
Laboratory Rat Thrombi Lose One-Third of Their Stiffness When Exposed to Large Oscillating Shear Stress Amplitudes: Contrasting Behavior to Human Clots



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Blood Samples
2.3. Rheometry
2.4. Experimental Protocols
2.5. Other Laboratory Data
2.6. Scanning Electron Microscopy (SEM)
2.7. Statistic
3. Results
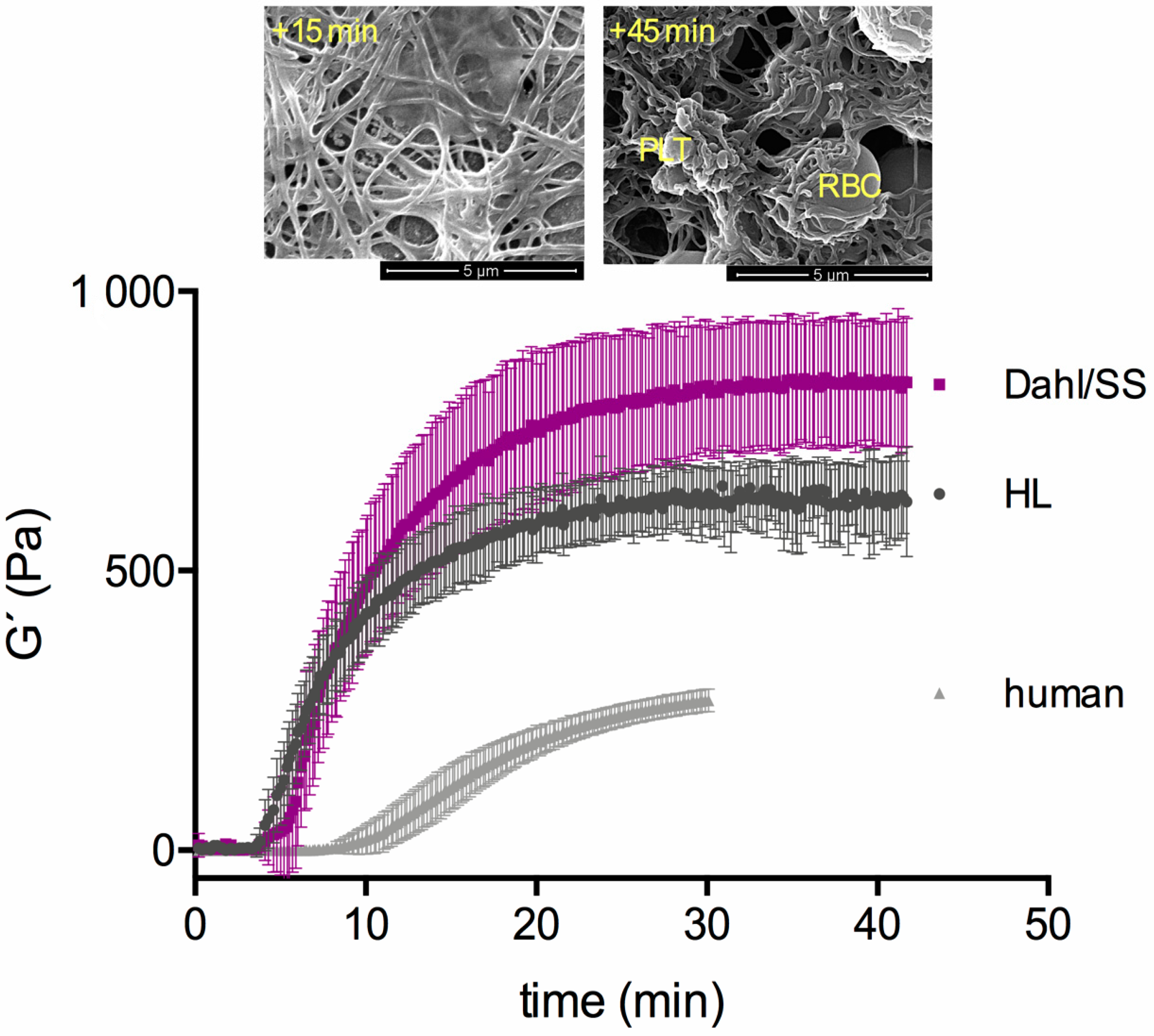
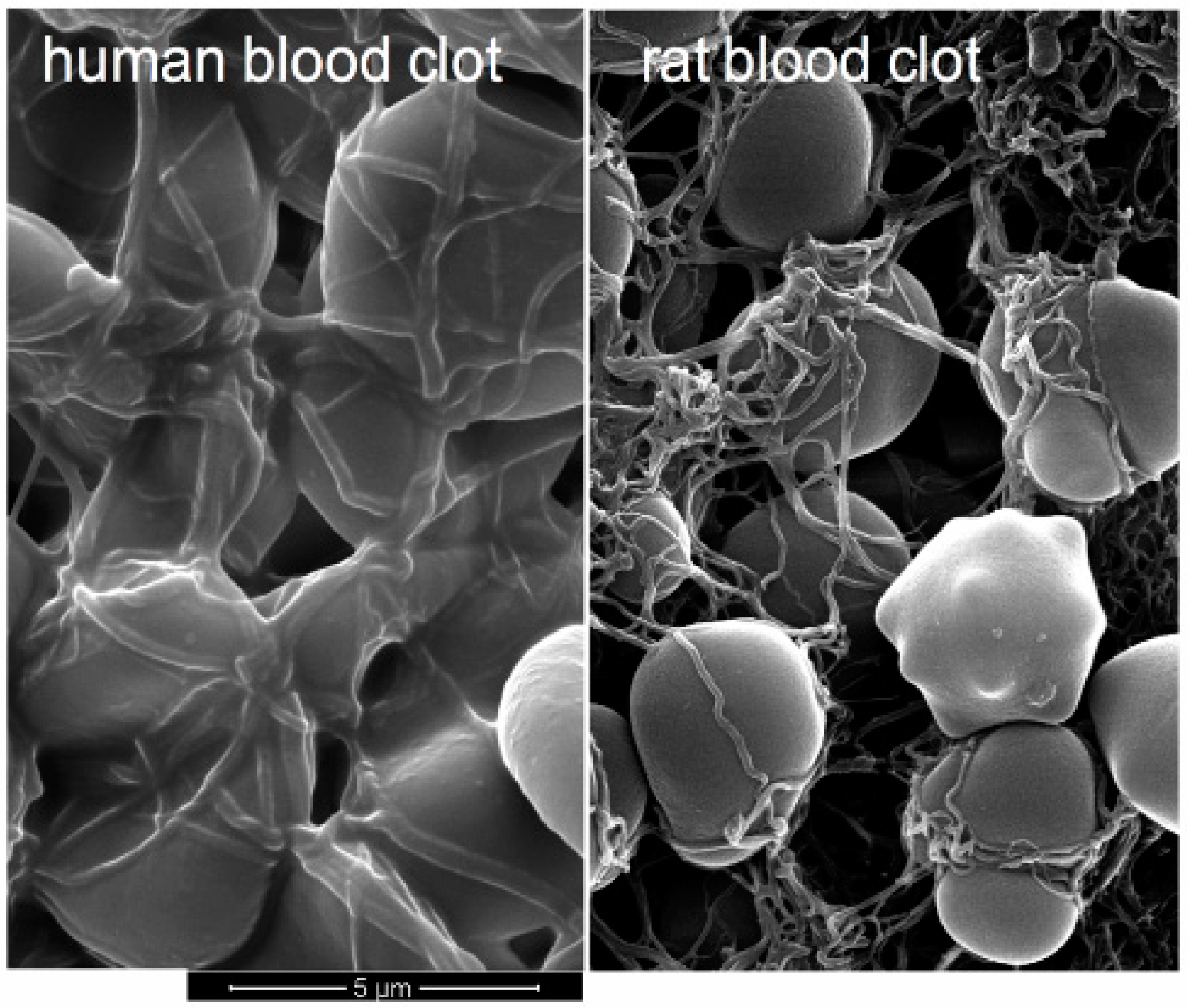
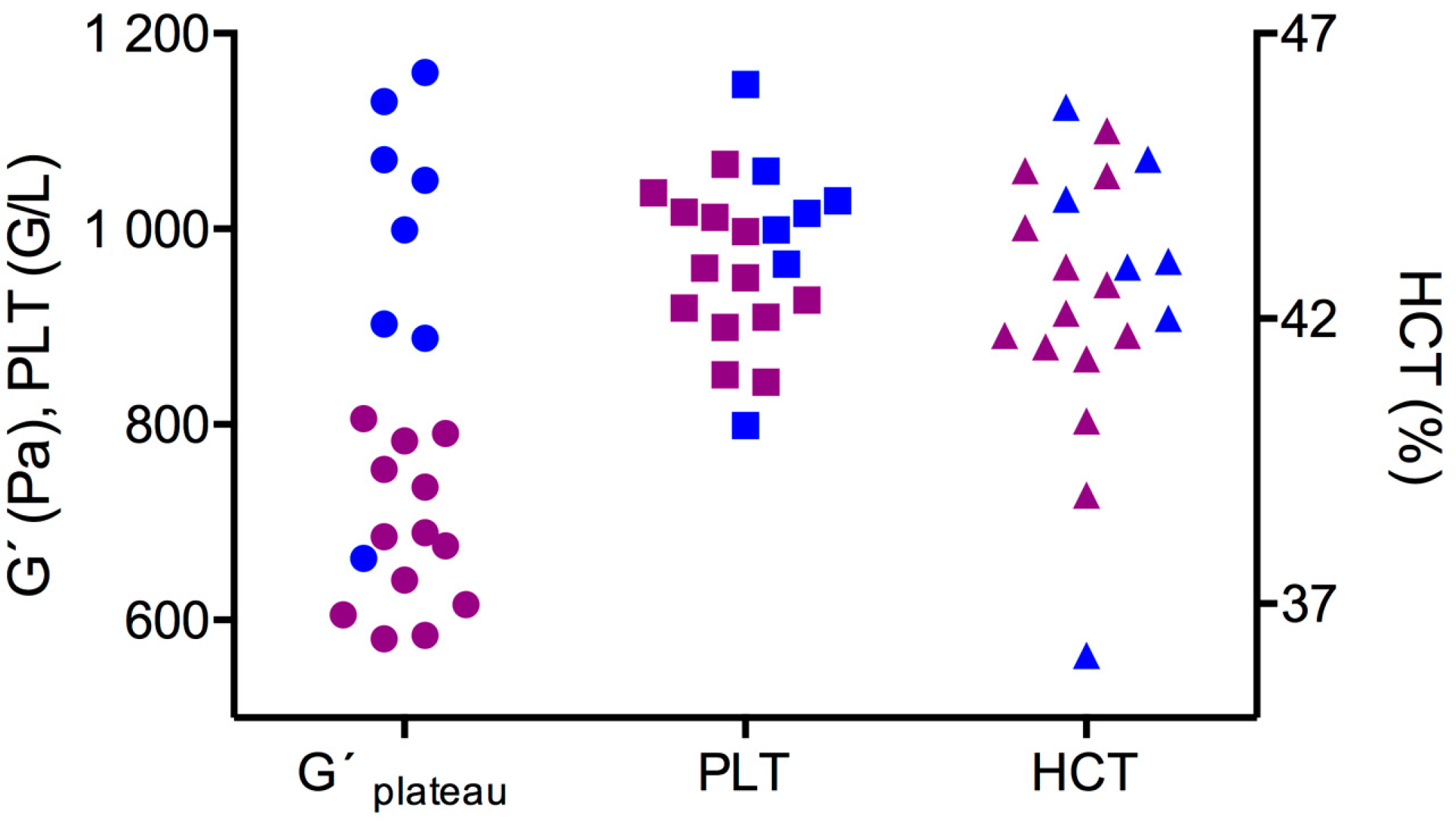
3.1. First Test Series
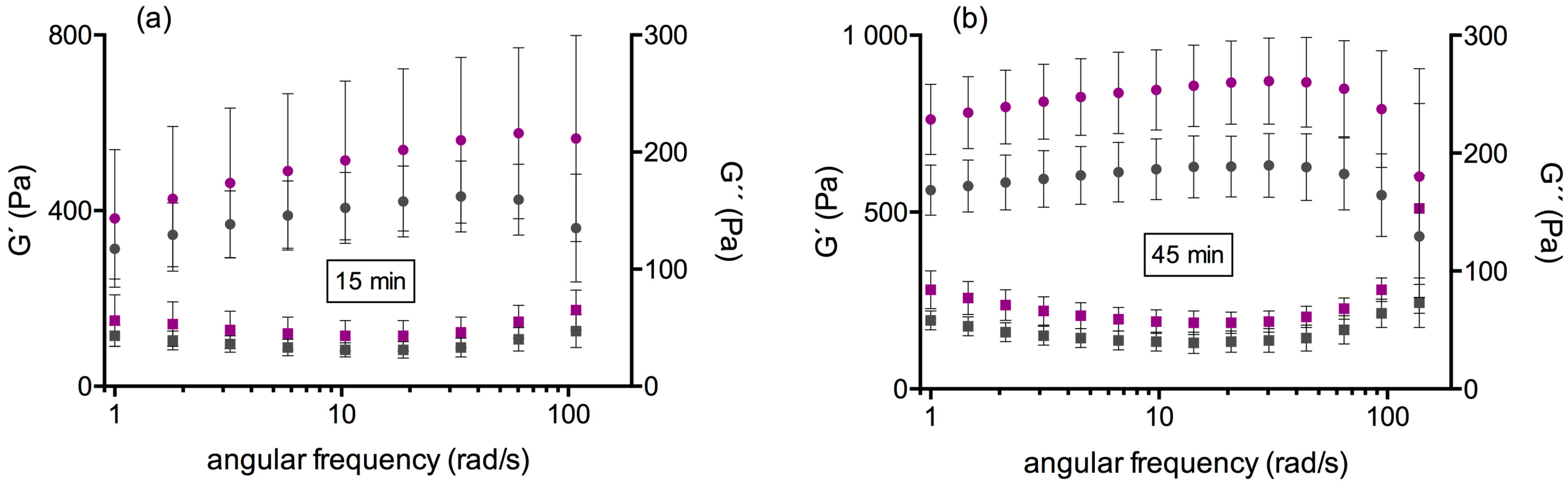
3.2. Second Test Series
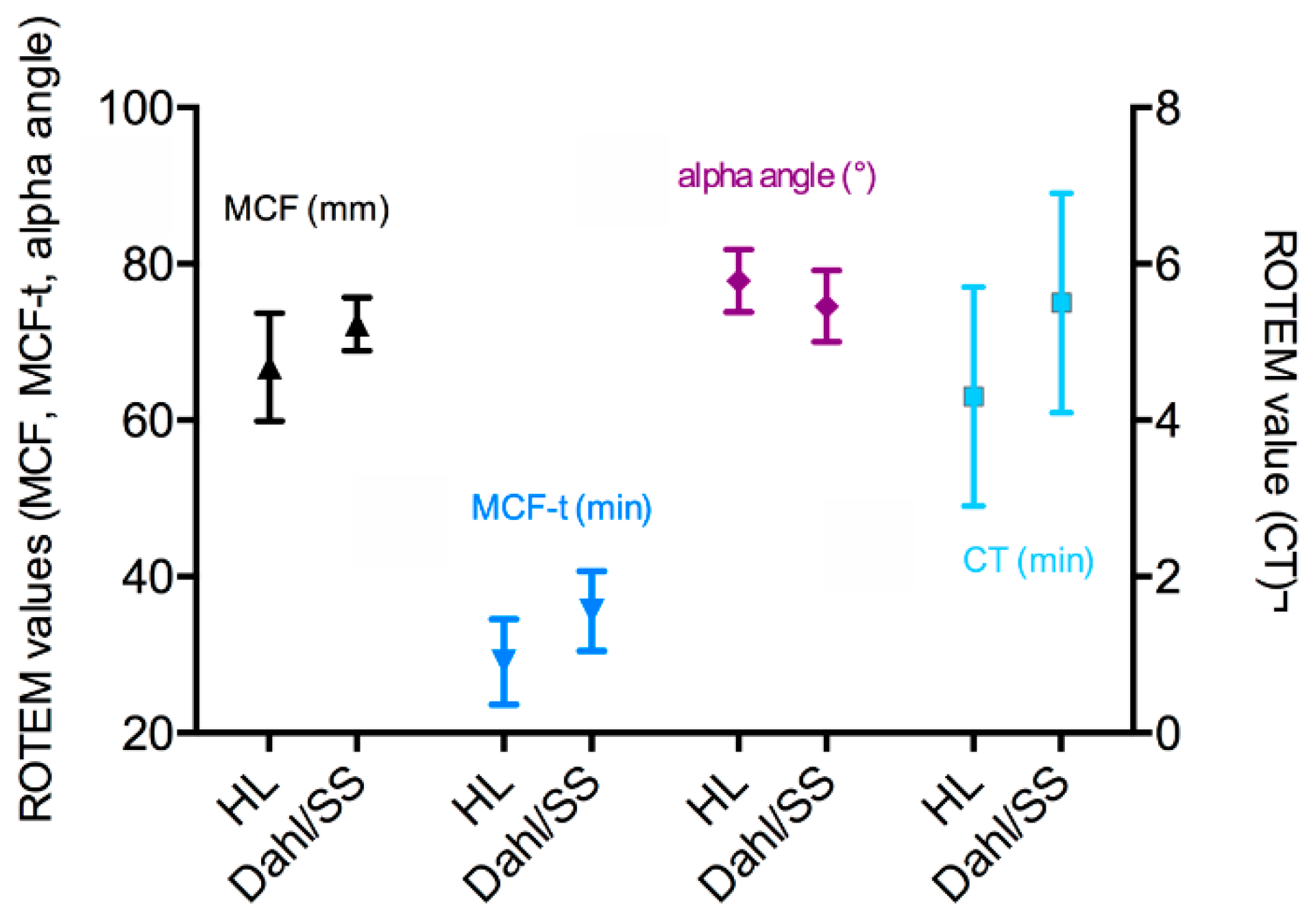
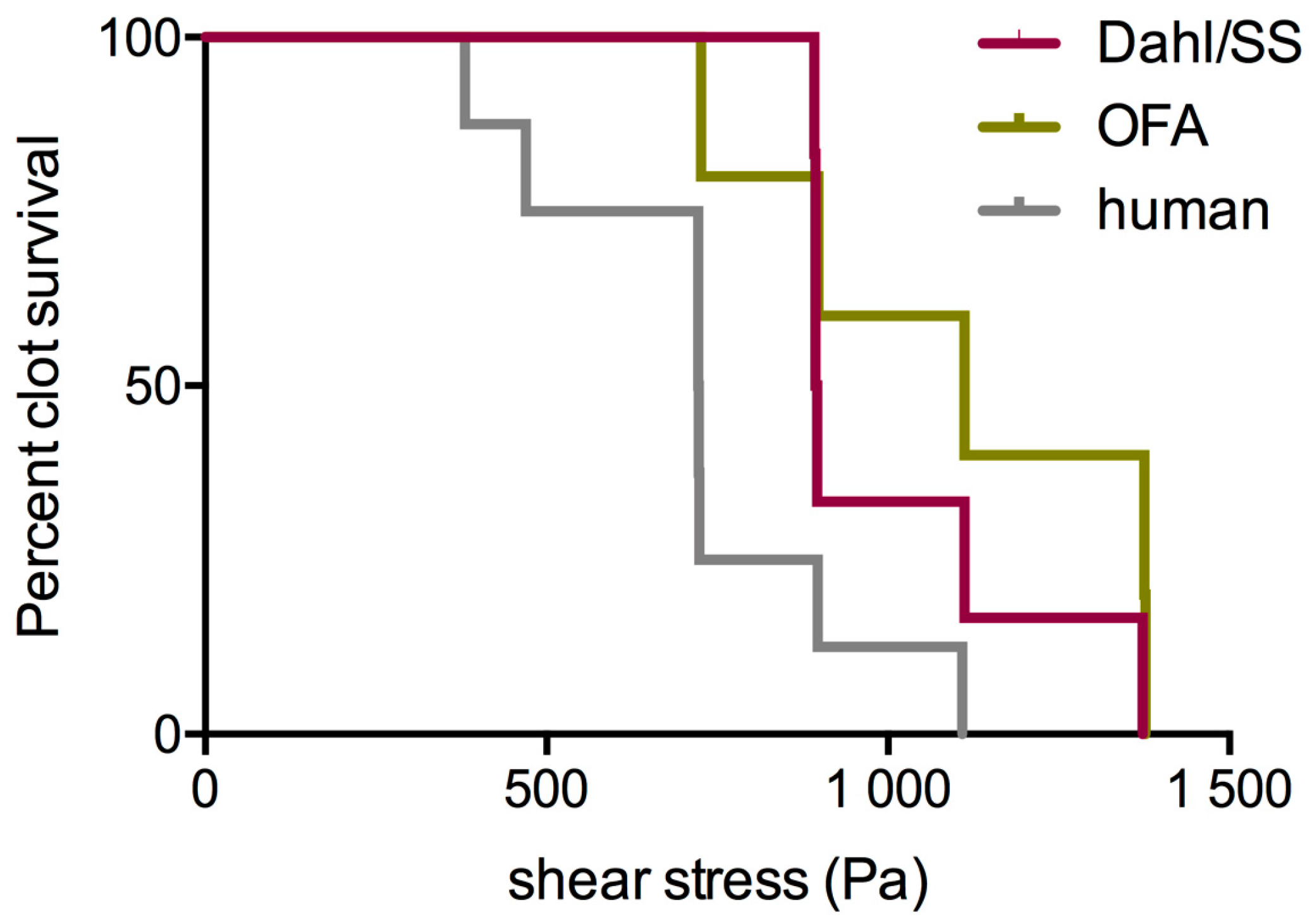
3.3. Third Test Series
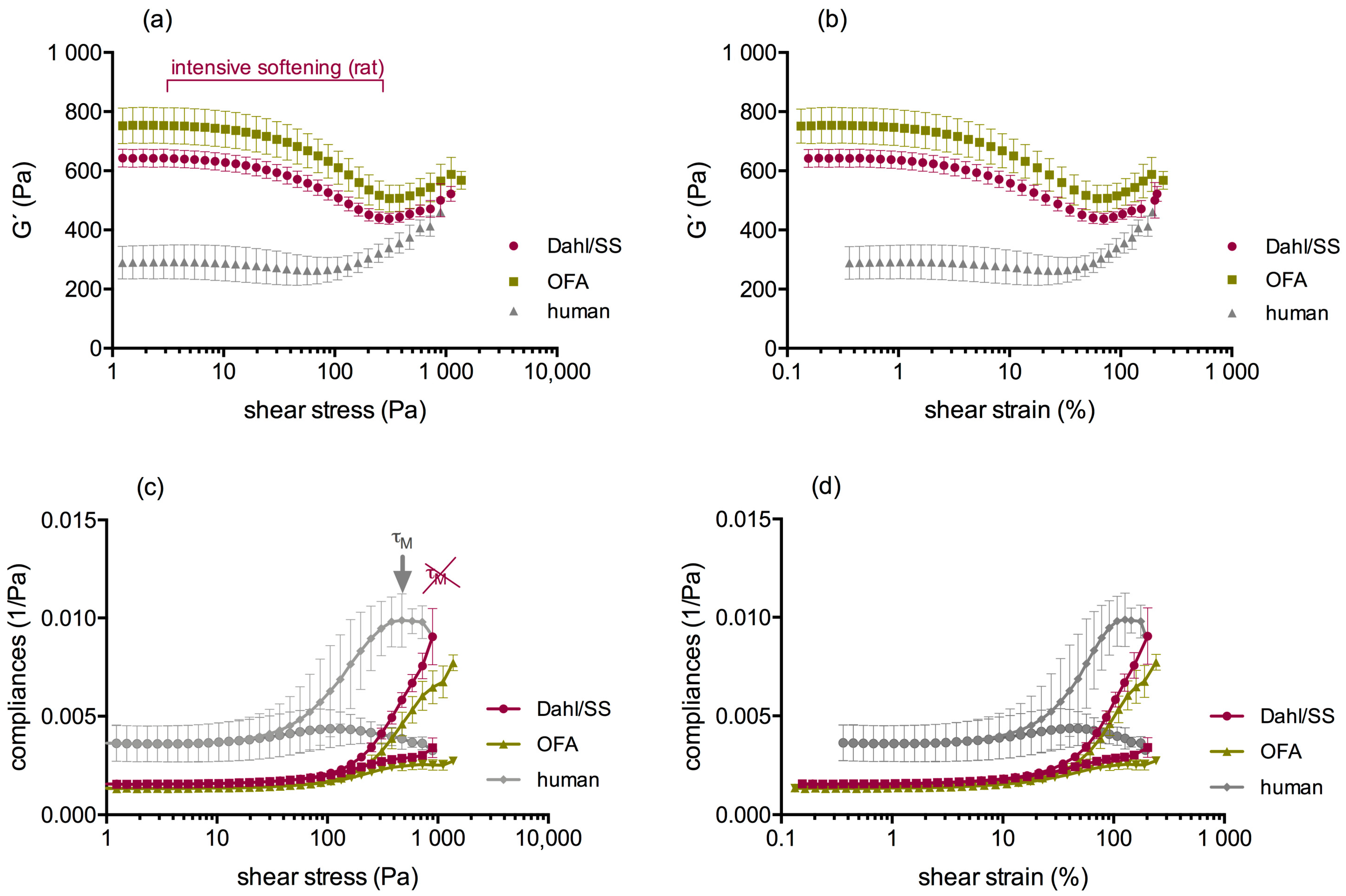
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, J.H.; Van Thiel, D.H.; Hasiba, U.; Spero, J.A.; Gavaler, J. Comparative hematology and coagulation: Studies on rodentia (Rats). Comp. Biochem. Physiol. Part A Physiol. 1985, 82, 211–215. [Google Scholar] [CrossRef]
- Gentry, P.A. Comparative aspects of blood coagulation. Vet. J. 2004, 168, 238–251. [Google Scholar] [CrossRef] [PubMed]
- García-Manzano, A.; González-Llaven, J.; Lemini, C.; Rubio-Póo, C. Standardization of rat blood clotting tests with reagents used for humans. Proc. West. Pharmacol. Soc. 2001, 44, 153–155. [Google Scholar] [PubMed]
- Siller-Matula, J.M.; Plasenzotti, R.; Spiel, A.; Quehenberger, P.; Jilma, B. Interspecies differences in coagulation profile. Thromb. Hemost. 2008, 100, 397–404. [Google Scholar] [CrossRef]
