

Biomass and β-Glucosidase Production by the Cyanobacterium Pseudanabaena sp. under Heterotrophic Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sampling
2.2. Isolation
2.3. DNA Extraction, PCR Amplification, Cloning, and Sequencing
2.4. Nucleotide Sequences and Accession Numbers
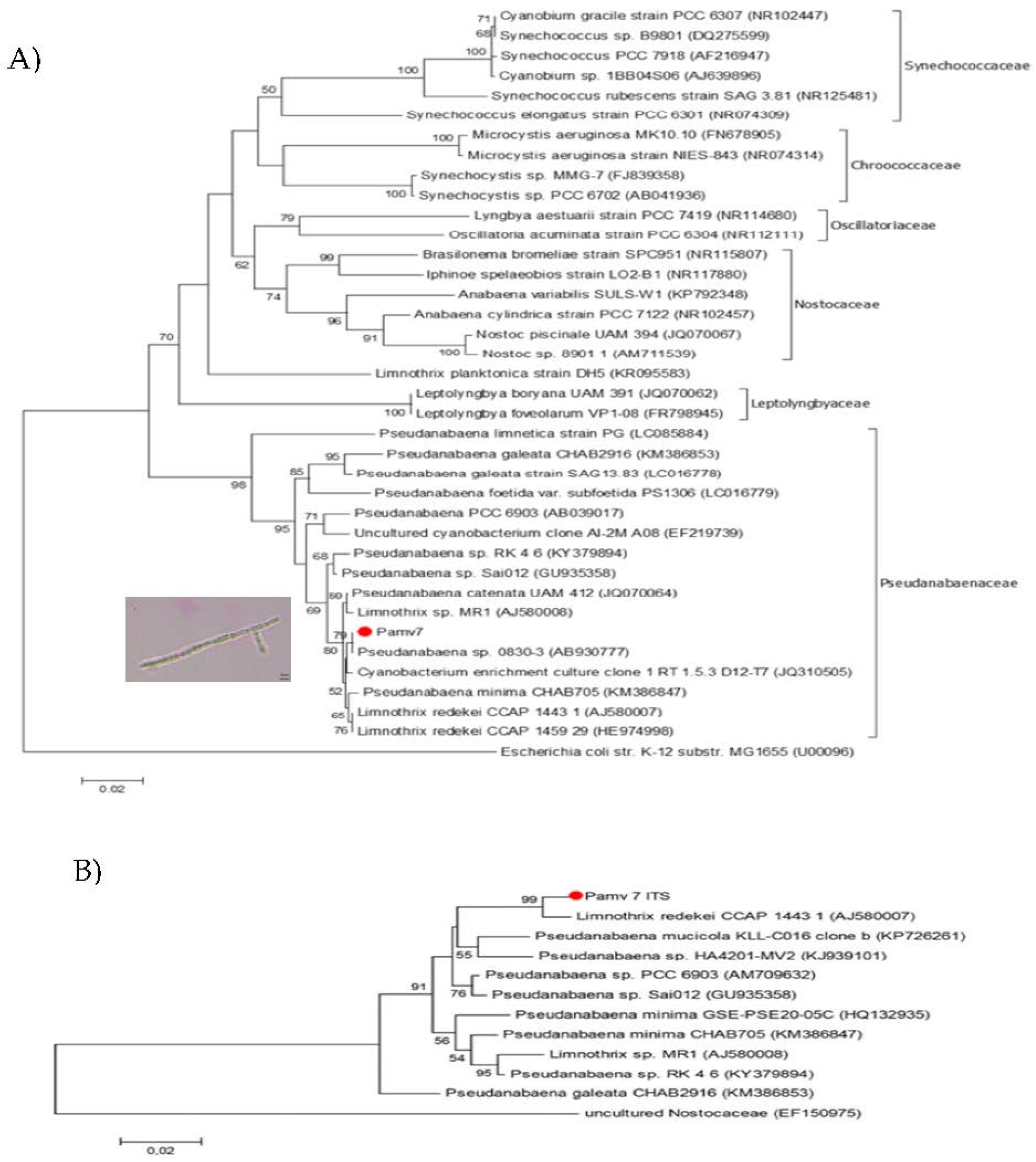
2.5. Phylogenetic Characterization
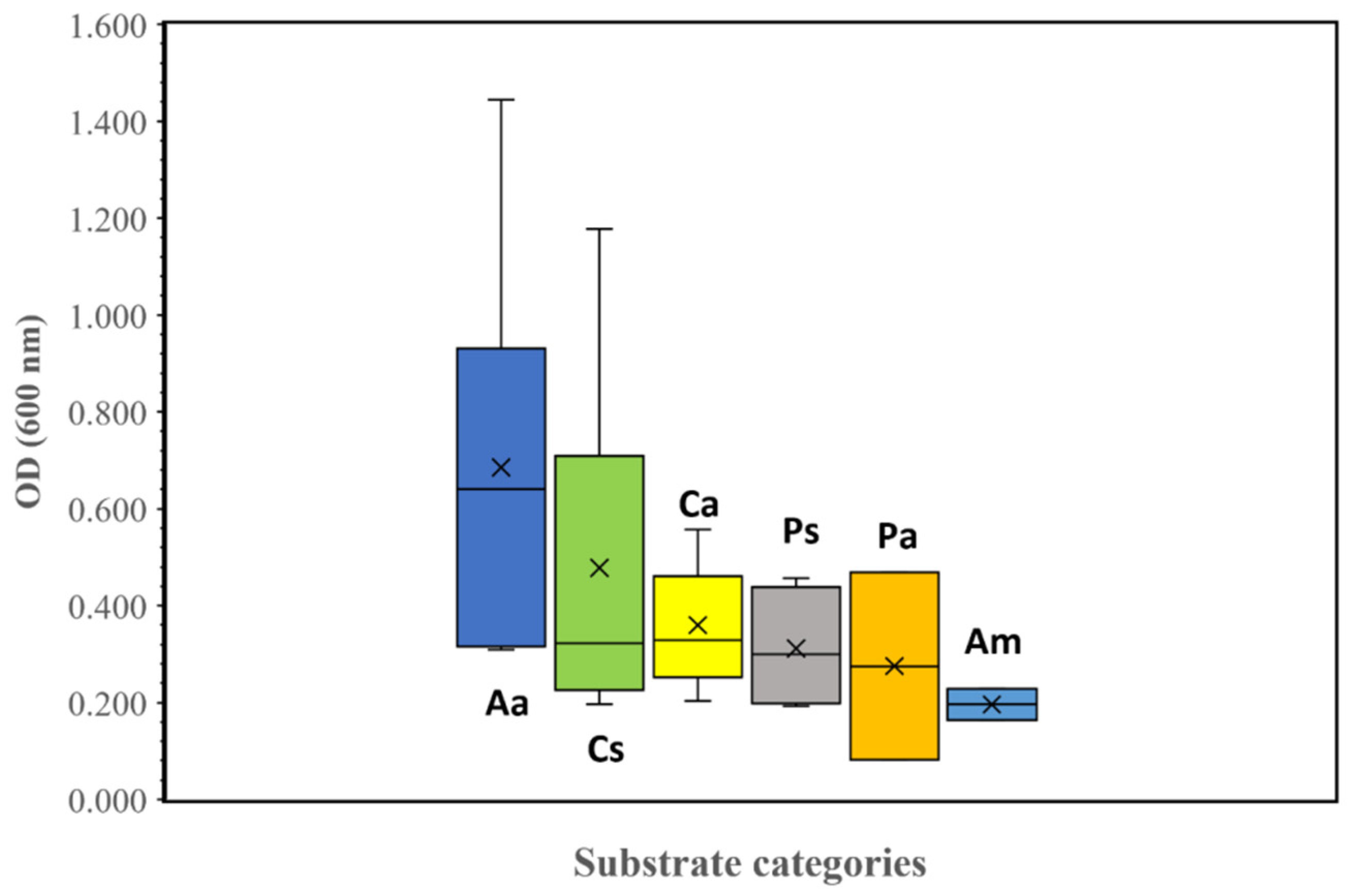
2.6. Screening of Substrates for Heterotrophic Growth of Pseudanabaena sp. Cyanobacterium (BIOLOG)
2.7. Culture Growth Conditions of Pseudanabaena sp. Cells and β-Glucosidase Production
2.8. β-Glucosidase Assay
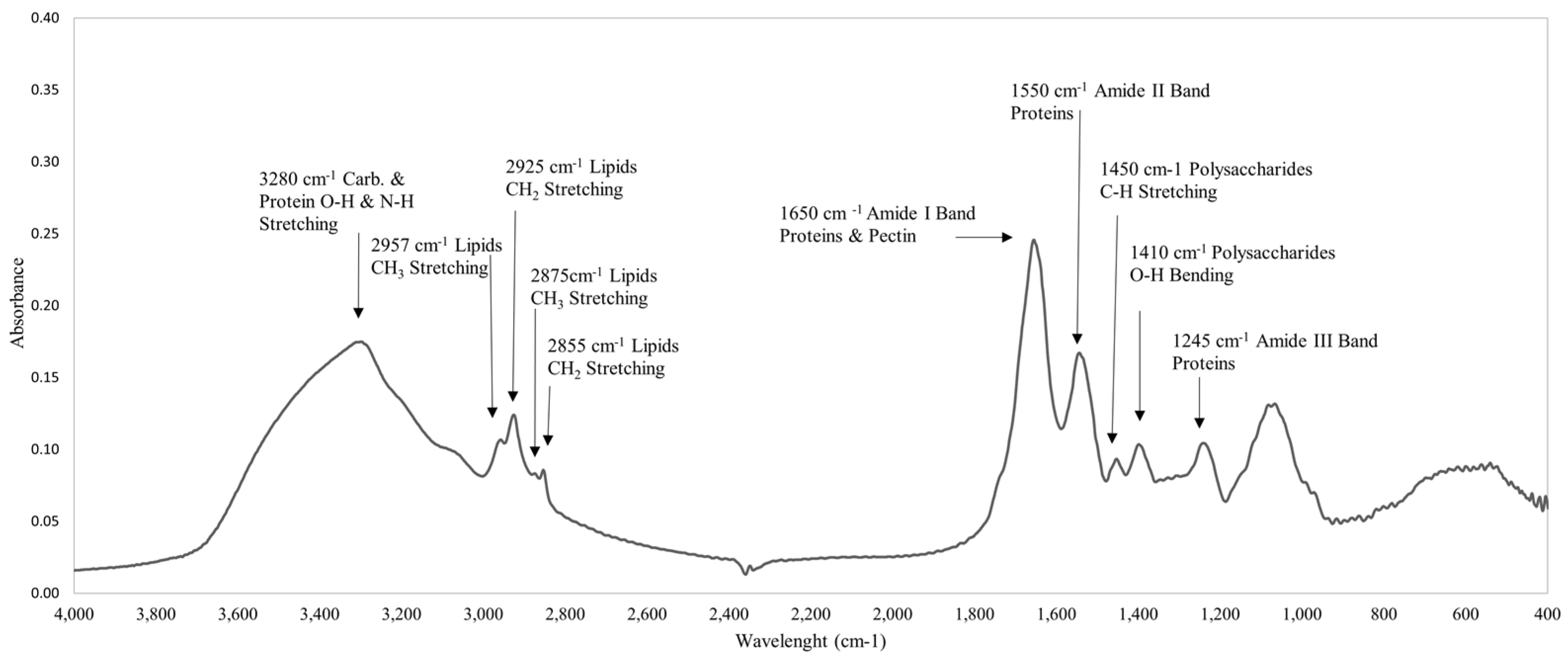
2.9. FT-IR Spectrum
2.10. Protein and Lipid Determination
2.11. Statistical Analysis
3. Results and Discussion
3.1. Isolation and Phylogenetic Characterization
3.2. Heterotrophic Characterization by Biolog-ECO Plate
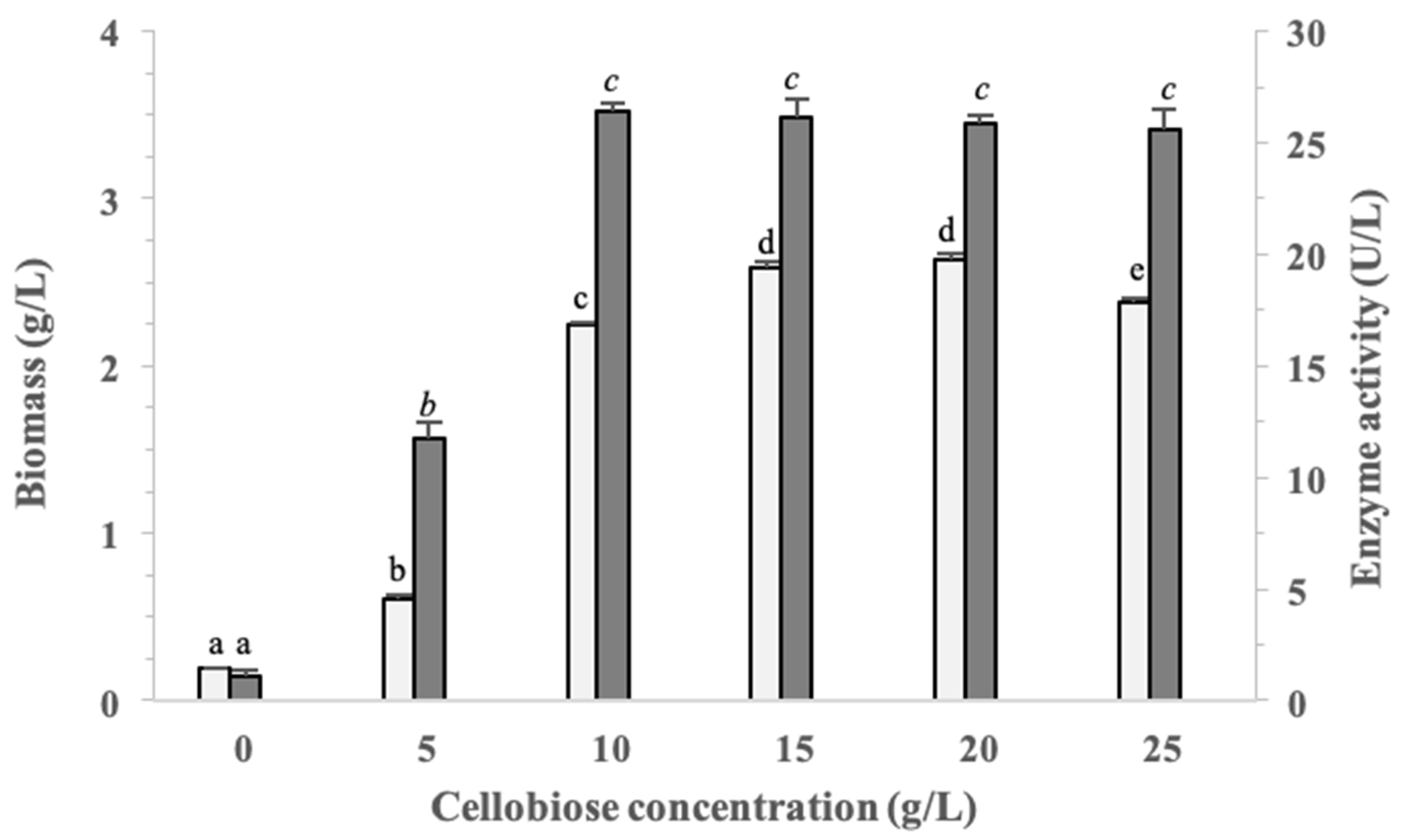
3.3. Optimization of Cellobiose Concentration on Biomass and β-Glucosidase Production in Pseudanabaena sp. Cells
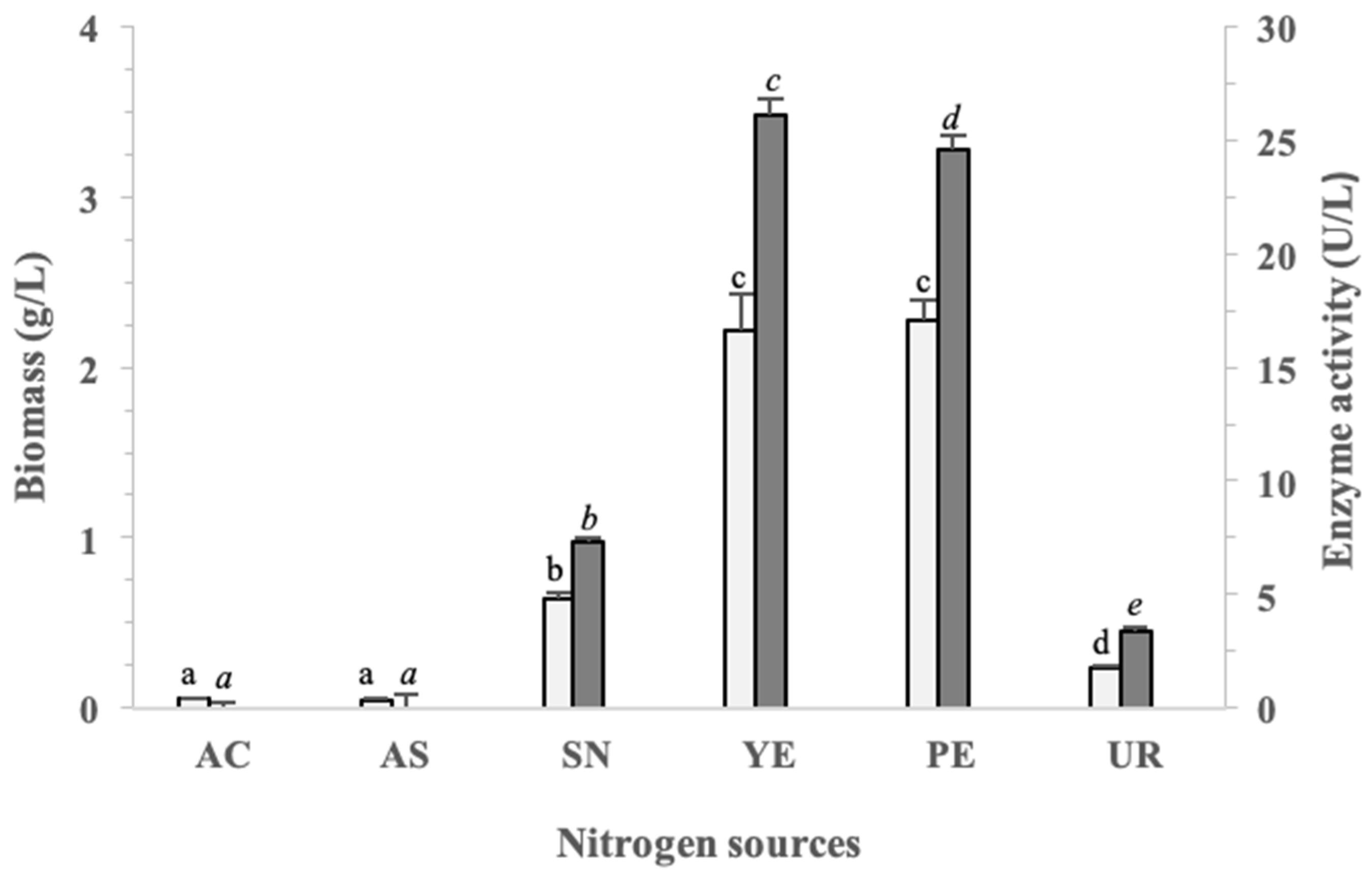
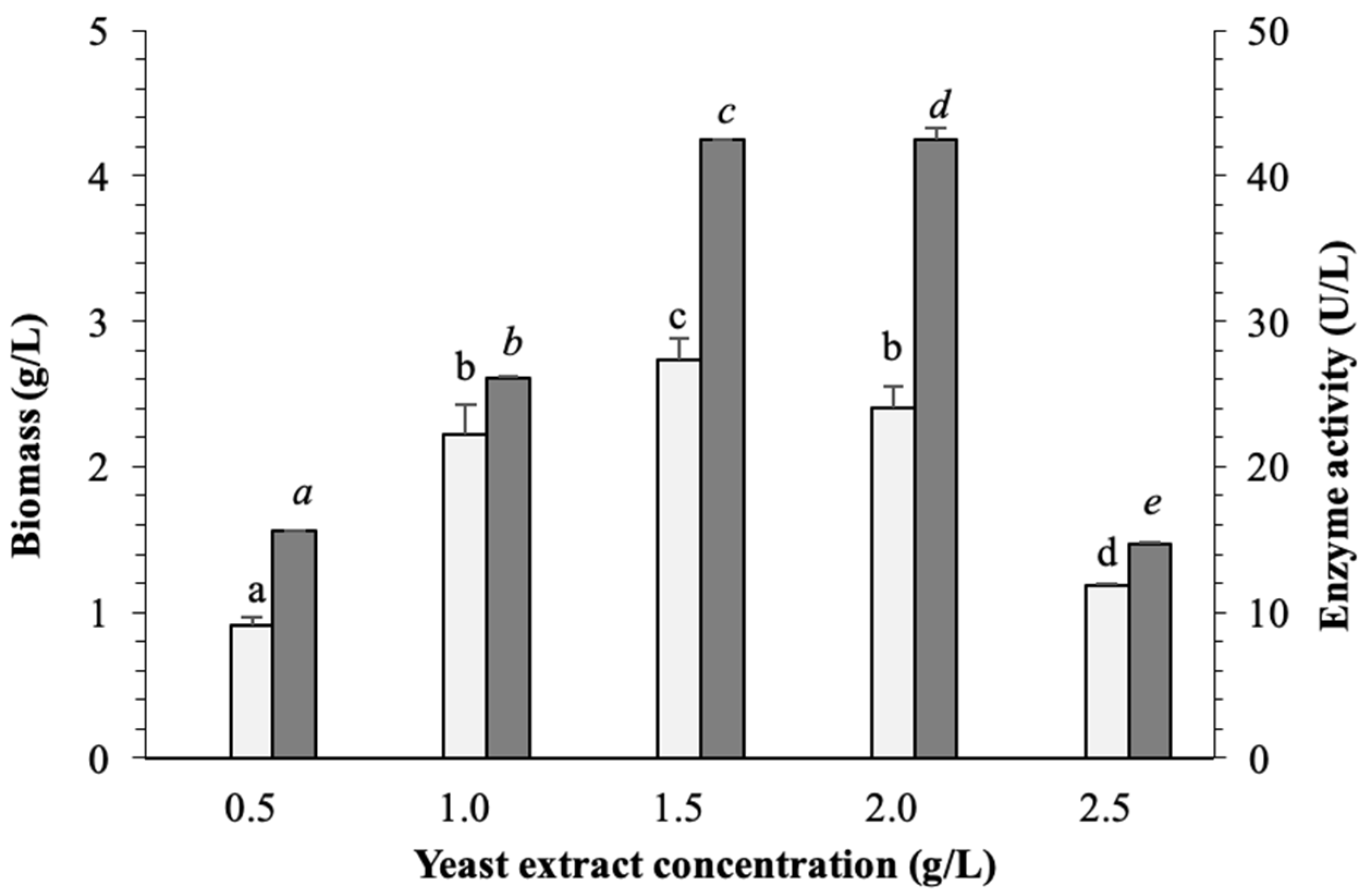
3.4. Optimization of Nitrogen Source on Biomass and β-Glucosidase Production in Pseudanabaena sp. Cells
3.5. Optimization of Nitrogen Concentration on Biomass and β-Glucosidase Production in Pseudanabaena sp. Cells
3.6. Optimization of Temperature on Biomass and β-Glucosidase Production in Pseudanabaena sp. Cells
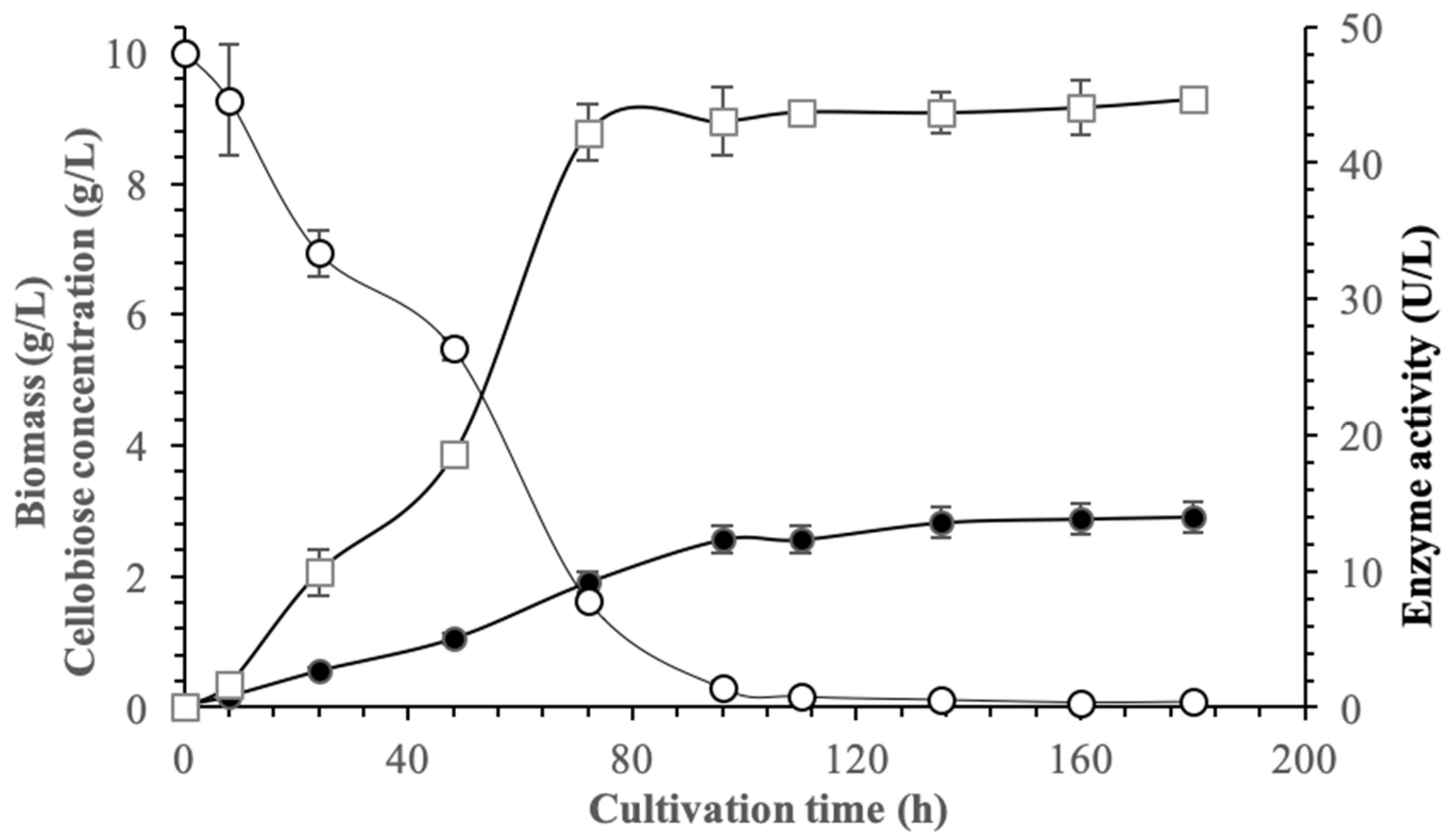
3.7. Growth of Pseudanabaena sp. under Optimum Conditions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmed, A.; Batool, K.; Bibi, A. Microbial β-Glucosidase: Sources, Production and Applications. J. Appl. Environ. Microbiol. 2017, 5, 31–46. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Meng, T.; Li, Z.; Liu, P.; Wang, Y. Characterization of a beta-glucosidase from Bacillus licheniformis and its effect on bioflocculant degradation. AMB Express. 2017, 7, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singhania, R.R.; Sukumaran, R.K.; Larroche, C.; Pandey, A. Role and significance of beta-glucosidases in the hydrolysis of cellulose for bioethanol production. Bioresour. Technol. 2013, 127, 500–507. [Google Scholar] [CrossRef]
