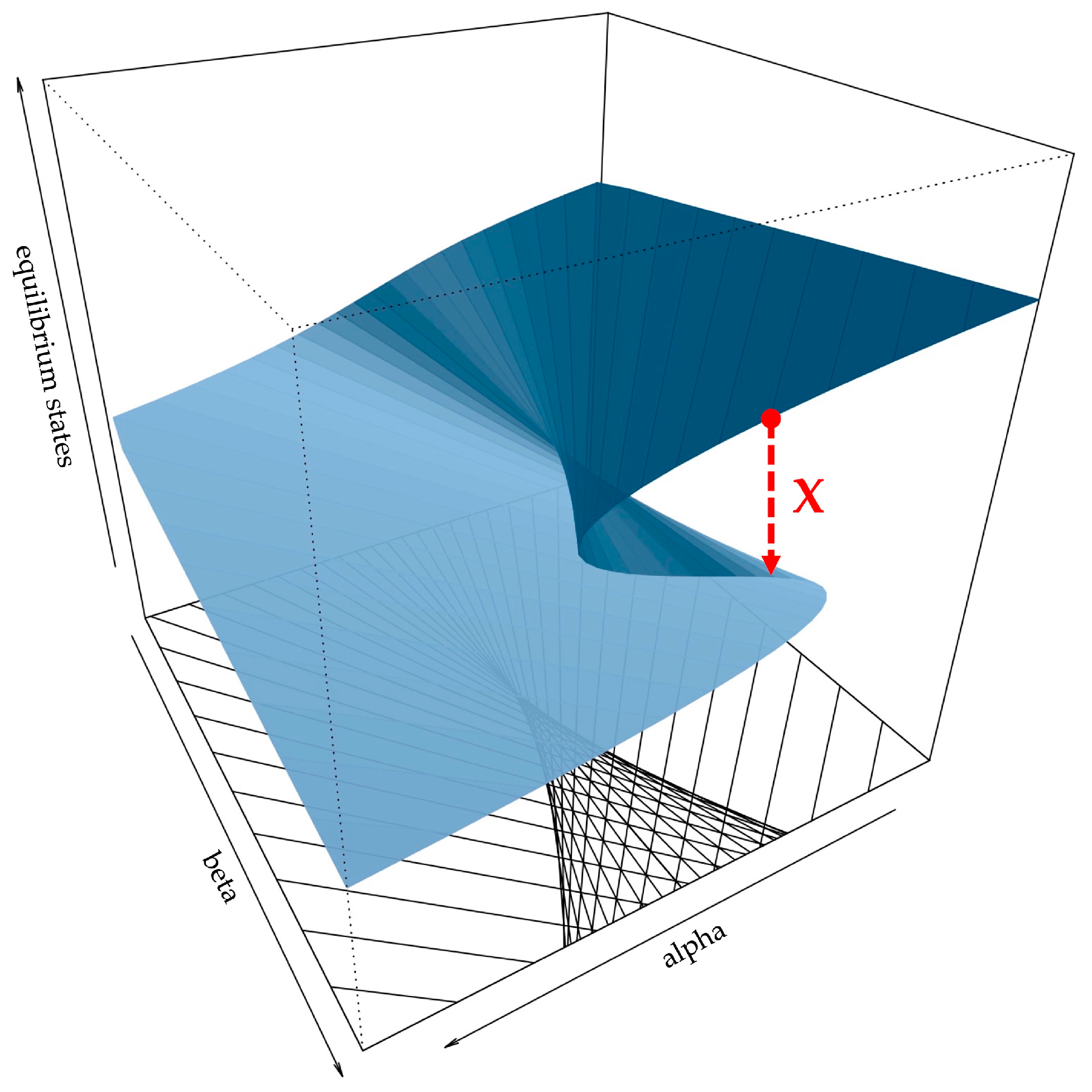{kind=link}
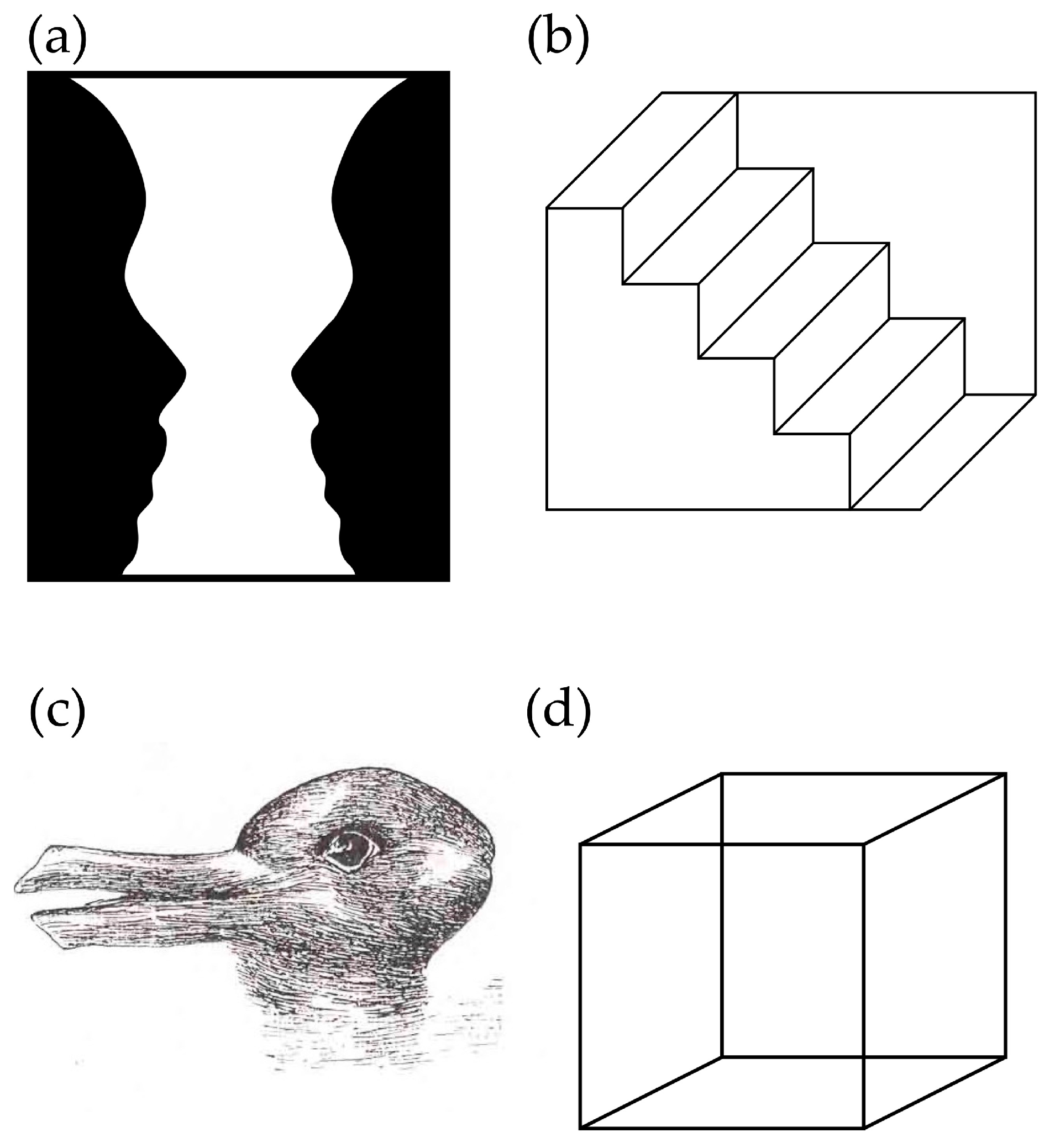{kind=link}
Abstract
:1. Introduction
2. Bayesianism in the Cognitive and Neural Sciences
2.1. A Brief Introduction to Bayes’ Theorem
2.2. Linearity and Its Consequences for Bayesian Modeling
2.2.1. Examples of Linearity in Bayesian Modeling
2.2.2. Examples of Noise in Bayesian Modeling
3. Complex Dynamical Systems Theory
4. Complex Dynamical Bayesianism
5. Challenges and Future Directions
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- May, R.M. Simple mathematical models with very complicated dynamics. Nature 1976, 261, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Bressler, S.L.; Kelso, J.A.S. Coordination dynamics in cognitive neuroscience. Front. Neurosci. 2016, 10, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friston, K. The free-energy principle: A unified brain theory? Nat. Rev. Neurosci. 2010, 11, 127–138. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, A.R. Towards a network theory of cognition. Neural Netw. 2000, 13, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Edelman, G.M. Neural Darwinism: The Theory of Neuronal Group Selection; Basic Books: New York, NY, USA, 1987. [Google Scholar]