- Weber, B.; Lackner, I.; Haffner-Luntzer, M.; Palmer, A.; Pressmar, J.; Scharffetter-Kochanek, K.; Knöll, B.; Schrezenemeier, H.; Relja, B.; Kalbitz, M. Modeling trauma in rats: Similarities to humans and potential pitfalls to consider. J. Transl. Med. 2019, 17, 305. [Google Scholar] [CrossRef] [Green Version]
- Gangloff, C.; Grimault, O.; Theron, M.; Pichavant, K.; Galinat, H.; Mingant, F.; Ozier, Y. A clinically relevant and bias-controlled murine model to study acute traumatic coagulopathy. Sci. Rep. 2018, 8, 5783. [Google Scholar] [CrossRef]
- Karges, H.E.; Funk, K.A.; Ronneberger, H. Activity of coagulation and fibrinolysis parameters in animals. Arzneim. Forsch. 1994, 44, 793–797. [Google Scholar]
- Hawkey, C. Comparative Mammalian Haematology: Cellular Components and Blood Coagulation of Captive Wild Animals; Butterworth-Heinemann: London, UK, 1975. [Google Scholar]
- Muhlrad, A.; Eldor, A.; Bar-On, H.; Kahane, I. The distribution of platelet contractile proteins in normal and streptozotocin diabetic rats. Thromb. Res. 1979, 14, 621–629. [Google Scholar] [CrossRef]
- Shang, J.; Wang, M.; Feng, W.; Wu, Y.; Wu, W.; Chen, Z.; Li, Q.; Kurowski, S.; Chintala, M. Assessment of Coagulation and Platelet Activation in Rat Models of Type 2 Diabetes and Hypertension. Blood 2011, 118, 2261. [Google Scholar] [CrossRef]
- Lijnen, H.R.; Van Hoef, B.; Beelen, V.; Collen, D. Characterization of the Murine Plasma Fibrinolytic System. Eur. J. Biochem. 1994, 224, 863–871. [Google Scholar] [CrossRef]