- Tiwari, P.; Misra, B.N.; Sangwan, N.S. Β-Glucosidases from the fungus Trichoderma: An efficient cellulase machinery in biotechnological applications. BioMed. Res. Int. 2013, 2013, 203735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.; Aslam, M.; Ashraf, M.; ul-Hassan Nasim, F.; Batool, K.; Bibi, A. Microbial β-Glucosidases: Screening, Characterization, Cloning and Applications. J. Appl. Environ. Microbiol. 2017, 5, 57–73. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Lee, Y.-S.; Fang, S.-J.; Park, D.-J.; Choi, Y.-L. Hydrolysis of isoflavone glycoside by immobilization of β-glucosidase on a chitosan-carbon in two-phase system. Int. J. Biol. Macromol. 2013, 61, 465–470. [Google Scholar] [CrossRef]
- GV Sharada, R.; Venkateswar, S.; AnandRao, M. Applications of cellulases-review. Int. J. Pharm. Chem. Biol. Sci. 2014, 4, 424–437. [Google Scholar]
- Sørensen, A.; Lübeck, M.; Lübeck, P.S.; Ahring, B.K. Fungal Beta-Glucosidases: A Bottleneck in Industrial Use of Lignocellulosic Materials. Biomolecules 2013, 3, 612–631. [Google Scholar] [CrossRef] [Green Version]
- Seeberger, P.H.; Werz, D.B. Synthesis and medical applications of oligosaccharides. Nature 2007, 446, 1046–1051. [Google Scholar] [CrossRef]
- Lambertz, C.; Garvey, M.; Klinger, J.; Heesel, D.; Klose, H. Challenges and advances in the heterologous expression of cellulolytic enzymes: A review. Biotechnol. Biofuels 2014, 7, 135. [Google Scholar] [CrossRef] [Green Version]
- Nigam, P.S. Microbial enzymes with special characteristics for biotechnological applications. Biomolecules 2013, 3, 597–611. [Google Scholar] [CrossRef]
- Mogharabia, M. Are Algae the Future Source of Enzymes? Trends Pept. Protein Sci. 2016, 1, 1–6. [Google Scholar]
- Economou, C.N.; Moustaka-Gouni, M.; Kehayias, G.; Aggelis, G.; Vayenas, G.V. Lipid production by the filamentous cyanobacterium Limnothrix sp. growing in synthetic wastewater in suspended- and attached-growth photobioreactor systems. Ann. Microbiol. 2015, 65, 1941–1948. [Google Scholar] [CrossRef]
- Meireles dos Santos, A.; Vieira, K.R.; Basso Sartori, R.; Meireles dos Santos, A.; Queiroz, M.I.; Queiroz Zepka, L.; Jacob-Lopes, E. Heterotrophic Cultivation of Cyanobacteria: Study of Effect of Exogenous Sources of Organic Carbon, Absolute Amount of Nutrients, and Stirring Speed on Biomass and Lipid Productivity. Front. Bioeng. Biotechnol. 2017, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.; Singh, D.; Tyagi, M.B.; Kumar, A. Chapter 16—Cyanobacteria: Applications in Biotechnology. In Cyanobacteria from Basic Science to Applications; Academic Press: Cambridge, MA, USA, 2019; pp. 327–346. [Google Scholar]
