

Uptake and Cellular Effects of Polymethylmethacrylate on Human Cell Lines


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Confocal Imaging
2.3. Transfection of HEK293 Cells
2.4. Image Analysis
2.5. Beads
3. Results
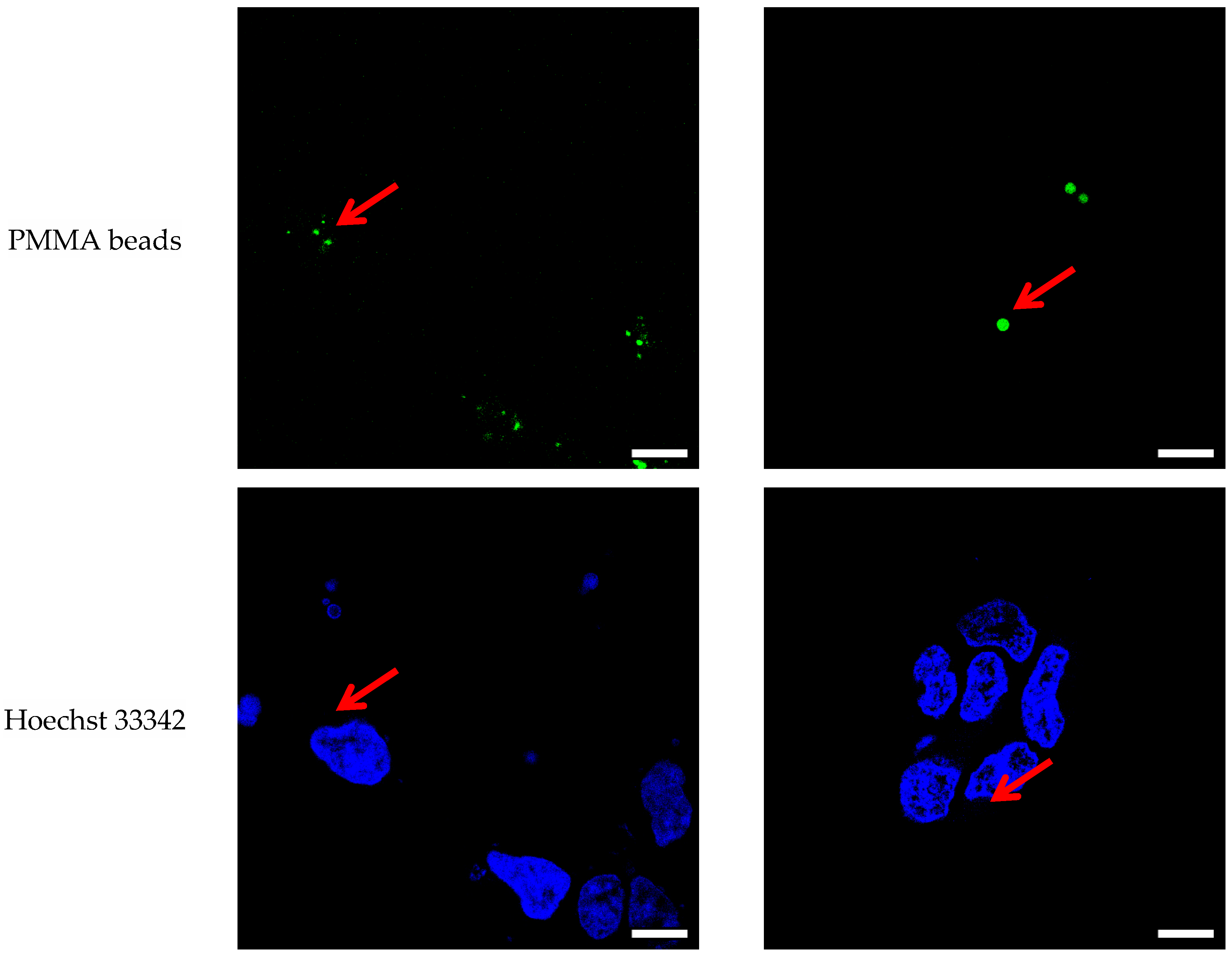
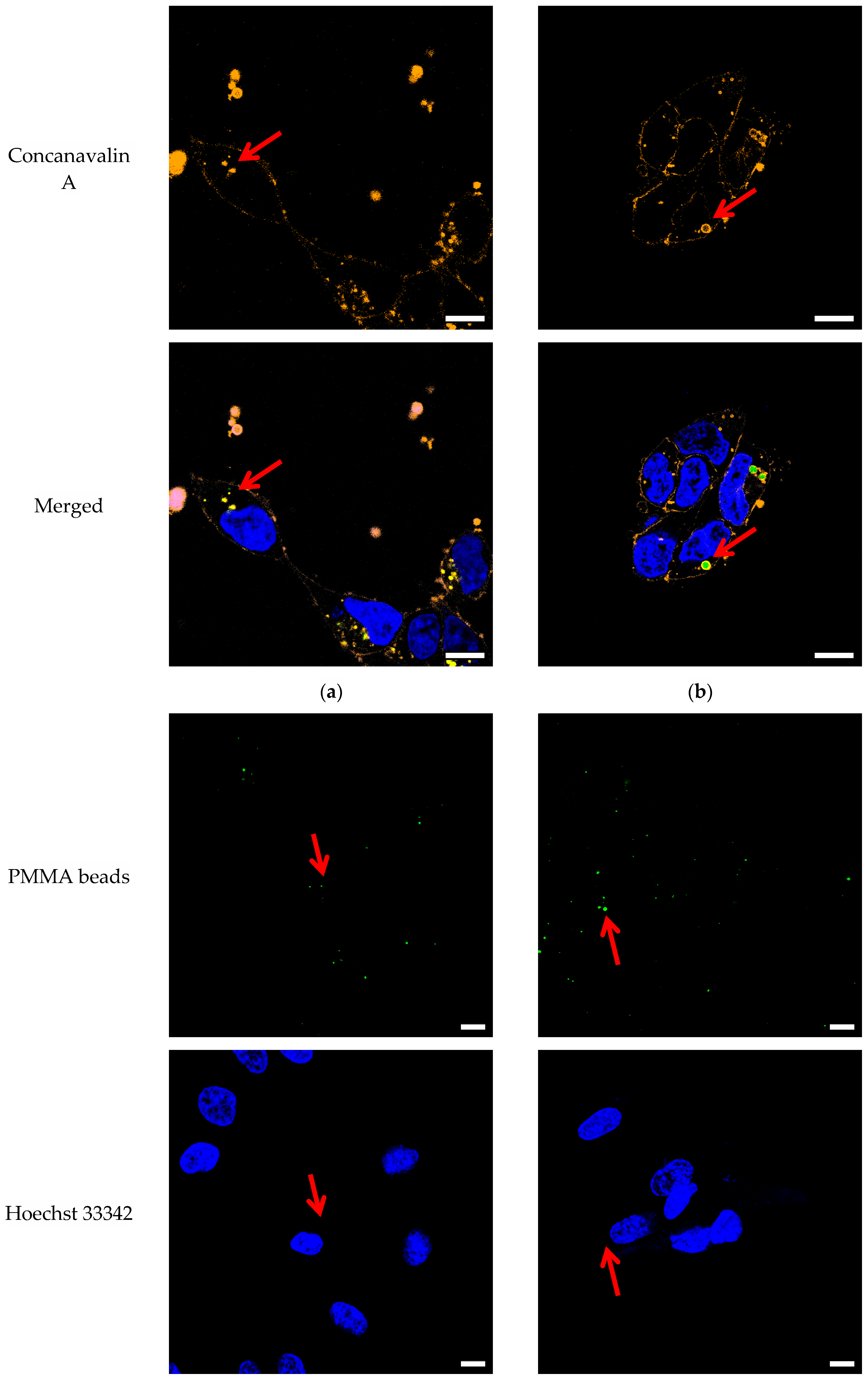
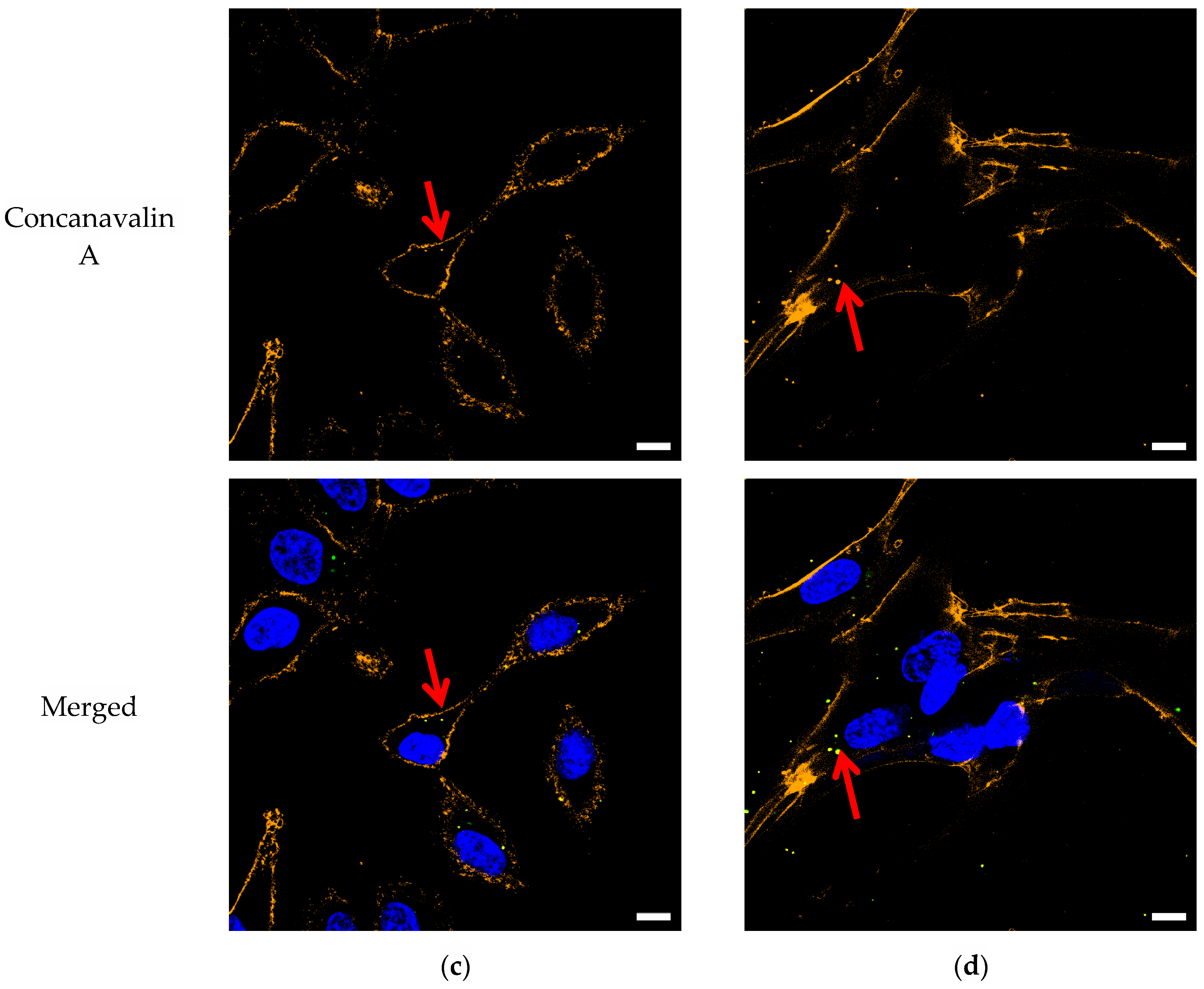
3.1. Confocal Imaging
3.2. Doubling Time
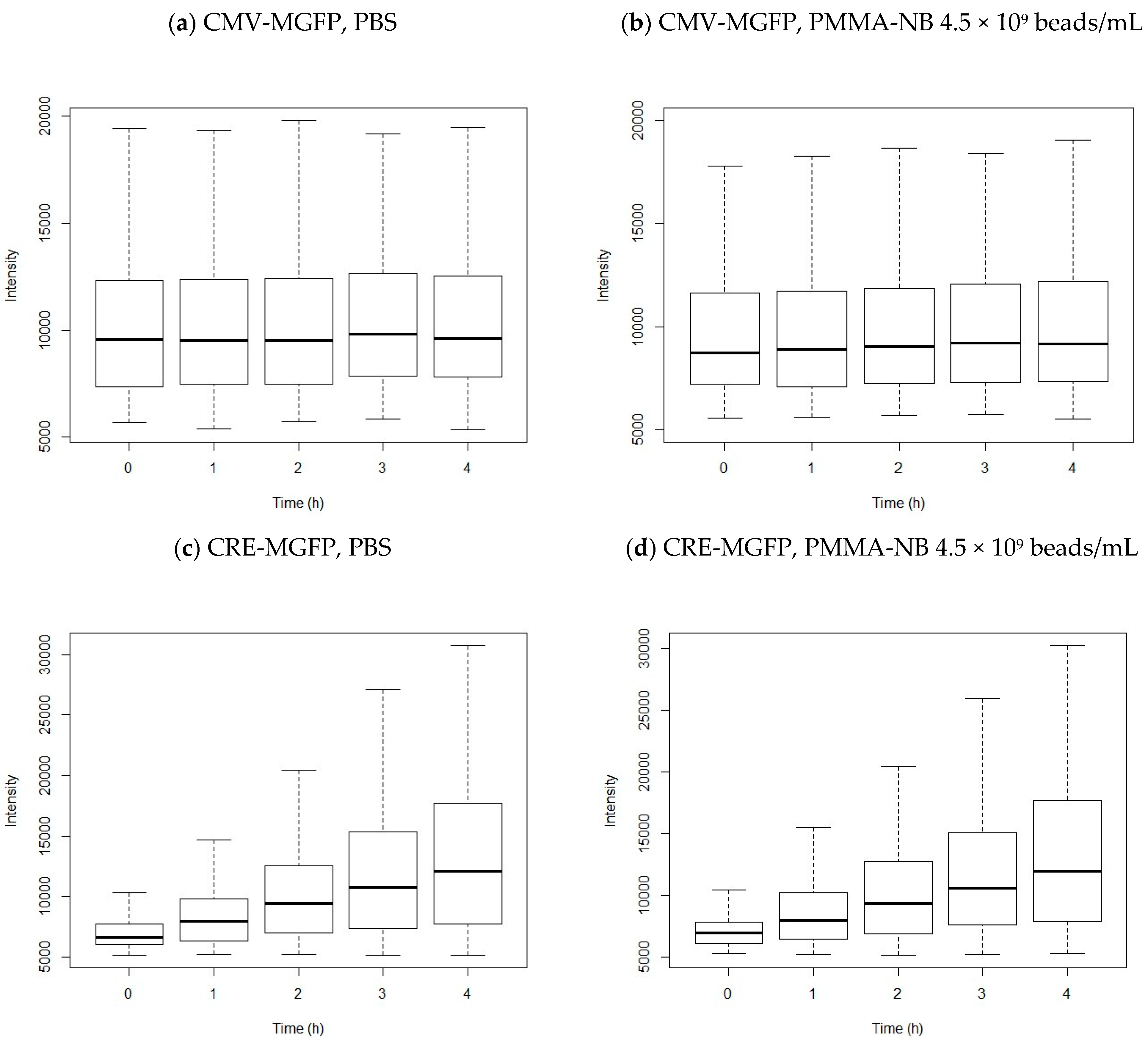
3.3. Transfection of HEK 293 Cells
4. Discussion
4.1. Confocal Imaging
4.2. Doubling Time
4.3. Transfection
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, Use, and Fate of All Plastics Ever Made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.J. Synthetic Polymers in the Marine Environment: A Rapidly Increasing, Long-Term Threat. Environ. Res. 2008, 108, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Ruz, V.; Gutow, L.; Thompson, R.C.; Thiel, M. Microplastics in the Marine Environment: A Review of the Methods Used for Identification and Quantification. Environ. Sci. Technol. 2012, 46, 3060–3075. [Google Scholar] [CrossRef]
- Yokota, K.; Waterfield, H.; Hastings, C.; Davidson, E.; Kwietniewski, E.; Wells, B. Finding the Missing Piece of the Aquatic Plastic Pollution Puzzle: Interaction between Primary Producers and Microplastics. Limnol. Oceanogr. Lett. 2017, 2, 91–104. [Google Scholar] [CrossRef]
- Caruso, G. Microplastics as Vectors of Contaminants. Mar. Pollut. Bull. 2019, 146, 921–924. [Google Scholar] [CrossRef] [PubMed]
