
Polystyrene Microplastics Modulate the Toxicity of the Hydrophilic Insecticide Thiacloprid for Chironomid Larvae and Also Influence Their Burrowing Behavior

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chironomus Riparius
2.2. Microplastics
2.2.1. Pre-Experiment with Fluorescent PS Particles
2.2.2. Polystyrene Particles Used in Main Experiments
2.3. Sediment
2.4. Test Solutions
2.5. Acute Toxicity Test
2.6. Burrowing Behavior
2.7. Chemical Analyses
2.7.1. Analyses of Thiacloprid
2.7.2. Sorption Behavior of Thiacloprid to PS
2.8. Statistical Analyses
3. Results
3.1. Pre-Experiment with Fluorescent PS
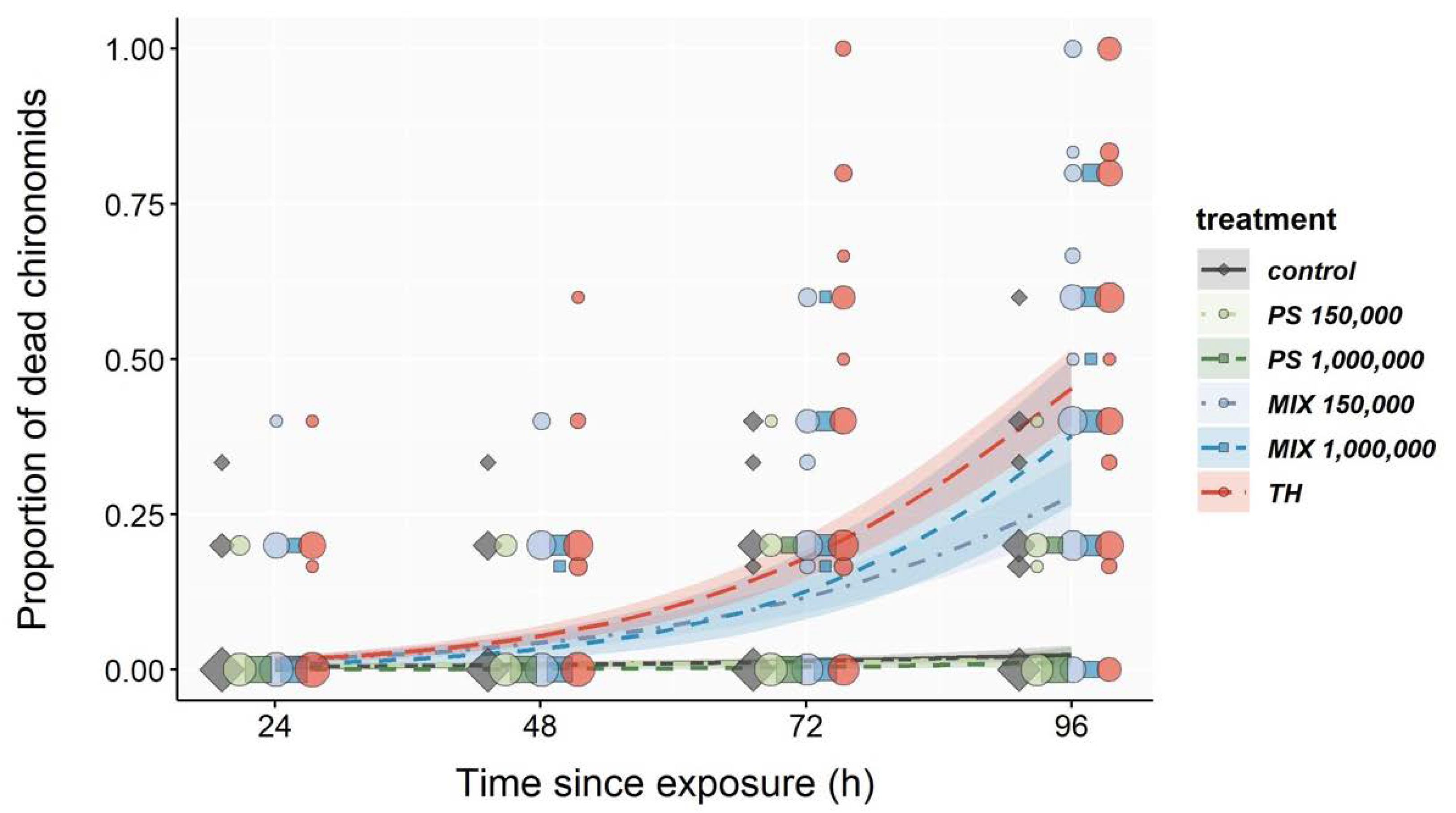
3.2. Acute Toxicity Test
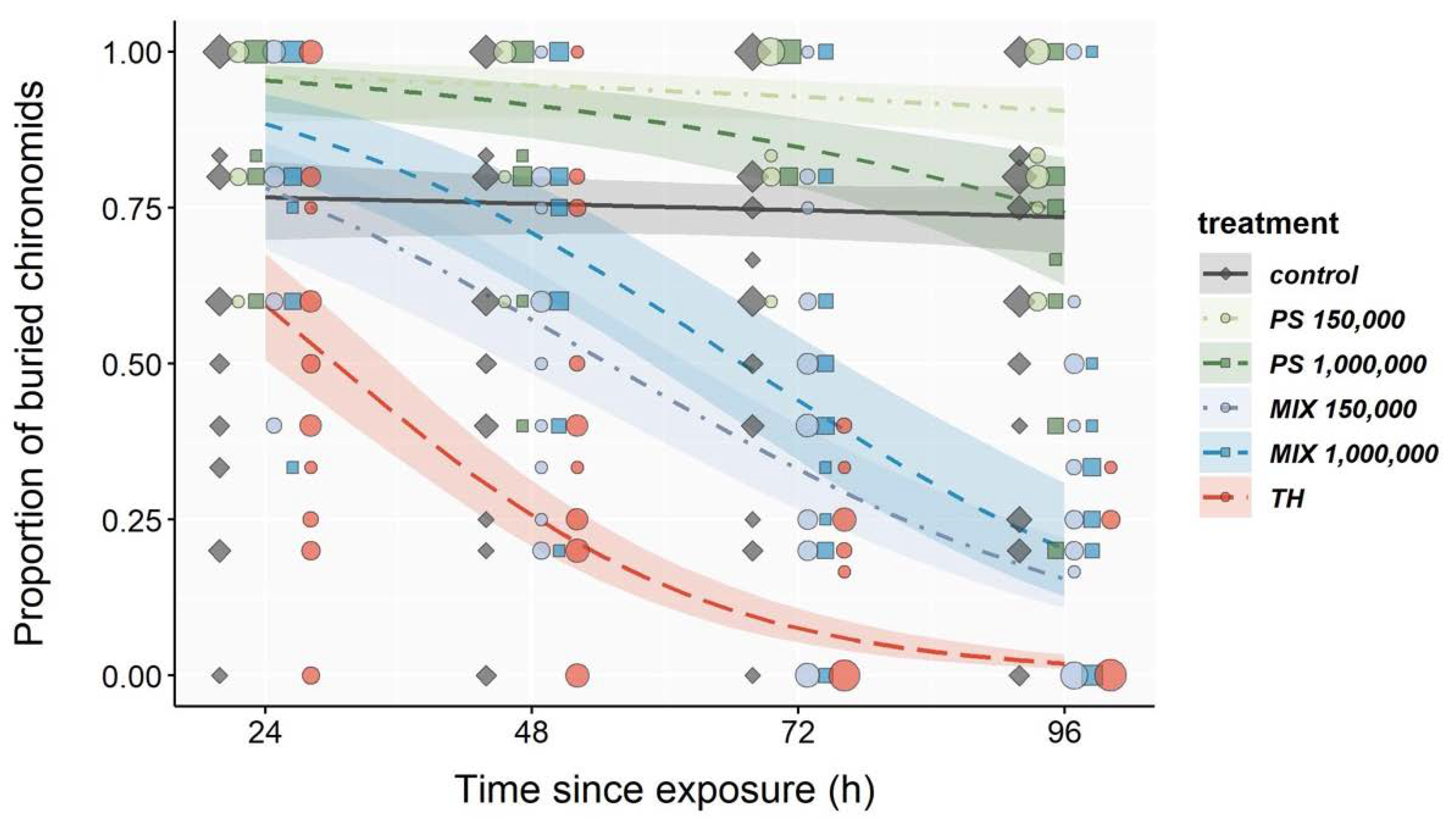
3.3. Burrowing Behavior
3.4. Chemical Analyses
3.4.1. Thiacloprid
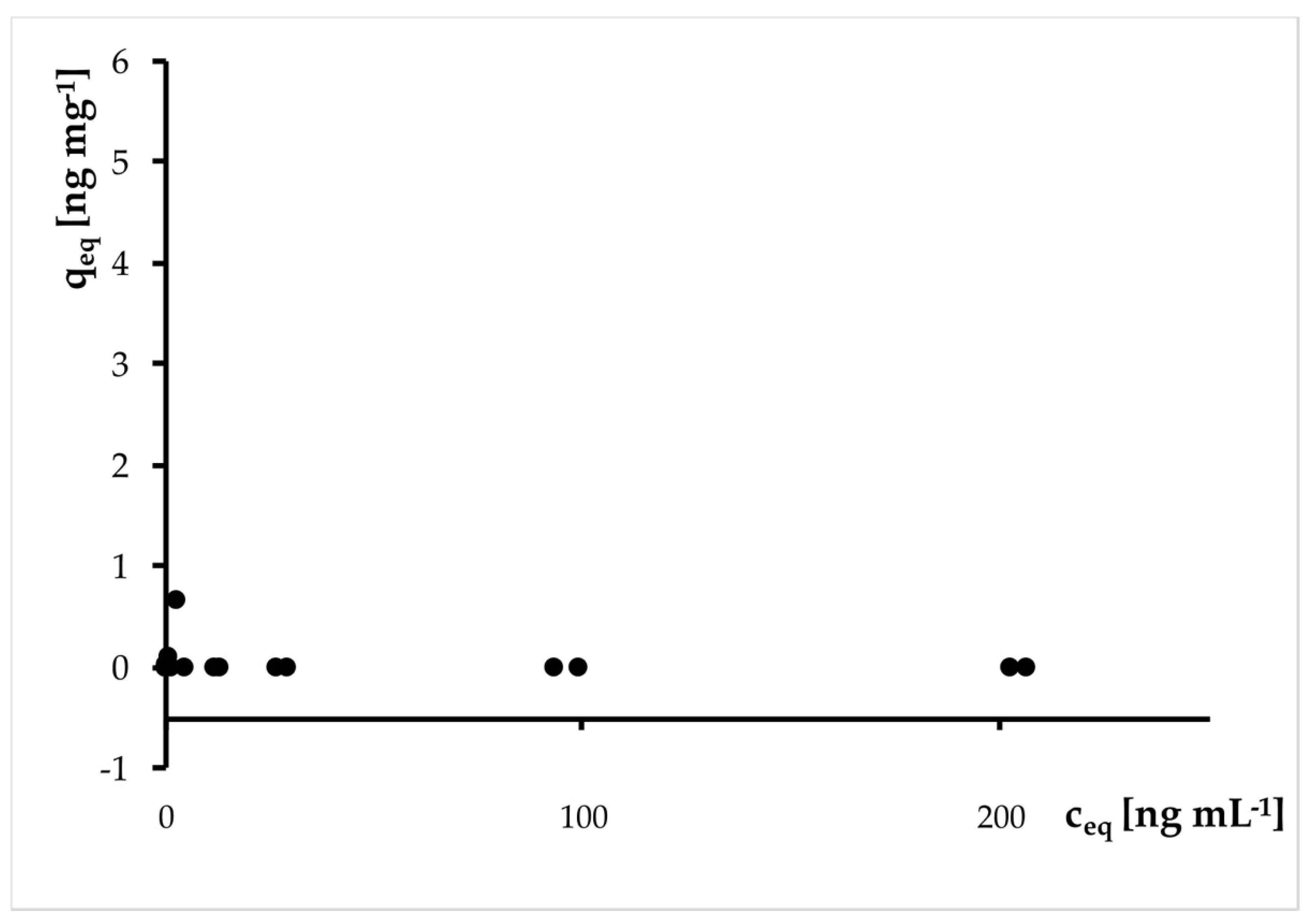
3.4.2. Sorption
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carpenter, E.J.; Smith, K.L. Plastics on the Sargasso Sea Surface. Science 1972, 175, 1240–1241. [Google Scholar] [CrossRef]
- Hartmann, N.B.; Hüffer, T.; Thompson, R.C.; Hassellöv, M.; Verschoor, A.; Daugaard, A.E.; Rist, S.; Karlsson, T.; Brennholt, N.; Cole, M.; et al. Are We Speaking the Same Language? Recommendations for a Definition and Categorization Framework for Plastic Debris. Environ. Sci. Technol. 2019, 53, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Imhof, H.K.; Ivleva, N.P.; Schmid, J.; Niessner, R.; Laforsch, C. Contamination of beach sediments of a subalpine lake with microplastic particles. Curr. Biol. 2013, 23, R867–R868. [Google Scholar] [CrossRef] [PubMed]
- Horton, A.A.; Vijver, M.G.; Lahive, E.; Spurgeon, D.J.; Svendsen, C.; Heutink, R.; van Bodegom, P.M.; Baas, J. Acute toxicity of organic pesticides to Daphnia magna is unchanged by co-exposure to polystyrene microplastics. Ecotoxicol. Environ. Saf. 2018, 166, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.; Worch, E.; Knepper, T.P. Occurrence and Spatial Distribution of Microplastics in River Shore Sediments of the Rhine-Main Area in Germany. Environ. Sci. Technol. 2015, 49, 6070–6076. [Google Scholar] [CrossRef]
- Triebskorn, R.; Braunbeck, T.; Grummt, T.; Hanslik, L.; Huppertsberg, S.; Jekel, M.; Knepper, T.P.; Krais, S.; Müller, Y.K.; Pittroff, M. Relevance of nano-and microplastics for freshwater ecosystems: A critical review. TrAC Trends Anal. Chem. 2019, 110, 375–392. [Google Scholar] [CrossRef]
- Li, C.; Busquets, R.; Campos, L.C. Assessment of microplastics in freshwater systems: A review. Sci. Total Environ. 2020, 707, 135578. [Google Scholar] [CrossRef]