- Khalifa, S.; Shedid, E.S.; Saied, E.M.; Jassbi, A.R.; Jamebozorgi, F.H.; Rateb, M.E.; Du, M.; Abdel-Daim, M.M.; Kai, G.Y.; Al-Hammady, M.; et al. Cyanobacteria-From the Oceans to the Potential Biotechnological and Biomedical Applications. Mar. Drugs 2021, 19, 241. [Google Scholar] [CrossRef]
- Romanowska-Duda, Z.; Szufa, S.; Grzesik, M.; Piotrowski, K.; Janas, R. The Promotive Effect of Cyanobacteria and Chlorella sp. Foliar Biofertilization on Growth and Metabolic Activities of Willow (Salix viminalis L.) Plants as Feedstock Production, Solid Biofuel and Biochar as C Carrier for Fertilizers via Torrefaction Process. Energies 2021, 14, 5262. [Google Scholar]
- Jacob-Lopes, E.; Santos, A.B.; Severo, I.A.; Manoreze, M.M.; Zepka, L.Q. Dual production of bioenergy in heterotrophic cultures of cyanobacteria: Process performance, carbon balance, biofuel quality and sustainability metrics. Biomass Bioenergy 2020, 142, 105756. [Google Scholar] [CrossRef]
- Roman, K.; Barwicki, J.; Rzodkiewicz, W.; Dawidowski, M. Evaluation of Mechanical and Energetic Properties of the Forest Residues Shredded Chips during Briquetting Process. Energies 2021, 14, 3270. [Google Scholar] [CrossRef]
- Rodjaroen, S.N.; Mahakhant, A.; Miyamoto, K. High biomass production and starch accumulation in native green algal strains and cyanobacterial strains of Thailand. Kasetsart J. (Nat. Sci.) 2007, 41, 570–575. [Google Scholar]
- Singh, G. Cyanobacteria as a novel source of reneable energy. Plant Arch. 2017, 17, 1790–1794. [Google Scholar]
- Markou, G.; Georgakakis, D. Cultivation of filamentous cyanobacteria (blue-green algae) in agro-industrial wastes and wastewaters: A review. Appl. Energy 2011, 88, 3389–3401. [Google Scholar] [CrossRef]
- Kroumov, A.D.; Scheufele, F.B.; Zaharieva, M.M.; Zheleva-Dimitrova, D.; Najdenski, H. Optimal Biomass Production by Cyanobacteria, Mathematical Evaluation, and Improvements in the Light of Biorefinery Concept. In Ecophysiology and Biochemistry of Cyanobacteria; Rastogi, R.P., Ed.; Springer: Singapore, 2021; pp. 401–429. [Google Scholar]
- Karageorgou, D.; Zygouri, P.; Tsakiridis, T.; Hammami, M.A.; Chalmpes, N.; Subrati, M.; Sainis, I.; Spyrou, K.; Katapodis, P.; Gournis, D.; et al. Green Synthesis and Characterization of Silver Nanoparticles with High Antibacterial Activity Using Cell Extracts of Cyanobacterium Pseudanabaena/Limnothrix sp. Nanomaterials 2022, 12, 2296. [Google Scholar] [CrossRef] [PubMed]
- Karageorgou, D.; Patel, A.; Rova, U.; Christakopoulos, P.; Katapodis, P.; Matsakas, L. Heterotrophic Cultivation of the CynobacteriumPseudanabaena sp. on Forest Biomass Hydrolysates towards Sustainable Biodiesel Production. Microorganisms 2022, 10, 1756. [Google Scholar] [CrossRef] [PubMed]