- Xiang, C.; Chen, H.; Liu, X.; Dang, Y.; Li, X.; Yu, Y.; Li, B.; Li, X.; Sun, Y.; Ding, P.; et al. UV-Aged Microplastics Induces Neurotoxicity by Affecting the Neurotransmission in Larval Zebrafish. Chemosphere 2023, 324, 138252. [Google Scholar] [CrossRef] [PubMed]
- Qiao, R.; Sheng, C.; Lu, Y.; Zhang, Y.; Ren, H.; Lemos, B. Microplastics Induce Intestinal Inflammation, Oxidative Stress, and Disorders of Metabolome and Microbiome in Zebrafish. Sci. Total Environ. 2019, 662, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Vaishya, R.; Chauhan, M.; Vaish, A. Bone Cement. J. Clin. Orthop. Trauma 2013, 4, 157–163. [Google Scholar] [CrossRef]
- Becker, L.C.; Bergfeld, W.F.; Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.C.; Marks, J.G.; Shank, R.C.; Slaga, T.J.; Snyder, P.W.; et al. Final Report of the Cosmetic Ingredient Review Expert Panel Safety Assessment of Polymethyl Methacrylate (PMMA), Methyl Methacrylate Crosspolymer, and Methyl Methacrylate/Glycol Dimethacrylate Crosspolymer. Int. J. Toxicol. 2011, 30, 54S–65S. [Google Scholar] [CrossRef]
- Gad, M.; Fouda, S.; Al-Harbi, F.; Näpänkangas, R.; Raustia, A. PMMA Denture Base Material Enhancement: A Review of Fiber, Filler, and Nanofiller Addition. Int. J. Nanomedicine 2017, 12, 3801–3812. [Google Scholar] [CrossRef]
- Lin, M.; Wang, H.; Meng, S.; Zhong, W.; Li, Z.; Cai, R.; Chen, Z.; Zhou, X.; Du, Q. Structure and Release Behavior of PMMA/Silica Composite Drug Delivery System. J. Pharm. Sci. 2007, 96, 1518–1526. [Google Scholar] [CrossRef] [PubMed]