- Anderson, M.L. Neural reuse: A fundamental organizational principle of the brain. Behav. Brain Sci. 2010, 33, 245–266. [Google Scholar] [CrossRef] [Green Version]
- Clark, A. Surfing Uncertainty: Prediction, Action, and the Embodied Mind; Oxford University Press: New York, NY, USA, 2016. [Google Scholar]
- Doya, K.; Ishii, S.; Pouget, A.; Rao, R.P. (Eds.) Bayesian Brain: Probabilistic Approaches to Neural Coding; The MIT Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Rescorla, M. Bayesian modeling of the mind: From norms to neurons. Wiley Interdiscip. Rev. Cogn. Sci. 2020, 1, e1540. [Google Scholar] [CrossRef]
- Shams, L.; Beierholm, U. Bayesian causal inference: A unifying neuroscience theory. Neurosci. Biobehav. Rev. 2022, 134, 1–14. [Google Scholar] [CrossRef]
- Haker, H.; Schneebeli, M.; Stephan, K.E. Can Bayesian theories of autism spectrum disorder help improve clinical practice? Front. Psychiatry 2016, 7, 107. [Google Scholar] [CrossRef] [Green Version]
- Nave, K.; Deane, G.; Miller, M.; Clark, A. Wilding the predictive brain. Wiley Interdiscip. Rev. Cogn. Sci. 2020, 11, e1542. [Google Scholar] [CrossRef]
- Jacobsen, D.J.; Hansen, L.K.; Madsen, K.H. Bayesian model comparison in nonlinear BOLD fMRI hemodynamics. Neural Comput. 2008, 20, 738–755. [Google Scholar] [CrossRef]
- Naselaris, T.; Prenger, R.J.; Kay, K.N.; Oliver, M.; Gallant, J.L. Bayesian reconstruction of natural images from human brain activity. Neuron 2009, 63, 902–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyas, S.; O’Shea, D.J.; Ryu, S.I.; Shenoy, K.V. Causal role of motor preparation during error-driven learning. Neuron 2020, 106, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Feldman, J. Bayesian models of perceptual organization. In The Oxford Handbook of Perceptual Organization; Wagemans, J., Ed.; Oxford University Press: New York, NY, USA, 2015; pp. 1008–1016. [Google Scholar]
- Kersten, D.; Mamassian, P.; Yuille, A. Object perception as Bayesian inference. Annu. Rev. Psychol. 2004, 55, 271–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kording, K.P.; Wolpert, D.M. Bayesian decision theory in sensorimotor control. Trends Cogn. Sci. 2006, 10, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.J.; Kording, K.P.; Goldreich, D. Bayesian Models of Perception and Action: An Introduction; The MIT Press: Cambridge, MA, USA, 2023; Draft Version 3 (May 5, 2022); Available online: http://www.cns.nyu.edu/malab/bayesianbook.html (accessed on 19 December 2022).