- Rillig, M.C.; Lehmann, A. Microplastic in terrestrial ecosystems. Science 2020, 368, 1430–1431. [Google Scholar] [CrossRef]
- Lusher, A.L.; Tirelli, V.; O’Connor, I.; Officer, R. Microplastics in Arctic polar waters: The first reported values of particles in surface and sub-surface samples. Sci. Rep. 2015, 5, 14947. [Google Scholar] [CrossRef]
- Zhang, K.; Su, J.; Xiong, X.; Wu, X.; Wu, C.; Liu, J. Microplastic pollution of lakeshore sediments from remote lakes in Tibet plateau, China. Environ. Pollut. 2016, 219, 450–455. [Google Scholar] [CrossRef]
- Woodall, L.C.; Sanchez-Vidal, A.; Canals, M.; Paterson, G.L.; Coppock, R.; Sleight, V.; Calafat, A.; Rogers, A.D.; Narayanaswamy, B.E.; Thompson, R.C. The deep sea is a major sink for microplastic debris. R. Soc. Open Sci. 2014, 1, 140317. [Google Scholar] [CrossRef] [PubMed]
- Materić, D.; Kasper-Giebl, A.; Kau, D.; Anten, M.; Greilinger, M.; Ludewig, E.; van Sebille, E.; Röckmann, T.; Holzinger, R. Micro-and nanoplastics in Alpine Snow: A new method for chemical identification and (semi) quantification in the nanogram range. Environ. Sci. Technol. 2020, 54, 2353–2359. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, B. Ingestion of microplastics by fish and its potential consequences from a physical perspective. Integr. Environ. Assess. Manag. 2017, 13, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Cordova, M.R.; Riani, E.; Shiomoto, A. Microplastics ingestion by blue panchax fish (Aplocheilus sp.) from Ciliwung Estuary, Jakarta, Indonesia. Mar. Pollut. Bull. 2020, 161, 111763. [Google Scholar] [CrossRef]
- Wang, W.; Ge, J.; Yu, X. Bioavailability and toxicity of microplastics to fish species: A review. Ecotoxicol. Environ. Saf. 2020, 189, 109913. [Google Scholar] [CrossRef] [PubMed]
- Rochman, C.M.; Tahir, A.; Williams, S.L.; Baxa, D.V.; Lam, R.; Miller, J.T.; Teh, F.-C.; Werorilangi, S.; Teh, S.J. Anthropogenic debris in seafood: Plastic debris and fibers from textiles in fish and bivalves sold for human consumption. Sci. Rep. 2015, 5, 14340. [Google Scholar] [CrossRef]
- Van Cauwenberghe, L.; Claessens, M.; Vandegehuchte, M.B.; Janssen, C.R. Microplastics are taken up by mussels (Mytilus edulis) and lugworms (Arenicola marina) living in natural habitats. Environ. Pollut. 2015, 199, 10–17. [Google Scholar] [CrossRef]
- Weber, A.; Jeckel, N.; Wagner, M. Combined effects of polystyrene microplastics and thermal stress on the freshwater mussel Dreissena polymorpha. Sci. Total Environ. 2020, 718, 137253. [Google Scholar] [CrossRef]
- Nel, H.A.; Dalu, T.; Wasserman, R.J. Sinks and sources: Assessing microplastic abundance in river sediment and deposit feeders in an Austral temperate urban river system. Sci. Total Environ. 2018, 612, 950–956. [Google Scholar] [CrossRef]