- Prabakaran, S.; Jeyaraj, M.; Nagaraj, A.; Sadasivuni, K.K.; Rajan, M. Polymethyl Methacrylate–Ovalbumin@Graphene Oxide Drug Carrier System for High Anti-Proliferative Cancer Drug Delivery. Appl. Nanosci. 2019, 9, 1487–1500. [Google Scholar] [CrossRef]
- Da Silva Brito, W.A.; Singer, D.; Miebach, L.; Saadati, F.; Wende, K.; Schmidt, A.; Bekeschus, S. Comprehensive in Vitro Polymer Type, Concentration, and Size Correlation Analysis to Microplastic Toxicity and Inflammation. Sci. Total Environ. 2023, 854, 158731. [Google Scholar] [CrossRef]
- Yang, Y.-F.; Chen, C.-Y.; Lu, T.-H.; Liao, C.-M. Toxicity-Based Toxicokinetic/Toxicodynamic Assessment for Bioaccumulation of Polystyrene Microplastics in Mice. J. Hazard. Mater. 2019, 366, 703–713. [Google Scholar] [CrossRef]
- Pironti, C.; Notarstefano, V.; Ricciardi, M.; Motta, O.; Giorgini, E.; Montano, L. First Evidence of Microplastics in Human Urine, a Preliminary Study of Intake in the Human Body. Toxics 2022, 11, 40. [Google Scholar] [CrossRef]
- Pellegrino, A.; Danne, D.; Weigel, C.; Seitz, H. An In Vitro Assay to Quantify Effects of Micro- and Nano-Plastics on Human Gene Transcription. Microplastics 2023, 2, 122–131. [Google Scholar] [CrossRef]
- Kida, S.; Serita, T. Functional Roles of CREB as a Positive Regulator in the Formation and Enhancement of Memory. Brain Res. Bull. 2014, 105, 17–24. [Google Scholar] [CrossRef]