- Parr, T.; Friston, K.J. The anatomy of inference: Generative models and brain structure. Front. Comput. Neurosci. 2018, 12, 90. [Google Scholar] [CrossRef]
- Friston, K. Embodied inference and spatial cognition. Cogn. Process. 2012, 13, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Friston, K.; FitzGerald, T.; Rigoli, F.; Schwartenbeck, P.; Pezzulo, G. Active inference: A process theory. Neural Comput. 2017, 29, 1–49. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M. Bayesian cognitive science, predictive brains, and the nativism debate. Synthese 2018, 195, 4817–4838. [Google Scholar] [CrossRef] [Green Version]
- Friston, K. The history of the future of the Bayesian brain. NeuroImage 2012, 62, 1230–1233. [Google Scholar] [CrossRef] [Green Version]
- Knill, D.C.; Pouget, A. The Bayesian brain: The role of uncertainty in neural coding and computation. Trends Neurosci. 2004, 27, 712–719. [Google Scholar] [CrossRef]
- Kording, K.P. Bayesian statistics: Relevant for the brain? Curr. Opin. Neurobiol. 2014, 25, 130–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Rao, R.P.N. Predictive coding. Wiley Interdiscip. Rev. Cogn. Sci. 2011, 2, 580–593. [Google Scholar] [CrossRef] [PubMed]
- Spratling, M.W. Predictive coding as a model of cognition. Cogn. Process. 2016, 17, 279–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A. Whatever next? Predictive brains, situated agents, and the future of cognitive science. Behav. Brain Sci. 2013, 36, 181–204. [Google Scholar] [CrossRef]
- Clark, A. Perception as Controlled Hallucination: Predictive Processing and the Nature of Conscious Experience. Edge. Available online: https://www.edge.org/conversation/andy_clark-perception-as-controlled-hallucination (accessed on 6 June 2019).
- Hohwy, J. New directions in predictive processing. Mind Lang. 2020, 35, 209–223. [Google Scholar] [CrossRef]
- Wiese, W.; Metzinger, T. Vanilla PP for philosophers: A primer on predictive processing. In Philosophy and Predictive Processing (1); Metzinger, T., Wiese, W., Eds.; MIND Group: Frankfurt am Main, Germany, 2017. [Google Scholar] [CrossRef]
- Yon, D.; Frith, C.D. Precision and the Bayesian brain. Curr. Biol. 2021, 31, R1026–R1032. [Google Scholar] [CrossRef]
- Kirchhoff, M.D.; Kiverstein, J. Extended Consciousness and Predictive Processing: A Third-Wave View; Routledge: New York, NY, USA, 2019. [Google Scholar]
- Calvo, P.; Baluška, F.; Sims, A. “Feature detection” vs. “predictive coding” models of plant behavior. Front. Psychol. Cogn. Sci. 2016, 7, 1505. [Google Scholar] [CrossRef] [Green Version]
- Friston, K. Does predictive coding have a future? Nat. Neurosci. 2018, 21, 1019–1021. [Google Scholar] [CrossRef]
- Bowers, J.S.; Davis, C.J. Bayesian just-so stories in psychology and neuroscience. Psychol. Bull. 2012, 138, 389. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.L.; Chemero, T. The problem with brain GUTs: Conflation of different senses of “prediction” threatens metaphysical disaster. Behav. Brain Sci. 2013, 36, 204–205. [Google Scholar] [CrossRef]
- Raja, V.; Valluri, D.; Baggs, E.; Chemero, A.; Anderson, M.L. The Markov blanket trick: On the scope of the free energy principle and active inference. Phys. Life Rev. 2021, 39, 49–72. [Google Scholar] [CrossRef] [PubMed]
- Kogo, N.; Trengove, C. Is predictive coding theory articulated enough to be testable? Front. Comput. Neurosci. 2015, 9, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayo, D.G. Statistical Inference as Severe Testing; Cambridge University Press: New York, NY, USA, 2018. [Google Scholar]
- Colombo, M.; Elkin, L.; Hartmann, S. Being realist about Bayes, and the predictive processing theory of mind. Br. J. Philos. Sci. 2018, axy059. [Google Scholar] [CrossRef] [Green Version]
- Harkness, D.L.; Keshava, A. Moving from the what to the how and where—Bayesian models and predictive processing. In Philosophy and Predictive Processing (15); Metzinger, T., Wiese, W., Eds.; MIND Group: Frankfurt am Main, Germany, 2017. [Google Scholar] [CrossRef]