- Sidhu, P.; Sutherland, S.B.; Ganguly, P. Interaction of thrombin with mammalian platelets. Am. J. Physiol. Heart Circ. Physiol. 1979, 6, H353–H358. [Google Scholar] [CrossRef] [PubMed]
- Derian, C.K.; Santulli, R.J.; Tomko, K.A.; Haertlein, B.J.; Andrade-Gordon, P. Species differences in platelet responses to thrombin and SFLLRN. Receptor-mediated calcium mobilization and aggregation, and regulation by protein kinases. Thromb. Res. 1995, 78, 505–519. [Google Scholar] [CrossRef]
- Yun-Choi, H.S.; Park, K.M.; Pyo, M. Epinephrine induced platelet aggregation in rat platelet-rich plasma. Thromb. Res. 2000, 100, 511–518. [Google Scholar] [CrossRef]
- Tarandovskiy, I.D.; Shin, H.K.H.; Baek, J.H.; Karnaukhova, E.; Buehler, P.W. Interspecies comparison of simultaneous thrombin and plasmin generation. Sci. Rep. 2020, 10, 3885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, E.A.; Mockros, L.F.; Weisel, J.W.; Lorand, L. Structural origins of fibrin clot rheology. Biophys. J. 1999, 77, 2813–2826. [Google Scholar] [CrossRef] [Green Version]
- Dibiasi, C.; Plewka, J.; Ploszczanski, L.; Glanz, V.; Lichtenegger, H.; Windberger, U. Viscoelasticity and structure of blood clots generated in-vitro by rheometry: A comparison between human, horse, rat, and camel. Clin. Hemorheol. Microcirc. 2018, 69, 515–531. [Google Scholar] [CrossRef]
- Cruz, M.V.; Luker, J.N.; Carney, B.C.; Brummel-Ziedins, K.E.; Bravo, M.C.; Orfeo, T.; Chen, J.H.; Moffatt, L.T.; Shupp, J.W. Reference ranges for rotational thromboelastometry in male Sprague Dawley rats. Thromb. J. 2017, 15, 31. [Google Scholar] [CrossRef]