- Oike, Y.; Takakura, N.; Hata, A.; Kaname, T.; Akizuki, M.; Yamaguchi, Y.; Yasue, H.; Araki, K.; Yamamura, K.; Suda, T. Mice Homozygous for a Truncated Form of CREB-Binding Protein Exhibit Defects in Hematopoiesis and Vasculo-Angiogenesis. Blood 1999, 93, 2771–2779. [Google Scholar] [CrossRef] [PubMed]
- Saura, C.A.; Valero, J. The Role of CREB Signaling in Alzheimer’s Disease and Other Cognitive Disorders. Revneuro 2011, 22, 153–169. [Google Scholar] [CrossRef]
- Shapiro, L.; Katchur, N. Microplastic Exposure and the of Parkinson’s Disease: The Effects of Microplastics in the Body and Similarities to the Pathogenesis of Parkinson’s Disease. J. Stud. Res. 2023, 11. [Google Scholar] [CrossRef]
- Thompson, R.; Casali, C.; Chan, C. Forskolin and IBMX Induce Neural Transdifferentiation of MSCs Through Downregulation of the NRSF. Sci. Rep. 2019, 9, 2969. [Google Scholar] [CrossRef]
- Pitt, J.A.; Kozal, J.S.; Jayasundara, N.; Massarsky, A.; Trevisan, R.; Geitner, N.; Wiesner, M.; Levin, E.D.; Di Giulio, R.T. Uptake, Tissue Distribution, and Toxicity of Polystyrene Nanoparticles in Developing Zebrafish (Danio Rerio). Aquat. Toxicol. 2018, 194, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, F.; Van Eck, G.; Cole, M.; Salvati, A.; Bos, S.; Gosens, R.; Melgert, B. Exposure to Textile Microplastic Fibers Impairs Epithelial Growth. Eur. Respir. J. 2020, 56, 1972. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, J.; Wang, W.; Gonzalez-Gil, G.; Vrouwenvelder, J.S.; Li, Z. Effects of Nano- and Microplastics on Kidney: Physicochemical Properties, Bioaccumulation, Oxidative Stress and Immunoreaction. Chemosphere 2022, 288, 132631. [Google Scholar] [CrossRef] [PubMed]