- Bernardo, J.M.; Smith, A.F.M. Bayesian Theory; John Wiley & Sons, Inc: New York, NY, USA, 2000. [Google Scholar]
- Easwaran, K. Bayesianism I: Introduction and arguments in favor. Philos. Compass 2011, 6, 312–320. [Google Scholar] [CrossRef]
- Easwaran, K. Bayesianism II: Applications and criticisms. Philos. Compass 2011, 6, 321–332. [Google Scholar] [CrossRef]
- Kruschke, J.K. Doing Bayesian Data Analysis: A Tutorial with R, JAGS, and Stan, 2nd ed.; Academic Press: San Diego, CA, USA, 2015. [Google Scholar]
- Kruschke, J.K.; Liddell, T.M. Bayesian data analysis for newcomers. Psychon. Bull. Rev. 2018, 25, 155–177. [Google Scholar] [CrossRef] [Green Version]
- Spiegelhalter, D.; Rice, K. Bayesian statistics. Scholarpedia 2009, 4, 5230. [Google Scholar] [CrossRef]
- Merckelbach, H.; van de Ven, V. Another White Christmas: Fantasy proneness and reports of ‘hallucinatory experiences’ in undergraduate students. J. Behav. Ther. Exp. Psychiatry 2001, 32, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Lam, L. Nonlinear Physics for Beginners: Fractals, Chaos, Solitons, Pattern Formation, Cellular Automata and Complex Systems; World Scientific: River Edge, NJ, USA, 1998. [Google Scholar]
- Ivancevic, V.G.; Ivancevic, T.T. Computational Mind: A Complex Dynamics Perspective; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Amon, M.J.; Holden, J.G. The mismatch of intrinsic fluctuations and the static assumptions of linear statistics. Rev. Philos. Psychol. 2021, 12, 149–173. [Google Scholar] [CrossRef]
- Klein, J.L. Statistical Visions in Time: A History of Time Series Analysis, 1662–1938; Cambridge University Press: New York, NY, USA, 1997. [Google Scholar]
- Gauss, C.F. Theoria Motus Corporum Coelestium in Sectionibus Conicis Solem Ambientium [Theory of the Motion of the Heavenly Bodies Moving about the Sun in Conic Sections]; Perthes et Besser: Hamburg, Germany, 1809; Volume 7. [Google Scholar]
- Fischer, H. A History of the Central Limit Theorem: From Classical to Modern Probability Theory; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Laplace, P.S. Théorie Analytique des Probabilités [Analytical Theory of Probabilities]; Courcier: Edmond, OK, USA, 1820. [Google Scholar]
- Cumming, G. Understanding the New Statistics: Effect Sizes, Confidence Intervals, and Meta-Analysis; Routledge: New York, NY, USA, 2013. [Google Scholar]
- Everitt, B.S. The Cambridge Dictionary of Statistics, 3rd ed.; Cambridge University Press: New York, NY, USA, 2006. [Google Scholar]
- Hoekstra, R.; Kiers, H.; Johnson, A. Are assumptions of well-known statistical techniques checked, and why (not)? Front. Psychol. Quant. Psychol. Meas. 2012, 3, 137. [Google Scholar] [CrossRef] [Green Version]
- Dienes, Z. Using Bayes to get the most out of non-significant results. Front. Psychol. Quant. Psychol. Meas. 2014, 5, 781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelman, A.; Stern, H.S.; Carlin, J.B.; Dunson, D.B.; Vehtari, A.; Rubin, D.B. Bayesian Data Analysis, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2009. [Google Scholar]
- Rao, R.P.; Ballard, D.H. Predictive coding in the visual cortex: A functional interpretation of some extra-classical receptive-field effects. Nat. Neurosci. 1999, 2, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Makin, J.G.; Dichter, B.K.; Sabes, P.N. Learning to estimate dynamical state with probabilistic population codes. PLoS Comput. Biol. 2015, 11, e1004554. [Google Scholar] [CrossRef] [PubMed]
- Farmer, J.D. A Rosetta stone for connectionism. Phys. D Nonlinear Phenom. 1990, 42, 153–187. [Google Scholar] [CrossRef] [Green Version]
- Hohwy, J. The Predictive Mind; Oxford University Press: New York, NY, USA, 2013. [Google Scholar]
- James, R.C. Life Magazine, 19 February 1965. p. 120. Available online: http://books.google.fr/books?id=KUEEAAAAMBAJ&lpg=PP1&hl=fr&pg=PA120#v=onepage&q&f=false (accessed on 21 December 2022).