- Cole, M.; Lindeque, P.; Fileman, E.; Halsband, C.; Goodhead, R.; Moger, J.; Galloway, T.S. Microplastic Ingestion by Zooplankton. Environ. Sci. Technol. 2013, 47, 6646–6655. [Google Scholar] [CrossRef]
- Scherer, C.; Weber, A.; Lambert, S.; Wagner, M. Interactions of microplastics with freshwater biota. In Freshwater Microplastics; Springer: Cham, Switzerland, 2018; pp. 153–180. [Google Scholar]
- Browne, M.A.; Dissanayake, A.; Galloway, T.S.; Lowe, D.M.; Thompson, R.C. Ingested Microscopic Plastic Translocates to the Circulatory System of the Mussel, Mytilus edulis (L.). Environ. Sci. Technol. 2008, 42, 5026–5031. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, Y.; Deng, Y.; Jiang, W.; Zhao, Y.; Geng, J.; Ding, L.; Ren, H. Uptake and Accumulation of Polystyrene Microplastics in Zebrafish (Danio rerio) and Toxic Effects in Liver. Environ. Sci. Technol. 2016, 50, 4054–4060. [Google Scholar] [CrossRef]
- Ramsperger, A.; Narayana, V.K.B.; Groß, W.; Mohanraj, J.; Thelakkat, M.; Greiner, A.; Schmalz, H.; Kress, H.; Laforsch, C. Environmental exposure enhances the internalization of microplastic particles into cells. Sci. Adv. 2020, 6, eabd1211. [Google Scholar] [CrossRef]
- Hahladakis, J.N.; Velis, C.A.; Weber, R.; Iacovidou, E.; Purnell, P. An overview of chemical additives present in plastics: Migration, release, fate and environmental impact during their use, disposal and recycling. J. Hazard. Mater. 2018, 344, 179–199. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Allgeier, A.; Yin, D.; Hollert, H. Leaching of endocrine disrupting chemicals from marine microplastics and mesoplastics under common life stress conditions. Environ. Int. 2019, 130, 104938. [Google Scholar] [CrossRef]
- Luo, H.; Li, Y.; Zhao, Y.; Xiang, Y.; He, D.; Pan, X. Effects of accelerated aging on characteristics, leaching, and toxicity of commercial lead chromate pigmented microplastics. Environ. Pollut. 2020, 257, 113475. [Google Scholar] [CrossRef] [PubMed]
- Au, S.Y.; Bruce, T.F.; Bridges, W.C.; Klaine, S.J. Responses of Hyalella azteca to acute and chronic microplastic exposures. Environ. Toxicol. Chem. 2015, 34, 2564–2572. [Google Scholar] [CrossRef]
- Ziajahromi, S.; Kumar, A.; Neale, P.A.; Leusch, F.D. Impact of microplastic beads and fibers on waterflea (Ceriodaphnia dubia) survival, growth, and reproduction: Implications of single and mixture exposures. Environ. Sci. Technol. 2017, 51, 13397–13406. [Google Scholar] [CrossRef]