- Adinolfi, B.; Pellegrino, M.; Tombelli, S.; Trono, C.; Giannetti, A.; Domenici, C.; Varchi, G.; Sotgiu, G.; Ballestri, M.; Baldini, F. Polymeric Nanoparticles Promote Endocytosis of a Survivin Molecular Beacon: Localization and Fate of Nanoparticles and Beacon in Human A549 Cells. Life Sci. 2018, 215, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.P.; Jones, C.M.; Baille, J.P. Characteristics of a Human Diploid Cell Designated MRC-5. Nature 1970, 227, 168–170. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.C.; Graham, F.L.; Smiley, J.; Nairn, R. Characteristics of a Human Cell Line Transformed by DNA from Human Adenovirus Type 5. J. Gen. Virol. 1977, 36, 59–72. [Google Scholar] [CrossRef]
- Assanga, I. Cell Growth Curves for Different Cell Lines and Their Relationship with Biological Activities. Int. J. Biotechnol. Mol. Biol. Res. 2013, 4, 60–70. [Google Scholar] [CrossRef]
- Moore, T.L.; Rodriguez-Lorenzo, L.; Hirsch, V.; Balog, S.; Urban, D.; Jud, C.; Rothen-Rutishauser, B.; Lattuada, M.; Petri-Fink, A. Nanoparticle Colloidal Stability in Cell Culture Media and Impact on Cellular Interactions. Chem. Soc. Rev. 2015, 44, 6287–6305. [Google Scholar] [CrossRef]