- Cegłowska, M.; Szubert, K.; Grygier, B.; Lenart, M.; Plewka, J.; Milewska, A.; Lis, K.; Szczepański, A.; Chykunova, Y.; Barreto-Duran, E.; et al. Pseudanabaenagaleata CCNP1313-Biological Activity and Peptides Production. Toxins 2022, 14, 330. [Google Scholar] [CrossRef] [PubMed]
- Cano-Europa, E.; Ortiz-Butrón, R.; Gallardo-Casas, C.A.; Blas-Valdivia, V.; Pineda-Reynoso, M.; Olvera-Ramírez, R.; Franco-Colin, M. Phycobiliproteins from Pseudanabaena tenuis rich in c-phycoerythrin protect against HgCl2-caused oxidative stress and cellular damage in the kidney. J. Appl. Phycol. 2010, 22, 495–501. [Google Scholar] [CrossRef]
- Vareli, K.; Pilidis, G.; Mavrogiorgou, M.C.; Briasoulis, E.; Sainis, I. Molecular characterization of cyanobacterial diversity and yearly fluctuations of Microcystin loads in a suburban Mediterranean Lake (Lake Pamvotis, Greece). J. Environ. Monit. 2009, 11, 1506–1512. [Google Scholar] [CrossRef]
- Touka, A.; Vareli, K.; Igglezou, M.; Monokrousos, N.; Alivertis, D.; Halley, J.M.; Hadjikakou, S.; Frillingos, S.; Sainis, I. Ancient European Lakes: Reservoirs of Hidden Microbial Diversity? The Case of Lake Pamvotis (NW Greece). Open J. Ecol. 2018, 8, 537–578. [Google Scholar] [CrossRef] [Green Version]
- Edem, P.A.; Schmidt, T.M.; Blakemore, T.R.; Pace, N.R. Phylogenetic Analysis of Aquaspirillummagnetotacticum Using Polymerase Chain Reaction-Amplified 16s rRNA-Specific DNA. Int. J. Syst. Bacteriol. 1991, 41, 324–325. [Google Scholar] [CrossRef]
- Muyzer, G.; De Waal, E.C.; Uitterlinden, A. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Janse, I.; Meima, M.; Kardinaal, W.E.A.; Zwart, G. High-resolution differentiation of Cyanobacteria by using rRNA-internal transcribed spacer denaturing gradient gel electrophoresis. Appl. Environ. Microbiol. 2003, 69, 6634–6643. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Tian-Yuan, Z.; Yin-Hu, W.; Lin-Lan, Z.; Xiao-Xiong, W.; Hong-Ying, H. Screening heterotrophic microalgal strains by using the Biolog method for biofuel production from organic wastewater. Algal Res. 2014, 6, 175–179. [Google Scholar] [CrossRef]
- Choi, K.-H.; Dobbs, F.C. Comparison of two kinds of Biolog microplates (GN and ECO) in their ability to distinguish among aquatic microbial communities. J. Microbiol. Methods 1999, 36, 203–213. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Komárek, J.; Kaštovský, J.; Mareš, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera), using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Sutherland, D.L.; Burke, J.; Ralph, P.J. High-throughput screening for heterotrophic growth in microalgae using the Biolog Plate assay. New Biotechnol. 2021, 65, 61–68. [Google Scholar] [CrossRef]