- Redondo-Hasselerharm, P.E.; Falahudin, D.; Peeters, E.T.H.M.; Koelmans, A.A. Microplastic Effect Thresholds for Freshwater Benthic Macroinvertebrates. Environ. Sci. Technol. 2018, 52, 2278–2286. [Google Scholar] [CrossRef]
- Ziajahromi, S.; Kumar, A.; Neale, P.A.; Leusch, F.D. Environmentally relevant concentrations of polyethylene microplastics negatively impact the survival, growth and emergence of sediment-dwelling invertebrates. Environ. Pollut. 2018, 236, 425–431. [Google Scholar] [CrossRef]
- Silva, C.J.; Silva, A.L.P.; Gravato, C.; Pestana, J.L. Ingestion of small-sized and irregularly shaped polyethylene microplastics affect Chironomus riparius life-history traits. Sci. Total Environ. 2019, 672, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Scherer, C.; Wolf, R.; Völker, J.; Stock, F.; Brennhold, N.; Reifferscheid, G.; Wagner, M. Toxicity of microplastics and natural particles in the freshwater dipteran Chironomus riparius: Same same but different? Sci. Total Environ. 2020, 711, 134604. [Google Scholar] [CrossRef] [PubMed]
- Khosrovyan, A.; Doria, H.B.; Kahru, A.; Pfenninger, M. Polyamide microplastic exposure elicits rapid, strong and genome-wide evolutionary response in the freshwater non-biting midge Chironomus riparius. Chemosphere 2022, 299, 134452. [Google Scholar] [CrossRef]
- Straub, S.; Hirsch, P.E.; Burkhardt-Holm, P. Biodegradable and Petroleum-Based Microplastics Do Not Differ in Their Ingestion and Excretion but in Their Biological Effects in a Freshwater Invertebrate Gammarus fossarum. Int. J. Environ. Res. Public Health 2017, 14, 774. [Google Scholar] [CrossRef] [PubMed]
- Teuten, E.L.; Rowland, S.J.; Galloway, T.S.; Thompson, R.C. Potential for Plastics to Transport Hydrophobic Contaminants. Environ. Sci. Technol. 2007, 41, 7759–7764. [Google Scholar] [CrossRef]
- Fu, L.; Li, J.; Wang, G.; Luan, Y.; Dai, W. Adsorption behavior of organic pollutants on microplastics. Ecotoxicol. Environ. Saf. 2021, 217, 112207. [Google Scholar] [CrossRef]
- Hartmann, N.B.; Rist, S.; Bodin, J.; Jensen, L.H.; Schmidt, S.N.; Mayer, P.; Meibom, A.; Baun, A. Microplastics as vectors for environmental contaminants: Exploring sorption, desorption, and transfer to biota. Integr. Environ. Assess. Manag. 2017, 13, 488–493. [Google Scholar] [CrossRef]
- Koelmans, A.A.; Bakir, A.; Burton, G.A.; Janssen, C.R. Microplastic as a Vector for Chemicals in the Aquatic Environment: Critical Review and Model-Supported Reinterpretation of Empirical Studies. Environ. Sci. Technol. 2016, 50, 3315–3326. [Google Scholar] [CrossRef]
