1. Introduction
Global fishmeal production is approximately 5 million tons, but increasing demand for aquafeeds, competitiveness and rising prices have limited its availability [
1]. To cover the limited availability of fishmeal for aquafeeds, the increasing use of plant protein sources such as pea [
2], soybean [
3,
4], lupine [
5], corn gluten [
4,
5,
6], canola/rapeseed [
7] and cotton seed [
8] meals has become a global trend [
2]. The global crop production is estimated to be over 9 billion tons, and according to the United States Department of Agriculture (USDA), the world rice production in 2021/2022 is estimated to reach 513.02 million metric tons [
9], becoming the third most produced crop after maze and wheat [
9]. Because rice is produced for human food, its safety is guaranteed.
Although the protein content of rice is only 6.3–7.1%, rice protein concentrate (RPC), a by-product of rice starch production, contains more than 65%. In the general manufacturing process of rice starch, polished rice is dissolved in an alkaline solution, and a starch fraction is collected after centrifugation. Rice protein dissolved in a supernatant can be isolated after neutralization [
10].
Although RPC is commonly used in the animal feed industry [
11,
12], there are limited reports on the use of RPC as a fishmeal alternative in aquafeeds. Soybean meal and corn gluten meal are the most popular plant protein alternatives to fishmeal, and soybean and corn prices are closely related because they are commonly affected by similar factors such as inflation, crude oil price, and production [
12]. On the other hand, the association between rice and other crop prices is low [
12]. Thus, the replacement of soybean and corn gluten meal with rice by-products could contribute to the price stability of the diet. RPC has been tested as a protein source for the black spot bream (
Pagellus bogaraveo), blunt snout bream (
Megalobrama amblycephala), Nile tilapia (
Oreochromis niloticus), rainbow trout (
O. mykiss), Siberian sturgeon (
Acipenser baerii), hybrid sturgeon (
Acipenser naccarii ×
Acipenser baeri) and Pacific whiteleg shrimp (
Litopenaeus vanamei) [
6,
13,
14,
15,
16,
17,
18]. Palmegiano et al. (2006) tested the usefulness of RPC in the diet of rainbow trout [
13] and concluded that 20% RPC could be included in the diet of rainbow trout without a negative impact on fish growth [
13]. However, their diet contains 35% fishmeal. Normally, the fishmeal inclusion level in an aquafeed determines the acceptable upper limit of the inclusion level of alternative protein sources in a diet. Thus, it is not clear whether 20% RPC can be included without a negative impact on the growth of the rainbow trout fed a diet containing less than 35% of fishmeal. In addition, there were no reports on the replacement of soybean and corn gluten meal with RPC in a rainbow trout diet.
Enzyme treatment and fermentation are popular techniques for improving the performance of plant protein-based diets for fish [
19].
Aspelgillus niger is naturally isolated from rice fermentation and widely used for the production of rice wine and soy sauce [
20,
21]. Thus, the enzyme complex produced from
A. niger seems to effectively digest rice protein. However, there have been no reports on the treatment of RPC by the enzyme complex. In addition, one concern regarding an enzyme complex is the lower efficacy of an enzyme complex when it is a supplement in the diet for cold-water species because an enzyme complex may not be effective below 18 °C; therefore, a simple addition of an enzyme complex to a diet for cold-water species seems to be less effective. However, we hypothesized that the pretreatment of plant protein sources with an enzyme complex at an optimal temperature could improve the feed performance of cold-water species. Therefore, we pretreated RPC with the enzyme complex at 45 °C for 18 h before formulating an extruded diet. The objective of this study was to investigate the effect of a diet containing 5–15% of an enzyme complex-pretreated RPC on growth performance, nutrient digestibility and nutrient retention in juvenile rainbow trout.