- Gregory, R.L. The Intelligent Eye; Weidenfeld and Nicolson Ltd.: London, UK, 1970. [Google Scholar]
- Goldstein, M. Bayes linear analysis. Wiley StatsRef Stat. Ref. Online 2015, 1, 1–7. [Google Scholar] [CrossRef]
- Chen, C.C.; Kilner, J.M.; Friston, K.J.; Kiebel, S.J.; Jolly, R.K.; Ward, N.S. Nonlinear coupling in the human motor system. J. Neurosci. 2010, 30, 8393–8399. [Google Scholar] [CrossRef]
- Spratling, M.W. A review of predictive coding algorithms. Brain Cogn. 2017, 112, 92–97. [Google Scholar] [CrossRef] [Green Version]
- Friston, K.; Kiebel, S. Predictive coding under the free-energy principle. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Metropolis, N. The beginning of the Monte Carlo method. Los Alamos Sci. 1987, 1987, 125–130. [Google Scholar]
- Eliasmith, C.; Anderson, C.H. Neural Engineering: Computation, Representation, and Dynamics in Neurobiological Systems; The MIT Press: Cambridge, MA, USA, 2003. [Google Scholar]
- Aldrich, J.H.; Nelson, F.D. Linear Probability, Logit, and Probit Models; SAGE Publications, Inc.: Newbury Park, CA, USA, 1984. [Google Scholar]
- Ma, W.J.; Beck, J.M.; Latham, P.E.; Pouget, A. Bayesian inference with probabilistic population codes. Nat. Neurosci. 2006, 9, 1432–1438. [Google Scholar] [CrossRef]
- Mumford, J.A.; Ramsey, J.D. Bayesian networks for fMRI: A primer. NeuroImage 2014, 86, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, M.; Millidge, B.; Tschantz, A.; Buckley, C.L. How particular is the physics of the free energy principle? Phys. Life Rev. 2022, 40, 24–50. [Google Scholar] [CrossRef] [PubMed]
- Donders, F.C. On the speed of mental processes. Acta Psychol. 1969, 30, 412–431. [Google Scholar] [CrossRef] [PubMed]
- Huettel, S.A.; Song, A.W.; McCarthy, G. Functional Magnetic Resonance Imaging, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2009. [Google Scholar]
- Stam, C.J. Nonlinear dynamical analysis of EEG and MEG: Review of an emerging field. Clin. Neurophysiol. 2005, 116, 2266–2301. [Google Scholar] [CrossRef]
- Di Ieva, A. (Ed.) The Fractal Geometry of the Brain; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Shettigar, N.; Yang, C.L.; Tu, K.C.; Suh, C.S. On the biophysical complexity of brain dynamics: An outlook. Dynamics 2022, 2, 114–148. [Google Scholar] [CrossRef]
- Izhikevich, E.M. Dynamical Systems in Neuroscience: The Geometry of Excitability and Bursting; The MIT Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Beggs, J.M.; Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 2003, 23, 11167–11177. [Google Scholar] [CrossRef] [Green Version]
- Sporns, O. The human connectome: A complex network. Ann. N. Y. Acad. Sci. 2011, 1224, 109–125. [Google Scholar] [CrossRef]
- van Rooij, M.M.J.W.; Favela, L.H.; Malone, M.; Richardson, M.J. Modeling the dynamics of risky choice. Ecol. Psychol. 2013, 25, 293–303. [Google Scholar] [CrossRef]
- Riley, M.A.; Balasubramaniam, R.; Mitra, S.; Turvey, M.T. Visual influences on center of pressure dynamics in upright posture. Ecol. Psychol. 1998, 10, 65–91. [Google Scholar] [CrossRef]
- Tuller, B.; Case, P.; Ding, M.; Kelso, J.A. The nonlinear dynamics of speech categorization. J. Exp. Psychol. Hum. Percept. Perform. 1994, 20, 3–16. [Google Scholar] [CrossRef]