- Weisel, J.W.; Nagaswami, C. Computer modeling of fibrin polymerization kinetics correlated with electron microscope and turbidity observations: Clot structure and assembly are kinetically controlled. Biophys. J. 1992, 63, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Parise, L.V.; Phillips, D.R. Reconstitution of the purified platelet fibrinogen receptor: Fibrinogen binding properties of the glycoprotein IIb-IIIa complex. J. Biol. Chem. 1985, 260, 10698–10707. [Google Scholar] [CrossRef]
- Windberger, U.; Läuger, J. Blood clot phenotyping by rheometry: Platelets and fibrinogen chemistry affect stress-softening and -stiffening at large oscillation amplitude. Molecules 2020, 25, 3890. [Google Scholar] [CrossRef]
- Windberger, U.; Dibiasi, C.; Lotz, E.M.; Scharbert, G.; Reinbacher-Koestinger, A.; Ivanov, I.; Ploszczanski, L.; Antonova, N.; Lichtenegger, H. The effect of hematocrit, fibrinogen concentration and temperature on the kinetics of clot formation of whole blood. Clin. Hemorheol. Microcirc. 2020, 75, 431–445. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Kim, O.V.; MacHlus, K.R.; Liu, X.; Kupaev, T.; Lioi, J.; Wolberg, A.S.; Chen, D.Z.; Rosen, E.D.; Xu, Z.; et al. Correlation between fibrin network structure and mechanical properties: An experimental and computational analysis. Soft Matter 2011, 7, 4983–4992. [Google Scholar] [CrossRef]