- Francisco, E.C.; Franco, T.T.; Wagner, R.; Jacob-Lopes, E. Assessment of different carbohydrates as exogenous carbon source in cultivation of cyanobacteria. Bioprocess Biosyst. Eng. 2014, 37, 1497–1505. [Google Scholar] [CrossRef]
- Lee, T.-C.; Xiong, W.; Paddock, T.; Carrieri, D.; Chang, I.-F.; Chiu, H.-F.; Ungerer, J.; Juo, S.-H.H.; Maness, P.-C.; Yu, J. Engineered xylose utilization enhances bio-products productivity in the cyanobacterium Synechocystis sp. PCC 6803. Metab. Eng. 2015, 30, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.; Matsakas, L.; Rova, U.; Christakopoulos, P. Heterotrophic cultivation of Auxenochlorellaprotothecoides using forest biomass as a feedstock for sustainable biodiesel production. Biotechnol. Biofuels 2018, 11, 169. [Google Scholar] [CrossRef]
- Miao, X.; Li, P.; Li, R.; Zhong, J. In situ biodiesel production from fast-growing and high oil content Chlorella pyrenoidosa in rice straw hydrolysate. J. Biomed. Biotechnol. 2011, 2011, 141207. [Google Scholar]
- Gao, C.; Zhai, Y.; Ding, Y.; Wu, Q. Application of sweet sorghum for biodiesel production by heterotrophic microalga Chlorella protothecoides. Appl. Energy 2010, 87, 756–761. [Google Scholar] [CrossRef]
- Gilpin, L.C.; Davidson, K.; Roberts, E. The influence of changes in nitrogen: Silicon ratios on diatom growth dynamics. J. Sea Res. 2004, 51, 21–35. [Google Scholar] [CrossRef]
- Kim, G.; Mujtaba, G.; Lee, K. Effects of nitrogen sources on cell growth and biochemical composition of marine chlorophyte Tetraselmis sp. for lipid production. Algae 2016, 31, 257–266. [Google Scholar]
- Converti, A.; Scapazzoni, S.; Lodi, A.; Carvalho, J.C.M. Ammonium and urea removal by Spiru;ina platensis. J. Ind. Microbiol. 2006, 33, 8–16. [Google Scholar]
- Vieira Costa, J.A.; Cozza, K.L.; Oliviera, L.; Magagnin, G. Different nitrogen sources and growth responses of Spirulina platensis in microenvironments. World J. Microbiol. Biotechnol. 2001, 17, 439–442. [Google Scholar]
- Johnson, T.J.; Jahandideh, A.; Isaac, I.C.; Baldwin, E.L.; Muthukumarappan, K.; Zjou, R.; Gibbons, W.R. Determining the optimal nitrogen source for large-scale cultivation of filamentous cyanobacteria. J. Appl. Phycol. 2016, 29, 1–3. [Google Scholar] [CrossRef]