- Vollrath, A.; Schallon, A.; Pietsch, C.; Schubert, S.; Nomoto, T.; Matsumoto, Y.; Kataoka, K.; Schubert, U.S. A Toolbox of Differently Sized and Labeled PMMA Nanoparticles for Cellular Uptake Investigations. Soft Matter 2013, 9, 99–108. [Google Scholar] [CrossRef]
- Saftig, P.; Klumperman, J. Lysosome Biogenesis and Lysosomal Membrane Proteins: Trafficking Meets Function. Nat. Rev. Mol. Cell Biol. 2009, 10, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Barral, D.C.; Staiano, L.; Guimas Almeida, C.; Cutler, D.F.; Eden, E.R.; Futter, C.E.; Galione, A.; Marques, A.R.A.; Medina, D.L.; Napolitano, G.; et al. Current Methods to Analyze Lysosome Morphology, Positioning, Motility and Function. Traffic 2022, 23, 238–269. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.A.; Akhtar, S.; Almohazey, D.; Alomari, M.; Almofty, S.A.; Badr, I.; Elaissari, A. Targeted Delivery of Poly (Methyl Methacrylate) Particles in Colon Cancer Cells Selectively Attenuates Cancer Cell Proliferation. Artif. Cells Nanomedicine Biotechnol. 2019, 47, 1533–1542. [Google Scholar] [CrossRef] [PubMed]
- Feuser, P.E.; Gaspar, P.C.; Ricci-Júnior, E.; Silva, M.C.S.D.; Nele, M.; Sayer, C.; De Araújo, P.H.H. Synthesis and Characterization of Poly(Methyl Methacrylate) PMMA and Evaluation of Cytotoxicity for Biomedical Application. Macromol. Symp. 2014, 343, 65–69. [Google Scholar] [CrossRef]
- Everaert, G.; Van Cauwenberghe, L.; De Rijcke, M.; Koelmans, A.A.; Mees, J.; Vandegehuchte, M.; Janssen, C.R. Risk Assessment of Microplastics in the Ocean: Modelling Approach and First Conclusions. Environ. Pollut. 2018, 242, 1930–1938. [Google Scholar] [CrossRef] [PubMed]