- Van Kempen, T.H.S.; Donders, W.P.; van de Vosse, F.N.; Peters, G.W.M. A constitutive model for developing blood clots with various compositions and their nonlinear viscoelastic behavior. Biomech. Model. Mechanobiol. 2016, 15, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Windberger, U.; Spurny, K.; Graf, A.; Thomae, H. Hemorheology in experimental research: Is it necessary to consider blood fluidity differences in the laboratory rat? Lab. Anim. 2015, 49, 142–152. [Google Scholar] [CrossRef]
- Münster, S.; Jawerth, L.M.; Leslie, B.A.; Weitz, J.I.; Fabry, B.; Weitz, D.A. Strain history dependence of the nonlinear stress response of fibrin and collagen networks. Proc. Natl. Acad. Sci. USA 2013, 110, 12197–12202. [Google Scholar] [CrossRef] [Green Version]
- Ruggeri, Z.M.; Orje, J.N.; Habermann, R.; Federici, A.B.; Reininger, A.J. Activation-independent platelet adhesion and aggregation under elevated shear stress. Blood 2006, 108, 1903–1910. [Google Scholar] [CrossRef]
- Dimitriou, C.J.; Ewoldt, R.H.; McKinley, G.H. Describing and prescribing the constitutive response of yield stress fluids using large amplitude oscillatory shear stress (LAOStress). J. Rheol. 2013, 57, 27–70. [Google Scholar] [CrossRef] [Green Version]
- Ferraro, G.; Fratini, E.; Sacco, P.; Asaro, F.; Cuomo, F.; Donati, I.; Lopez, F. Structural characterization and physical ageing of mucilage from chia for food processing applications. Food Hydrocoll. 2022, 129, 107614. [Google Scholar] [CrossRef]
- Wen, Q.; Basu, A.; Janmey, P.A.; Yodh, A.G. Non-affine deformations in polymer hydrogels. Soft Matter 2012, 8, 8039–8049. [Google Scholar] [CrossRef] [Green Version]