- Norici, A.; Dalsass, A.; Giordano, M. Role of phosphoenolpyruvate carboxylase in anaplerosis in the green microalga Dunaliellasalina cultured under different nitrogen regimes. Physiol Plant. 2002, 116, 186–191. [Google Scholar]
- Wan, M.X.; Wang, R.M.; Xia, J.L.; Rosenberg, J.N.; Nie, Z.Y.; Kobayashi, N.; Oyler, G.A.; Betenbaugh, M.J. Physiological evaluation of a new Chlorella sorokiniana isolate for its biomass production and lipid accumulation in photoautotrophic and heterotrophic cultures. Biotechnol. Bioeng. 2012, 109, 1958–1964. [Google Scholar] [CrossRef]
- Patel, A.; Pruthi, V.; Pruthi, P.A. Synchronized nutrient stress conditions trigger the diversion of CDP-DG pathway of phospholipids synthesis towards de novo TAG synthesis in oleaginous yeast escalating biodiesel production. Energy 2017, 139, 962–974. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.P.; Singh, P. Effect of temperature and light on the growth of algae species: A review. Renew. Sust. Energ. Rev. 2015, 50, 431–444. [Google Scholar] [CrossRef]
- Gris, B.; Sforza, E.; Morosinotto, T.; Bertucco, A.; La Rocca, N. Influence of light and temperature on growth and high-value molecules productivity from Cyanobacterium aponinum. J. Appl. Phycol. 2017, 29, 1781–1790. [Google Scholar] [CrossRef]
- Kagalou, I.; Economidis, G.; Leonardos, I.; Papaloukas, C. Assessment of a Mediterranean shallow lentic ecosystem (Lake Pamvotis, Greece) using benthic community diversity: Response to environmental parameters. Limnologica 2006, 36, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Garcia, N.F.L.; da Silva Santos, F.R.; Goncalves, F.A.; de Paz, M.F.; Fonseca, G.G.; Leite, R.S.R. Production of β-glucosidase on solid-state fermentation by Lichtheimiaramosa in agroindustrial residues: Characterization and catalytic properties of the enzymatic extract. Electron. J. Biotechnol. 2015, 18, 314–319. [Google Scholar] [CrossRef] [Green Version]
- Olajuyigbe, F.M.; Nlekerem, C.M.; Ogunyewo, O.A. Production and Characterization of Highly Thermostable β-Glucosidase during the Biodegradation of Methyl Cellulose by Fusarium oxysporum. Biochem. Res. Int. 2016, 2016, 3978124. [Google Scholar] [CrossRef] [Green Version]
- Özer, T.; Yalçın, D.; Erkaya, I.A.; Udoh, A.U. Identification and Characterization of Some Species of Cyanobacteria, Chlorophyta and BacillariophytaUsing Fourier- Transform Infrared (FTIR) Spectroscopy. Int. J. Pharm. Biol. Sci. 2016, 11, 20–27. [Google Scholar]