- Oliveira, M.; Ribeiro, A.; Hylland, K.; Guilhermino, L. Single and combined effects of microplastics and pyrene on juveniles (0+ group) of the common goby Pomatoschistus microps (Teleostei, Gobiidae). Ecol. Indic. 2013, 34, 641–647. [Google Scholar] [CrossRef]
- Kleinteich, J.; Seidensticker, S.; Marggrander, N.; Zarfl, C. Microplastics reduce short-term effects of environmental contaminants. Part II: Polyethylene particles decrease the effect of polycyclic aromatic hydrocarbons on microorganisms. Int. J. Environ. Res. Public Health 2018, 15, 287. [Google Scholar] [CrossRef] [Green Version]
- Sleight, V.A.; Bakir, A.; Thompson, R.C.; Henry, T.B. Assessment of microplastic-sorbed contaminant bioavailability through analysis of biomarker gene expression in larval zebrafish. Mar. Pollut. Bull. 2017, 116, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Avio, C.G.; Gorbi, S.; Milan, M.; Benedetti, M.; Fattorini, D.; d’Errico, G.; Pauletto, M.; Bargelloni, L.; Regoli, F. Pollutants bioavailability and toxicological risk from microplastics to marine mussels. Environ. Pollut. 2015, 198, 211–222. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency/Office of Pesticide Programs. Pesticide Chemical Search. Thiacloprid. (111988-49-9). Pesticide Fact Sheet; United States Environmental Protection Agency/Office of Pesticide Programs: Washington, DC, USA, 2003. [Google Scholar]
- Péry, A.R.R.; Garric, J. Modelling Effects of Temperature and Feeding Level on the Life Cycle of the Midge Chironomus Riparius: An Energy-Based Modelling Approach. Hydrobiologia 2006, 553, 59. [Google Scholar] [CrossRef]
- Armitage, P.D.; Pinder, L.; Cranston, P. The Chironomidae: Biology and Ecology of Non-Biting Midges; Chapman & Hall: London, UK, 1995. [Google Scholar]
- Organisation for Economic Co-operation and Development (OECD). Test No. 218: Sediment-Water Chironomid Toxicity Using Spiked Sediment; Organisation for Economic Co-operation and Development: Paris, France, 2004. [Google Scholar]
- Organisation for Economic Co-operation and Development (OECD). Test No. 233: Sediment-Water Chironomid Life-Cycle Toxicity Test Using Spiked Water or Spiked Sediment; Organisation for Economic Co-operation and Development: Paris, France, 2010. [Google Scholar]
- Langer-Jaesrich, M.; Köhler, H.-R.; Gerhardt, A. Assessing toxicity of the insecticide thiacloprid on Chironomus riparius (Insecta: Diptera) using multiple end points. Arch. Environ. Contam. Toxicol. 2010, 58, 963–972. [Google Scholar]
- Stevens, M.; Helliwell, S.; Hughes, P. Toxicity of Bacillus thuringiensis var. israelensis formulations, spinosad, and selected synthetic insecticides to Chironomus tepperi larvae. J. Am. Mosq. Control Assoc. 2005, 21, 446–450. [Google Scholar] [CrossRef]