- Hudson, N.E.; Houser, J.R.; O’Brien, E.T.; Taylor, R.M.; Superfine, R.; Lord, S.T.; Falvo, M.R. Stiffening of individual fibrin fibers equitably distributes strain and strengthens networks. Biophys. J. 2010, 98, 1632–1640. [Google Scholar] [CrossRef] [Green Version]
- Darcourt, J.; Demchuk, A.M.; Olivot, J.-M. Platelets and Clot Stiffness. Stroke 2021, 52, 2518–2520. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.V.; Litvinov, R.I.; Alber, M.S.; Weisel, J.W. Quantitative structural mechanobiology of platelet-driven blood clot contraction. Nat. Commun. 2017, 8, 1274. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.K.; Oshinowo, O.; Ravindran, A.; Lam, W.A.; Myers, D.R. Feeling the Force: Measurements of Platelet Contraction and Their Diagnostic Implications. Semin. Thromb. Hemost. 2019, 45, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Pathare, S.J.; Eng, W.; Lee, S.-J.J.; Ramasubramanian, A.K. Fibrin prestress due to platelet aggregation and contraction increases clot stiffness. Biophys. Reports 2021, 1, 100022. [Google Scholar] [CrossRef]
- Boodt, N.; Snouckaert Van Schauburg, P.R.W.; Hund, H.M.; Fereidoonnezhad, B.; McGarry, J.P.; Akyildiz, A.C.; Van Es, A.C.G.M.; De Meyer, S.F.; Dippel, D.W.J.; Lingsma, H.F.; et al. Mechanical Characterization of Thrombi Retrieved with Endovascular Thrombectomy in Patients with Acute Ischemic Stroke. Stroke 2021, 52, 2510–2517. [Google Scholar] [CrossRef]
- Weisel, J.W.; Litvinov, R.I. Red blood cells: The forgotten player in hemostasis and thrombosis. J. Thromb. Haemost. 2019, 17, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Tutwiler, V.; Wang, H.; Litvinov, R.I.; Weisel, J.W.; Shenoy, V.B. Interplay of Platelet Contractility and Elasticity of Fibrin/Erythrocytes in Blood Clot Retraction. Biophys. J. 2017, 112, 714–723. [Google Scholar] [CrossRef] [Green Version]