- Leslie, H.A.; Van Velzen, M.J.M.; Brandsma, S.H.; Vethaak, A.D.; Garcia-Vallejo, J.J.; Lamoree, M.H. Discovery and Quantification of Plastic Particle Pollution in Human Blood. Environ. Int. 2022, 163, 107199. [Google Scholar] [CrossRef] [PubMed]
- Windheim, J.; Colombo, L.; Battajni, N.C.; Russo, L.; Cagnotto, A.; Diomede, L.; Bigini, P.; Vismara, E.; Fiumara, F.; Gabbrielli, S.; et al. Micro- and Nanoplastics’ Effects on Protein Folding and Amyloidosis. Int. J. Mol. Sci. 2022, 23, 10329. [Google Scholar] [CrossRef]
- Wang, J.; Cong, J.; Wu, J.; Chen, Y.; Fan, H.; Wang, X.; Duan, Z.; Wang, L. Nanoplastic-Protein Corona Interactions and Their Biological Effects: A Review of Recent Advances and Trends. TrAC Trends Anal. Chem. 2023, 166, 117206. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Figure | (a) | (b) | (c) | (d) |
|---|---|---|---|---|
| Cell line | HEK293 | HEK293 | A549 | MRC5 |
| Average size (µm) determined | 0.54 ± 0.29 | 2.03 ± 0.19 | 0.76 ± 0.27 | 0.66 ± 0.47 |
| Average size (µm) of the beads | 0.165 | 2 | 0.165 | 0.165 |
| Polydispersity Index | 9.1% | 8.9% | 9.1% | 9.1% |
| Cell Line | Ø | Beads/mL | t0 (h) |
|---|---|---|---|
| HEK293 | 165 nm | 4.5 × 109 | 26.00 ± 3.85 |
| 4.5 × 108 | 25.96 ± 3.49 | ||
| 0 | 27.44 ± 2.33 | ||
| 2 µm | 2.53 × 106 | 28.84 ± 2.95 | |
| 2.53 × 105 | 29.94 ± 3.01 | ||
| 0 | 29.56 ± 2.75 | ||