- Lorenz, C.S.; Wicht, A.-J.; Guluzada, L.; Luo, L.; Jäger, L.; Crone, B.; Karst, U.; Triebskorn, R.; Liang, Y.; Anwander, R. Nano-sized Al2O3 reduces acute toxic effects of thiacloprid on the non-biting midge Chironomus riparius. PLoS ONE 2017, 12, e0176356. [Google Scholar]
- Rasmussen, J.B. Comparison of gut contents and assimilation efficiency of fourth instar larvae of two coexisting chironomids, Chironomus riparius Meigen and Glyptotendipes paripes (Edwards). Can. J. Zool. 1984, 62, 1022–1026. [Google Scholar] [CrossRef]
- Scherer, C.; Brennholt, N.; Reifferscheid, G.; Wagner, M. Feeding type and development drive the ingestion of microplastics by freshwater invertebrates. Sci. Rep. 2017, 7, 17006. [Google Scholar]
- Vos, J.H. Feeding of Detritivores in Freshwater Sediments; Universiteit van Amsterdam: Amsterdam, The Netherlands, 2001. [Google Scholar]
- Eitzen, L.; Paul, S.; Braun, U.; Altmann, K.; Jekel, M.; Ruhl, A.S. The challenge in preparing particle suspensions for aquatic microplastic research. Environ. Res. 2019, 168, 490–495. [Google Scholar] [CrossRef]
- Schmieg, H.; Burmester, J.K.Y.; Krais, S.; Ruhl, A.S.; Tisler, S.; Zwiener, C.; Köhler, H.-R.; Triebskorn, R. Interacting Effects of Polystyrene Microplastics and the Antidepressant Amitriptyline on Early Life Stages of Brown Trout (Salmo trutta f. fario). Water 2020, 12, 2361. [Google Scholar] [CrossRef]
- Schmieg, H.; Huppertsberg, S.; Knepper, T.P.; Krais, S.; Reitter, K.; Rezbach, F.; Ruhl, A.S.; Köhler, H.-R.; Triebskorn, R. Polystyrene microplastics do not affect juvenile brown trout (Salmo trutta f. fario) or modulate effects of the pesticide methiocarb. Environ. Sci. Eur. 2020, 32, 49. [Google Scholar] [CrossRef]
- Organisation for Economic Co-operation and Development(OECD). Test No. 235: Chironomus sp., Acute Immobilisation Test; Organisation for Economic Co-operation and Development: Paris, Feance, 2011. [Google Scholar]
- Day, K.E.; Kirby, R.S.; Reynoldson, T.B. Sexual dimorphism in Chironomus riparius (meigen): Impact on interpretation of growth in whole-sediment toxicity tests. Environ. Toxicol. Chem. 1994, 13, 35–39. [Google Scholar] [CrossRef]
- Pestana, J.L.T.; Loureiro, S.; Baird, D.J.; Soares, A.M.V.M. Fear and loathing in the benthos: Responses of aquatic insect larvae to the pesticide imidacloprid in the presence of chemical signals of predation risk. Aquat. Toxicol. 2009, 93, 138–149. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; Van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Machler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 1 August 2022).