- Falvo, M.R.; Gorkun, O.V.; Lord, S. The molecular origins of the mechanical properties of fibrin. Biophys. Chem. 2010, 152, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Doolittle, R.F.; Kollman, J.M. Natively unfolded regions of the vertebrate fibrinogen molecule. Proteins Struct. Funct. Genet. 2006, 63, 391–397. [Google Scholar] [CrossRef]
- Falvo, M.R.; Millard, D.; O´Brian, T.; Superfine, R.; Lord, S. Length of Tandem Repeats in Fibrin’s αC Region Correlates with Fiber Extensibility. J. Thromb. Haemost. 2008, 6, 1991–1993. [Google Scholar] [CrossRef]
- Hochleitner, G.; Sutor, K.; Levett, C.; Leyser, H.; Schlimp, C.J.; Solomon, C. Revisiting Hartert’s 1962 Calculation of the Physical Constants of Thrombelastography. Clin. Appl. Thromb. 2017, 23, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badiei, N.; Sowedan, A.M.; Curtis, D.J.; Brown, M.R.; Lawrence, M.J.; Campbell, A.I.; Sabra, A.; Evans, P.A.; Weisel, J.W.; Chernysh, I.N.; et al. Effects of unidirectional flow shear stresses on the formation, fractal microstructure and rigidity of incipient whole blood clots and fibrin gels. Clin. Hemorheol. Microcirc. 2015, 60, 451–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.; Ramey, E.; Ramwell, P.W. Androgen mediated sensitivity in platelet aggregation. Am. J. Physiol. 1977, 232, H381–H385. [Google Scholar] [CrossRef] [PubMed]