- Amon, M.J.; Pavlov, O.C.; Holden, J.G. Synchronization and fractal scaling as foundations for cognitive control. Cogn. Syst. Res. 2018, 50, 155–179. [Google Scholar] [CrossRef]
- Aks, D.J.; Zelinsky, G.J.; Sprott, J.C. Memory across eye-movements: 1/f dynamic in visual search. Nonlinear Dyn. Psychol. Life Sci. 2002, 6, 1–25. [Google Scholar] [CrossRef]
- Favela, L.H. Cognitive science as complexity science. Wiley Interdiscip. Rev. Cogn. Sci. 2020, 11, e1525. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.J.; Paxton, A.; Kuznetsov, N.A. Nonlinear Methods for Understanding Complex Dynamical Phenomena in Psychological Science. American Psychological Association—Psychological Science Agenda, February 2017. Available online: https://www.apa.org/science/about/psa/2017/02/dynamical-phenomena (accessed on 19 December 2022).
- Riley, M.A.; van Orden, G.C. (Eds.) Tutorials in Contemporary Nonlinear Methods for the Behavioral Sciences; National Science Foundation: Arlington, VA, USA, 2005. Available online: https://www.nsf.gov/pubs/2005/nsf05057/nmbs/nmbs.pdf (accessed on 19 December 2022).
- Favela, L.H. Dynamical systems theory in cognitive science and neuroscience. Philos. Compass 2020, 15, e12695. [Google Scholar] [CrossRef]
- Gilmore, R. Catastrophe Theory for Scientists and Engineers; Dover Publications, Inc.: New York, NY, USA, 1981. [Google Scholar]
- Isnard, C.A.; Zeeman, E.C. Some models from catastrophe theory in the social sciences. In The Use of Models in the Social Sciences; Collins, L., Ed.; Routledge: Chicago, IL, USA, 2019; pp. 44–100. [Google Scholar]
- Thom, R. Structural Stability and Morphogenesis: An Outline of a General Theory of Models; Benjamin: Reading, MA, USA, 1975. [Google Scholar]
- Zeeman, E.C. Catastrophe theory. Sci. Am. 1976, 234, 65–83. [Google Scholar] [CrossRef]
- Guastello, S.J.; Koopmans, M.; Pincus, D. (Eds.) Chaos and Complexity in Psychology: The Theory of Nonlinear Dynamical Systems; Cambridge University Press: New York, NY, USA, 2008. [Google Scholar]
- Haken, H. Synergetics: An Introduction, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 1983. [Google Scholar]
- van der Maas, H.L.J.; Kolstein, R.; van der Pligt, J. Sudden transitions in attitudes. Sociol. Methods Res. 2003, 32, 125–152. [Google Scholar] [CrossRef]
- Wang, W.; Chen, X.; Li, S.; Yan, H.; Yu, B.; Xu, Y. Cusp catastrophe modeling of suicide behaviors among people living with HIV in China. Nonlinear Dyn. Psychol. Life Sci. 2019, 23, 491–515. [Google Scholar]
- Yu, B.; Chen, X.; Stanton, B.; Chen, D.D.; Xu, Y.; Wang, Y. Quantum changes in self-efficacy and condom-use intention among youth: A chained cusp catastrophe model. J. Adolesc. 2018, 68, 187–197. [Google Scholar] [CrossRef]
- Chen, D.-G.; Chen, X. Cusp catastrophe regression and its application in public health and behavioral research. Environ. Res. Public Health 2017, 14, 1220. [Google Scholar] [CrossRef] [Green Version]
- Grasman, R.; van der Maas, H.L.; Wagenmakers, E.J. Fitting the cusp catastrophe in R: A cusp package primer. J. Stat. Softw. 2009, 32, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Batterman, R.W. Multiple realizability and universality. Br. J. Philos. Sci. 2000, 51, 115–145. [Google Scholar] [CrossRef]