4. Discussion
Gaylord et al. (2010) and Gaylord and Barrows (2008) examined the gross nutrient and amino acid availability in rainbow trout from 25 feed ingredients in an extruded diet [
29,
30] and demonstrated low digestibility of crude protein, and low amino acid availability from RPC among the 16 plant ingredients examined [
29,
30], suggests a relatively low availability of RPC as a protein source for an extruded diet for rainbow trout. There were no significant differences in any of the growth parameters examined in rainbow trout. Although no significant difference was observed in fish-fed diets formulated with 15% RPC, a marked decrease was observed in the final weight, weight gain and SGR in this group. This also suggests that the inclusion of 15% RPC negatively affects fish growth. Considering previous experiments on the rainbow trout testing effects of RPC in a diet, it has been observed that with an increase in the percentage of RPC, the growth performance of fish is negatively affected [
13]. A significant decrease in growth performance was observed in Pacific whiteleg shrimp as the dietary RPC level increased in the experimental diets [
16]. It can be hypothesized that by increasing the RPC level in the diet of fish, the growth performance may be adversely affected. On the contrary, no negative impact was reported on the growth performance of the tilapia-fed diet, replacing all fishmeal in the diet with RPC [
15]. Additionally, a lysine-supplemented RPC-based non-fishmeal in the diet did not show adverse effects on the growth of blunt snout bream [
14]. This could be explained by the different feeding habits of the species; tilapia and blunt snout bream are omnihervivorous species and accept a relatively wider range of feedstuffs than carnivorous species such as rainbow trout. In addition, they observed a higher feed intake of fish fed a higher inclusion level of RPC than that of fish fed a fishmeal diet [
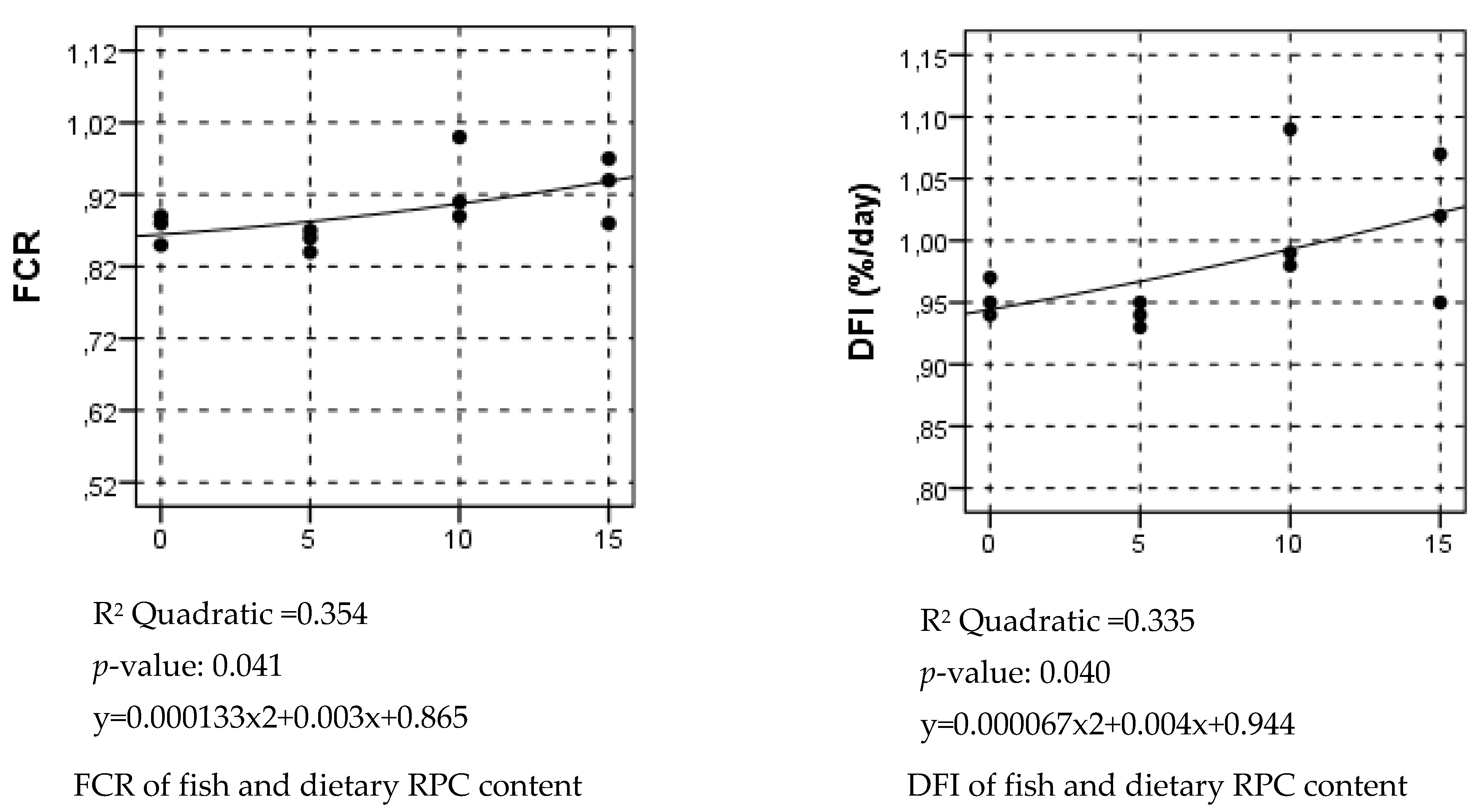
15]. However, such a higher feed intake was not observed in the rainbow trout in the present study. We thought that there were two possible explanations for the reasons for decreased feed intake; (1) an increasing level of RPC in the diet and (2) decreasing level of soybean and corn gluten meal accompanied by an increasing RPC level in the diet. Unfortunately, it was not clear whether RPC or soybean and corn gluten meal were more palatable for rainbow trout in the present study. Considering the greater negative impact of dietary RPC on DFI than FCR, it was inferred that one of the reasons for the decreased FCR was the decreased diet consumption.