- Chernysh, I.N.; Nagaswami, C.; Kosolapova, S.; Peshkova, A.D.; Cuker, A.; Cines, D.B.; Cambor, C.L.; Litvinov, R.I.; Weisel, J.W. The distinctive structure and composition of arterial and venous thrombi and pulmonary emboli. Sci. Rep. 2020, 10, 5112. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Windberger, U.; Glanz, V.; Ploszczanski, L. Laboratory Rat Thrombi Lose One-Third of Their Stiffness When Exposed to Large Oscillating Shear Stress Amplitudes: Contrasting Behavior to Human Clots. Int. J. Transl. Med. 2022, 2, 332-344. https://doi.org/10.3390/ijtm2030026
Windberger U, Glanz V, Ploszczanski L. Laboratory Rat Thrombi Lose One-Third of Their Stiffness When Exposed to Large Oscillating Shear Stress Amplitudes: Contrasting Behavior to Human Clots. International Journal of Translational Medicine. 2022; 2(3):332-344. https://doi.org/10.3390/ijtm2030026
Chicago/Turabian StyleWindberger, Ursula, Veronika Glanz, and Leon Ploszczanski. 2022. "Laboratory Rat Thrombi Lose One-Third of Their Stiffness When Exposed to Large Oscillating Shear Stress Amplitudes: Contrasting Behavior to Human Clots" International Journal of Translational Medicine 2, no. 3: 332-344. https://doi.org/10.3390/ijtm2030026
APA StyleWindberger, U., Glanz, V., & Ploszczanski, L. (2022). Laboratory Rat Thrombi Lose One-Third of Their Stiffness When Exposed to Large Oscillating Shear Stress Amplitudes: Contrasting Behavior to Human Clots. International Journal of Translational Medicine, 2(3), 332-344. https://doi.org/10.3390/ijtm2030026