- Thouless, D. Condensed matter physics in less than three dimensions. In The New Physics; Davies, P., Ed.; Cambridge University Press: New York, NY, USA, 1993; pp. 209–235. [Google Scholar]
- Jensen, H.J. Self-Organized Criticality: Emergent Complex Behavior in Physical and Biological Systems; Cambridge University Press: New York, NY, USA, 1998. [Google Scholar]
- Beggs, J.M. The criticality hypothesis: How local cortical networks might optimize information processing. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2008, 366, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Cocchi, L.; Gollo, L.L.; Zalesky, A.; Breakspear, M. Criticality in the brain: A synthesis of neurobiology, models and cognition. Prog. Neurobiol. 2017, 158, 132–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesse, J.; Gross, T. Self-organized criticality as a fundamental property of neural systems. Front. Syst. Neurosci. 2014, 8, 166. [Google Scholar] [CrossRef] [Green Version]
- Plenz, D.; Niebur, E. (Eds.) Criticality in Neural Systems; Wiley-VCH Verlag GmbH & Co.: Weinheim, Germany, 2014. [Google Scholar]
- Bak, P.; Tang, C.; Wiesenfeld, K. Self-organized criticality. Phys. Rev. A 1988, 38, 364–374. [Google Scholar] [CrossRef]
- Ramos, R.T.; Sassi, R.B.; Piqueira, J.R.C. Self-organized criticality and the predictability of human behavior. New Ideas Psychol. 2011, 29, 38–48. [Google Scholar] [CrossRef]
- Noori, H.R. Hysteresis Phenomena in Biology; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Schwitzgebel, E. The unreliability of naive introspection. Philos. Rev. 2008, 117, 245–273. [Google Scholar] [CrossRef] [Green Version]
- Sundareswara, R.; Schrater, P.R. Perceptual multistability predicted by search model for Bayesian decisions. J. Vis. 2008, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Sato, F.; Shiomoto, R.; Nakauchi, S.; Minami, T. Backward and forward neck tilt affects perceptual bias when interpreting ambiguous figures. Sci. Rep. 2022, 12, 10985. [Google Scholar] [CrossRef]
- Ta’eed, L.K.; Ta’eed, O.; Wright, J.E. Determinants involved in the perception of the Necker cube: An application of catastrophe theory. Behav. Sci. 1988, 33, 97–115. [Google Scholar] [CrossRef]
- Haken, H. Synergetics of brain function. Int. J. Psychophysiol. 2006, 60, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Whitney, D. Serial dependence in visual perception. Nat. Neurosci. 2014, 17, 738–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Favela, L.H.; Amon, M.J. Enhancing Bayesian Approaches in the Cognitive and Neural Sciences via Complex Dynamical Systems Theory. Dynamics 2023, 3, 115-136. https://doi.org/10.3390/dynamics3010008
Favela LH, Amon MJ. Enhancing Bayesian Approaches in the Cognitive and Neural Sciences via Complex Dynamical Systems Theory. Dynamics. 2023; 3(1):115-136. https://doi.org/10.3390/dynamics3010008
Chicago/Turabian StyleFavela, Luis H., and Mary Jean Amon. 2023. "Enhancing Bayesian Approaches in the Cognitive and Neural Sciences via Complex Dynamical Systems Theory" Dynamics 3, no. 1: 115-136. https://doi.org/10.3390/dynamics3010008
APA StyleFavela, L. H., & Amon, M. J. (2023). Enhancing Bayesian Approaches in the Cognitive and Neural Sciences via Complex Dynamical Systems Theory. Dynamics, 3(1), 115-136. https://doi.org/10.3390/dynamics3010008