- Ribeiro, P.J.; Diggle, P.J. geoR: A Package for Geostatistical Analysis. R-NEWS 2001, 1, 15–18. [Google Scholar]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R Package Version 0.4.1. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 1 August 2022).
- Korner-Nievergelt, F.; Roth, T.; Von Felten, S.; Guélat, J.; Almasi, B.; Korner-Nievergelt, P. Bayesian Data Analysis in Ecology Using Linear Models with R, BUGS, and Stan; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Halsey, L.G.; Curran-Everett, D.; Vowler, S.L.; Drummond, G.B. The fickle P value generates irreproducible results. Nat. Methods 2015, 12, 179–185. [Google Scholar] [CrossRef]
- Berner, D.; Amrhein, V. Why and how we should join the shift from significance testing to estimation. J. Evol. Biol. 2022, 35, 777–787. [Google Scholar] [CrossRef]
- Catarino, A.I.; Frutos, A.; Henry, T.B. Use of fluorescent-labelled nanoplastics (NPs) to demonstrate NP absorption is inconclusive without adequate controls. Sci. Total Environ. 2019, 670, 915–920. [Google Scholar] [CrossRef]
- Ziajahromi, S.; Kumar, A.; Neale, P.A.; Leusch, F.D. Effects of polyethylene microplastics on the acute toxicity of a synthetic pyrethroid to midge larvae (Chironomus tepperi) in synthetic and river water. Sci. Total Environ. 2019, 671, 971–975. [Google Scholar] [CrossRef]
- Bartonitz, A.; Anyanwu, I.N.; Geist, J.; Imhof, H.K.; Reichel, J.; Graßmann, J.; Drewes, J.E.; Beggel, S. Modulation of PAH toxicity on the freshwater organism G. roeseli by microparticles. Environ. Pollut. 2020, 260, 113999. [Google Scholar] [CrossRef]
- Naylor, C.; Rodrigues, C. Development of a test method for Chironomus riparius using a formulated sediment. Chemosphere 1995, 31, 3291–3303. [Google Scholar] [CrossRef]
- Halpern, M.; Gasith, A.; Broza, M. Does the tube of a benthic chironomid larva play a role in protecting its dweller against chemical toxicants? Hydrobiologia 2002, 470, 49–55. [Google Scholar] [CrossRef]
- Wentworth, C.K. A scale of grade and class terms for clastic sediments. J. Geol. 1922, 30, 377–392. [Google Scholar] [CrossRef]
- Khosrovyan, A.; Gabrielyan, B.; Kahru, A. Ingestion and effects of virgin polyamide microplastics on Chironomus riparius adult larvae and adult zebrafish Danio rerio. Chemosphere 2020, 259, 127456. [Google Scholar]
- Ben-Dov, E.; Saxena, D.; Wang, Q.; Manasherob, R.; Boussiba, S.; Zaritsky, A. Ingested particles reduce susceptibility of insect larvae to Bacillus thuringiensis. J. Appl. Entomol. 2003, 127, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Jarial, M.S.; Engstrom, L.E. Formation and Ultrastructure of the Peritrophic Membrane in Larval Midge Chironomus tetans (Diptera: Chironomidae). Zool. Sci. 1997, 14, 907–916. [Google Scholar] [CrossRef]
- Osborne, S.; Hurrell, S.; Simkiss, K.; Leidi, A. Factors influencing the distribution and feeding of the larvae of Chironomus riparius. Entomol. Exp. Appl. 2000, 94, 67–73. [Google Scholar] [CrossRef]
- Dadd, R. A method for comparing feeding rates in mosquito larvae. Mosq. News 1968, 28, 226–230. [Google Scholar]
- Dadd, R. Comparison of rates of ingestion of particulate solids by Culex pipiens larvae: Phagostimulant effect of water-soluble yeast extract. Entomol. Exp. Appl. 1970, 13, 407–419. [Google Scholar] [CrossRef]
- Nigamatzyanova, L.; Fakhrullin, R. Dark-field hyperspectral microscopy for label-free microplastics and nanoplastics detection and identification in vivo: A Caenorhabditis elegans study. Environ. Pollut. 2021, 271, 116337. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regression Estimates for Exposure Duration | % Dead After 96 h | ||||||