In this experiment, although no significant difference in lipid digestibility was observed in any of the groups, a significant decrease in phosphorus digestibility was observed in the RPC5E group. Soybean, corn and rice contain phytate-phosphorous [
31], which is not utilized as a phosphorous source in monogastric animals, including fish, without the use of exogenous phytase; however, in this study, we used an enzyme complex containing phytase. Therefore, phytate phosphorous in our diet can be a useful source of phosphorous. The phosphorus sources in our diet were monobasic calcium phosphate, fishmeal, soybean meal, corn gluten meal and RPC. Because we included the same level of fishmeal and monobasic calcium phosphate commonly in all diets, they could not be the cause of lower phosphorous availability in RPC15E. Different concentrations of soybean meal, corn gluten meal and RPC were formulated in RPC diets and treated with enzyme complexes before extrusion processing. The efficacy of phytase in liberating free phosphorous from different plant ingredients could differ depending on the feed formulation [
32]. The presence of higher levels of soybean meal and corn gluten meal in the RPC15E diet could be one of the causes of lower phosphorous absorption in the RPC5E group. The growth performance of the 10E and 15E groups could be suppressed, in part, due to the lower protein digestibility in the diet in our study. Rice contains serine protease inhibitors as well as oryzacystatin, another protease inhibitor [
32,
33]. These antinutritional factors limit protein digestibility and availability of RPC in a diet with a lower fishmeal percentage. However, the inclusion of a high percentage of fishmeal in a basal diet may mask the negative impact of protease inhibitors in RPC in rainbow trout diets. This idea is supported by the fact that no negative effect of RPC on the growth performance of rainbow trout was observed when it is formulated with 35% fishmeal in an extruded diet [
13]. Considering the optimal RPC level in an extruded diet for rainbow trout, 20% RPC can be included when the fishmeal content is 35% [
13]. Here, we also estimated that the optimal RPC level in the extruded diet for rainbow trout was approximately 5% when the dietary fishmeal level was 20%. Considering the differences in RPC and fishmeal content in the diet in the previous and present studies, when a fishmeal content is 15% higher than that of the RPC level in the diet, it does not seem to affect the growth of rainbow trout negatively.
Protein in rice is a seed storage protein called glutelin or prolamin and has a role in storing carbon, nitrogen and sulfur until it is used for seed germination [
34]. Rice protein in milled rice is composed of 5–8% prolamin, 15% albumin and globulin and approximately 80% glutelin [
11]. Seed storage proteins are hydrolyzed and utilized during germination but are protected by protease inhibitors until germination initiation [
34]. Therefore, when rice proteins are isolated and concentrated, these protease inhibitors are included together with prolamin and glutelin. Therefore, it is considered that the protease inhibitor contained is a contaminant in the RPC, thus reducing the digestion and absorption rate of the RPC protein.
The major focus of fishmeal replacement studies is monitoring growth performance, probably because it is directly associated with economic aspects. However, relatively few studies have reported its impact on the nutrient content of fish-fed alternative protein sources. It was reported that the inclusion of more than 20% RPC in the trout diet reduces nutrient digestibility and thus leads to lower growth [
13]. Here, we observed significantly lower lipid content in fish fed more than 10% RPC in the diet of the rainbow trout. This was also reflected in the low lipid retention of the fish. Lower lipid digestibility was observed in rainbow trout fed a diet containing more than 20% RPC. The reason our result did not match with those of the previous study could be explained by a different inclusion level of RPC; 5–15% in the diet used in the present diet vs. 20–53% RPC in the diet used by Palmegiano et al. [
13]. Higher levels of RPC in the diet had a negative impact on lipid digestibility in rainbow trout. Another explanation is that the diet in the present study contained RPC pretreated with enzyme complex. Enzyme pretreatment of RPC may partially reduce the negative impact of the antinutritional factors on lipid digestibility. It was also reported that protein digestion of the fish fed the RPC-based diet decreased linearly in rainbow trout [
13]. Our study also observed a negative impact of RPC on protein digestion. As previously discussed, the existence of a protease inhibitor has been suggested in RPC [
33,
34]. In addition, prolamin, one of the major components of the RPC, is resistant to pepsin digestion [
35]. The inclusion of protease inhibitors and the digestion-resistant nature of the protein component of RPC is considered to lead to lower protein digestion in fish fed the RPC-formulated diet in our study.


 ,
, 

{kind=link}