| A549 | 165 nm | 4.5 × 109 | 30.84 ± 1.64 |
| 4.5 × 108 | 33.17 ± 3.89 | ||
| 0 | 29.86 ± 1.62 | ||
| 2 µm | 2.53 × 106 | 29.21 ± 1.36 | |
| 2.53 × 105 | 28.61 ± 3.30 | ||
| 0 | 29.44 ± 2.76 | ||
| MRC5 | 165 nm | 4.5 × 109 | 51.26 ± 7.91 |
| 4.5 × 108 | 46.30 ± 4.66 | ||
| 0 | 45.44 ± 4.37 | ||
| 2 µm | 2.53 × 106 | 45.16 ± 7.56 | |
| 2.53 × 105 | 47.48 ± 9.07 | ||
| 0 | 46.25 ± 7.32 |
| 0 h | 1 h | 2 h | 3 h | 4 h | |
|---|---|---|---|---|---|
| CMV-MGFP, PBS | 316 | 329 | 369 | 389 | 432 |
| CMV-MGFP, PMMA-NB 4.5 × 109 beads/mL | 299 | 343 | 376 | 423 | 491 |
| CRE-MGFP, PBS | 314 | 575 | 720 | 788 | 837 |
| CRE-MGFP, PMMA-NB 4.5 × 109 beads/mL | 323 | 551 | 701 | 781 | 794 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braun, A.; Seitz, H. Uptake and Cellular Effects of Polymethylmethacrylate on Human Cell Lines. Microplastics 2024, 3, 205-216. https://doi.org/10.3390/microplastics3020012
Braun A, Seitz H. Uptake and Cellular Effects of Polymethylmethacrylate on Human Cell Lines. Microplastics. 2024; 3(2):205-216. https://doi.org/10.3390/microplastics3020012
Chicago/Turabian StyleBraun, Arthur, and Harald Seitz. 2024. "Uptake and Cellular Effects of Polymethylmethacrylate on Human Cell Lines" Microplastics 3, no. 2: 205-216. https://doi.org/10.3390/microplastics3020012
APA StyleBraun, A., & Seitz, H. (2024). Uptake and Cellular Effects of Polymethylmethacrylate on Human Cell Lines. Microplastics, 3(2), 205-216. https://doi.org/10.3390/microplastics3020012