- Bataller, B.G.; Capareda, S.C. A rapid and non-destructive method for quantifying biomolecules in Spirulina platensis via Fourier transform infrared—Attenuated total reflectance spectroscopy. Algal Res. 2018, 32, 341–352. [Google Scholar] [CrossRef]
- Meng, Y.; Yao, C.; Xue, S.; Yang, H. Application of Fourier transform infrared (FT-IR) spectroscopy in determination of microalgal compositions. Bioresour. Technol. 2014, 151, 347–354. [Google Scholar]
- Oufdou, K.; Mezrioui, N.; Oudra, B.; Loudiki, M.; Barakate, M.; Sbiyyaa, B. Bioactive compounds from Pseudanabaena species (Cyanobacteria). Microbios 2001, 106, 21–29. [Google Scholar]
- Rajeshwari, K.R.; Rajashekhar, M. Biochemical Composition of Seven Species of Cyanobacteria Isolated from Different Aquatic Habitats of Western Ghats, Southern India. Braz. Arch. Biol. Technol. 2011, 54, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Christaki, E.; Florou-Paneri, P.; Bonos, E. Microalgae: A novel ingredient in nutrition. Int. J. Food Sci. Nutr. 2011, 62, 794–799. [Google Scholar] [CrossRef]
- Kim, S.-K.; Kang, K.-H. Medicinal Effects of Peptides from Marine Microalgae. Adv. Food Nutr. 2011, 64, 313–323. [Google Scholar]
- Siqueira, S.F.; Queiroz, M.I.; Zepka, L.Q.; Jacob-Lopes, E. Introductory Chapter: Microalgae Biotechnology—A Brief Introduction. In Microalgal Biotechnology; IntechOpen: London, UK, 2018. [Google Scholar]
- Mohamed, M.; Wei, L.Z.; Ariff, A.B. Heterotrophic cultivation of microalgae for production of biodiesel. Recent Pat. Biotechnol. 2011, 5, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Mikes, F.; Matsakas, L. An Overview of Current Pretreatment Methods Used to Improve Lipid Extraction from Oleaginous Micro-Organisms. Molecules 2018, 23, 1562. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pamv7 | Species GenBank | % Overlapping | Identity | Accession (15 June 2019) |
|---|---|---|---|---|
| 16S rDNA partial (471 bp) | Pseudanabaena sp. 0830-3 | 100 | 99.3% | AB936777 |
| ITS full length (703 bp) | Pseudanabaena sp. RK_4_6 | 99 | 91.9% | KY379894 |
| ITS full length (703 bp) | LimnothrixredekeiCCAP 1443/1 | 99 | 88% | AJ580007 |
| Substrate | Specific Growth Rate | Substrate | Specific Growth Rate |
|---|---|---|---|
| Carbohydrates | μ (d−1) | Carboxylicacids | μ (d−1) |
| β-Methyl-D-glucoside | 0.17 ± 0.01 | Pyruvicacidmethylester | 0.50 ± 0.01 |
| D-Galactonicacid γ-lactone | 0.29 ± 0.01 | D-Galacturonicacid | 0.42 ± 0.01 |
| D-Xylose | 0.42 ± 0.01 | γ-Hydroxybutyricacid | 0.15 ± 0.01 |