|---|---|---|---|---|---|---|---|
| Exposure Treatment | Coefficient Estimate * | SE | Lower 95% CI | Upper 95% CI | Odds-Ratio * | Estimate | 95% CI |
| Control | 0.0234 | 0.0054 | 0.0128 | 0.0341 | 1.024 | 2.4 | 1.5–3.9 |
| PS 150,000 | 0.0159 | 0.0065 | 0.0031 | 0.0287 | 1.016 | 1.8 | 0.9–3.1 |
| PS 1,000,000 | 0.0434 | 0.0221 | >−0.001 | 0.0868 | 1.044 | 1.3 | 0.4–3.7 |
| MIX 150,000 | 0.0449 | 0.0031 | 0.0388 | 0.0511 | 1.046 | 27.9 | 22.5–33.8 |
| MIX 1,000,000 | 0.0596 | 0.0067 | 0.0465 | 0.0727 | 1.061 | 37.7 | 26.5–50.2 |
| TH | 0.0553 | 0.0027 | 0.0499 | 0.0608 | 1.057 | 45.3 | 39.4–51.6 |
| Regression Estimates for Exposure Duration | % Buried After 96 h | ||||||
|---|---|---|---|---|---|---|---|
| Exposure Treatment | Coefficient Estimate | SE | Lower 95% CI | Upper 95% CI | Odds-Ratio | Estimate | 95% CI |
| Control | −0.0064 | 0.0034 | −0.0131 | 0.0001 | 0.994 | 73.1 | 67.6–78.1 |
| PS 150,000 | −0.0128 | 0.0092 | −0.0308 | 0.0053 | 0.987 | 89.7 | 83.9–93.5 |
| PS 1,000,000 | −0.0266 | 0.0069 | −0.0401 | −0.0130 | 0.974 | 73.5 | 62.4–81.9 |
| MIX 150,000 | −0.0428 | 0.0050 | −0.0527 | −0.0328 | 0.959 | 16.0 | 11.4–22.1 |
| MIX 1,000,000 | −0.0452 | 0.0060 | −0.0569 | −0.0334 | 0.956 | 21.4 | 14.1–31.4 |
| TH | −0.0575 | 0.0055 | −0.0684 | −0.0466 | 0.944 | 2.3 | 1.3–4.0 |
| Treatment | Sample Size (nStart) (nEnd) | Thiacloprid Conc. [µg/L]; (MV + SD) | Thiacloprid Rel. Recovery Rate [%]; (MV + SD) |
|---|---|---|---|
| TH | (n = 4) (n = 5) | 0.675 ± 0.170 0.552 ± 0.223 | 87.66 ± 22.08 71.69 ± 28.96 |
| MIX 150,000 | (n = 4) (n = 5) | 0.505 ± 0.241 0.610 ± 0.101 | 65.58 ± 31.17 79.22 ± 13.12 |
| MIX 1,000,000 | (n = 2) (n = 2) | 0.555 ± 0.025 0.625 ± 0.075 | 72.08 ± 3.25 81.17 ± 9.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krais, S.; Anthes, N.; Huppertsberg, S.; Knepper, T.P.; Peschke, K.; Ruhl, A.S.; Schmieg, H.; Schwarz, T.; Köhler, H.-R.; Triebskorn, R. Polystyrene Microplastics Modulate the Toxicity of the Hydrophilic Insecticide Thiacloprid for Chironomid Larvae and Also Influence Their Burrowing Behavior. Microplastics 2022, 1, 505-519. https://doi.org/10.3390/microplastics1030036
Krais S, Anthes N, Huppertsberg S, Knepper TP, Peschke K, Ruhl AS, Schmieg H, Schwarz T, Köhler H-R, Triebskorn R. Polystyrene Microplastics Modulate the Toxicity of the Hydrophilic Insecticide Thiacloprid for Chironomid Larvae and Also Influence Their Burrowing Behavior. Microplastics. 2022; 1(3):505-519. https://doi.org/10.3390/microplastics1030036
Chicago/Turabian StyleKrais, Stefanie, Nils Anthes, Sven Huppertsberg, Thomas P. Knepper, Katharina Peschke, Aki S. Ruhl, Hannah Schmieg, Tabea Schwarz, Heinz-R. Köhler, and Rita Triebskorn. 2022. "Polystyrene Microplastics Modulate the Toxicity of the Hydrophilic Insecticide Thiacloprid for Chironomid Larvae and Also Influence Their Burrowing Behavior" Microplastics 1, no. 3: 505-519. https://doi.org/10.3390/microplastics1030036
APA StyleKrais, S., Anthes, N., Huppertsberg, S., Knepper, T. P., Peschke, K., Ruhl, A. S., Schmieg, H., Schwarz, T., Köhler, H. -R., & Triebskorn, R. (2022). Polystyrene Microplastics Modulate the Toxicity of the Hydrophilic Insecticide Thiacloprid for Chironomid Larvae and Also Influence Their Burrowing Behavior. Microplastics, 1(3), 505-519. https://doi.org/10.3390/microplastics1030036