| i-Erythritol | 0.15 ± 0.01 | D-Glucosaminic acid | 0.39 ± 0.01 |
| D-Mannitol | 0.52 ± 0.01 | Itaconic acid | 0.26 ± 0.01 |
| N-Acetyl-D-glycosamine | 0.40 ± 0.01 | α-KetobutyricAcid | 0.42 ± 0.01 |
| D-Cellobiose | 0.60 ± 0.01 | D-Malic acid | 0.28 ± 0.01 |
| Glucose-1-phosphate | 0.16 ± 0.01 | ||
| α-D-lactose | 0.19 ± 0.01 | AminoAcids | μ (d−1) |
| d.l-A-Glycerol phosphate | 0.32 ± 0.01 | L-Arginine | 0.43 ± 0.01 |
| L-Asparagine | 0.70 ± 0.01 | ||
| Amines | μ (d−1) | L-Phenylalanine | 0.28 ± 0.01 |
| Phenylethylamine | 0.08 ± 0.01 | L-Serine | 0.48 ± 0.01 |
| Putrescine | 0.18 ± 0.01 | L-Threonine | 0.28 ± 0.01 |
| Polymers | μ (d−1) | Phenoliccompounds | μ (d−1) |
| Tween 40 | 0.38 ± 0.01 | 2-Hydroxy benzoic acid | 0.00 ± 0.01 |
| Tween 80 | 0.37 ± 0.01 | 4-Hydroxy benzoicacid | 0.37 ± 0.01 |
| α-Cyclodextrin | 0.16 ± 0.01 | ||
| Glycogen | 0.14 ± 0.01 |
| Cultivation Temperature (°C) | Production by Pseudanabaena sp. | |
|---|---|---|
| Biomass (g/L) | β-Xylosidase (U/L) | |
| 15 | 1.64 ± 0.12 a | 30.27 ± 0.05 a |
| 18 | 1.63 ± 0.15 a | 30.01 ± 0.46 a |
| 20 | 2.00 ± 0.02 b | 37.66 ± 0.41 b |
| 23 | 2.80 ± 0.19 c | 44.69 ± 0.75 c |
| 25 | 2.76 ± 0.09 c | 43.05 ± 0.55 cd |
| 28 | 2.46 ± 0.11 d | 42.01 ± 0.89 d |
| 30 | 2.10 ± 0.10 b | 34.10 ± 0.72 e |
| 32 | 2.02 ± 0.16 b | 30.27 ± 0.34 a |
| 35 | 1.01 ± 0.06 e | 12.64 ± 0.42 f |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karageorgou, D.; Sainis, I.; Touka, A.; Vareli, K.; Stamatis, H.; Katapodis, P. Biomass and β-Glucosidase Production by the Cyanobacterium Pseudanabaena sp. under Heterotrophic Conditions. Biomass 2022, 2, 299-315. https://doi.org/10.3390/biomass2040020
Karageorgou D, Sainis I, Touka A, Vareli K, Stamatis H, Katapodis P. Biomass and β-Glucosidase Production by the Cyanobacterium Pseudanabaena sp. under Heterotrophic Conditions. Biomass. 2022; 2(4):299-315. https://doi.org/10.3390/biomass2040020
Chicago/Turabian StyleKarageorgou, Dimitra, Ioannis Sainis, Anastasia Touka, Katerina Vareli, Haralambos Stamatis, and Petros Katapodis. 2022. "Biomass and β-Glucosidase Production by the Cyanobacterium Pseudanabaena sp. under Heterotrophic Conditions" Biomass 2, no. 4: 299-315. https://doi.org/10.3390/biomass2040020
APA StyleKarageorgou, D., Sainis, I., Touka, A., Vareli, K., Stamatis, H., & Katapodis, P. (2022). Biomass and β-Glucosidase Production by the Cyanobacterium Pseudanabaena sp. under Heterotrophic Conditions. Biomass, 2(4), 299-315. https://doi.org/10.3390/biomass2040020