
The Kelch/Nrf2 Antioxidant System as a Target for Some Marine Fungal Metabolites



Abstract
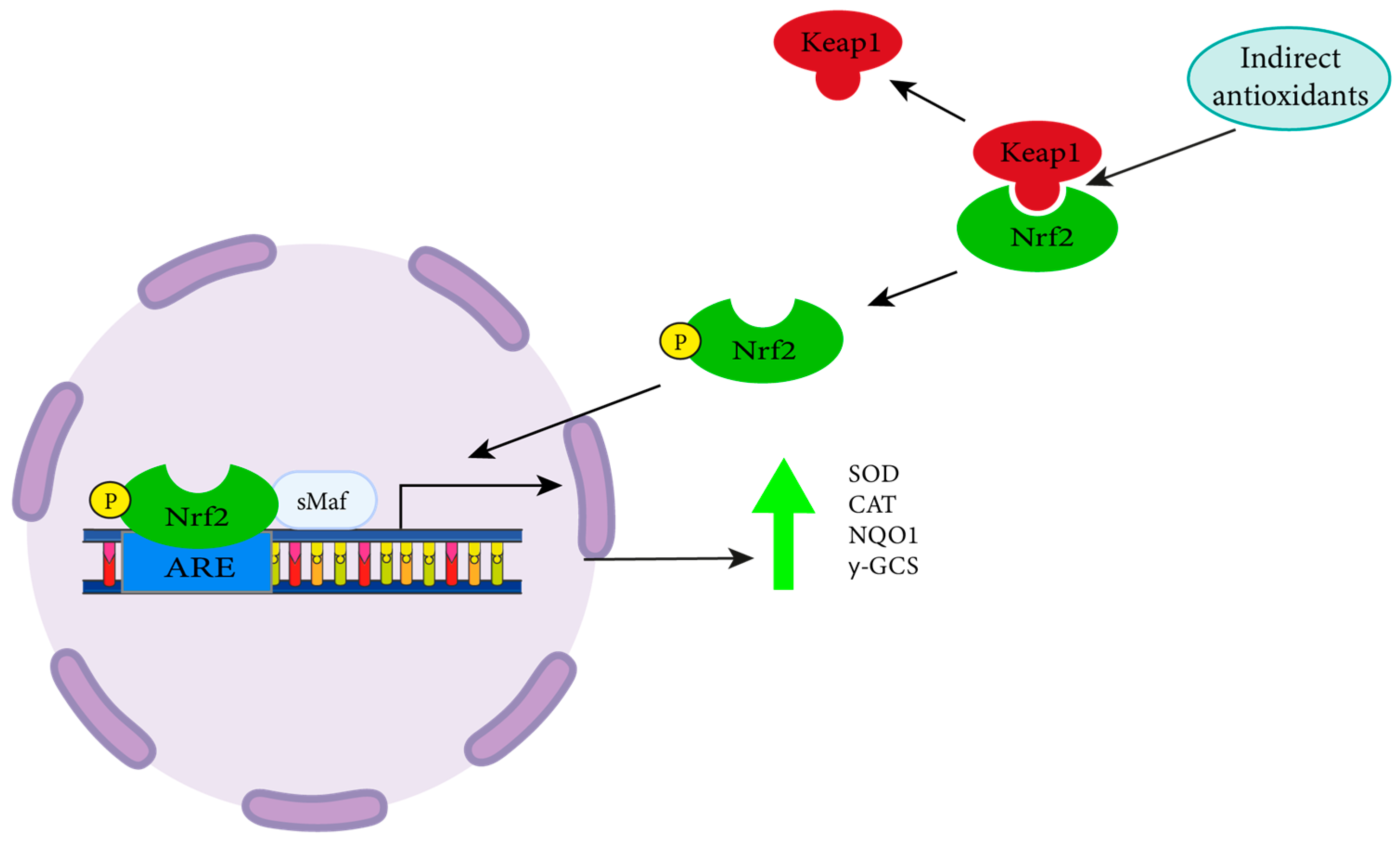
:1. Introduction
2. Materials and Methods
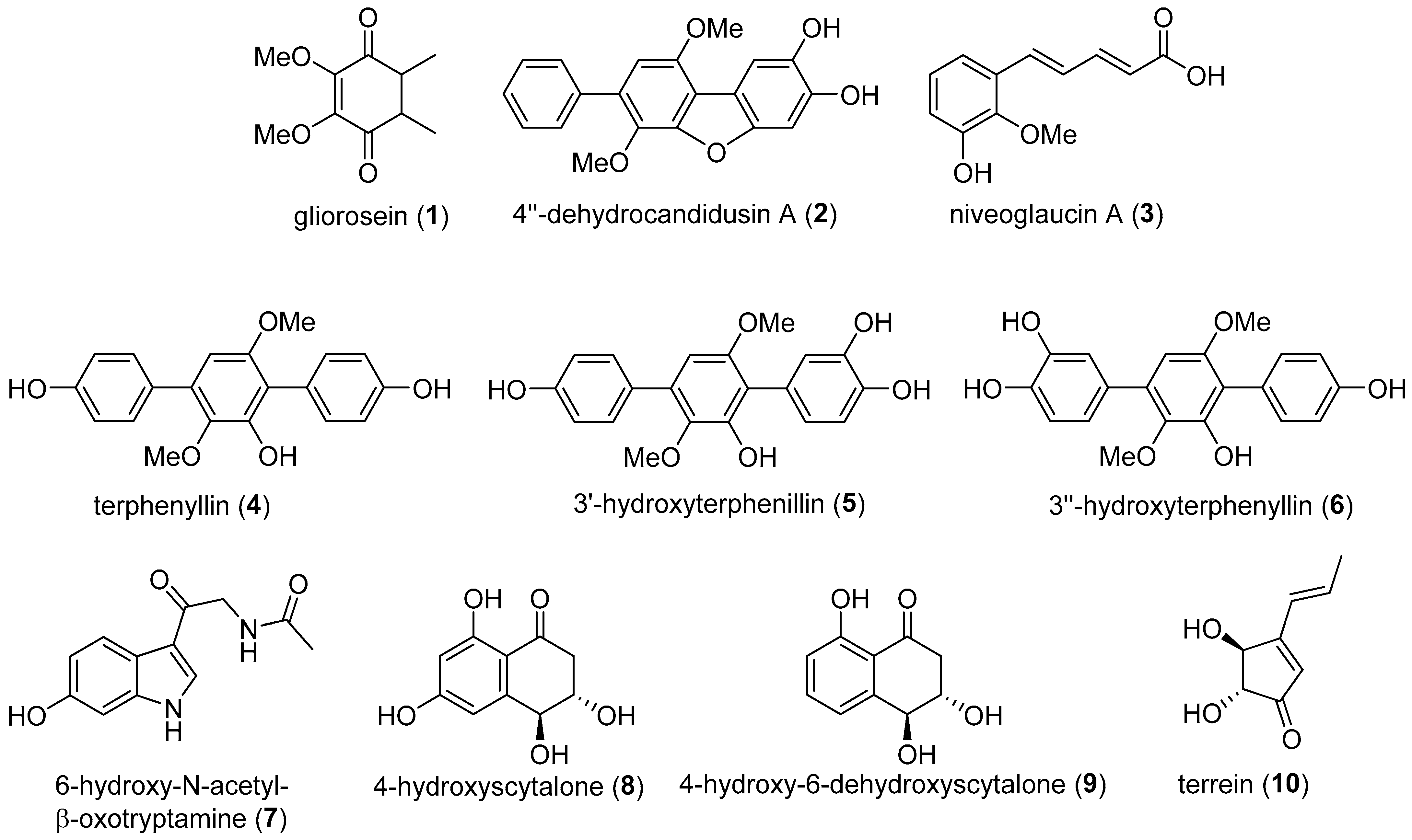
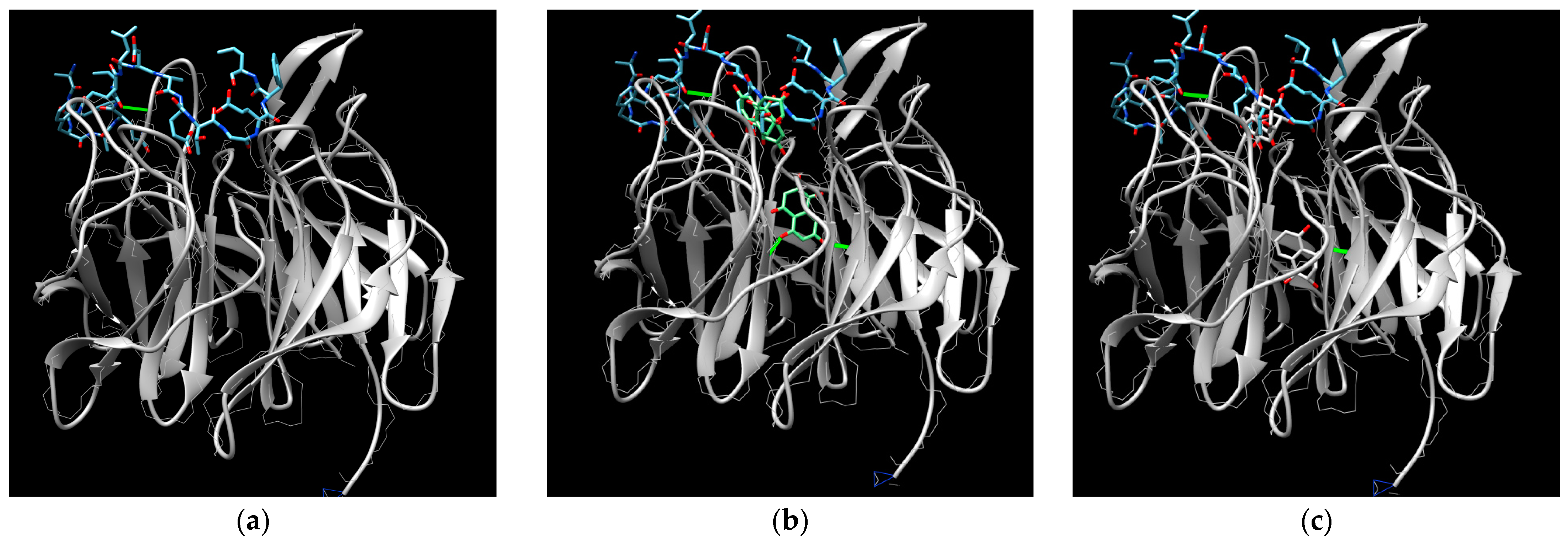
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Role of Nrf2 in Oxidative Stress and Toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef]
- Canning, P.; Sorrell, F.J.; Bullock, A.N. Structural basis of Keap1 interactions with Nrf2. Free Radic. Biol. Med. 2015, 88, 101–107. [Google Scholar] [CrossRef]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef]
- Jung, K.-A.; Kwak, M.-K. The Nrf2 System as a Potential Target for the Development of Indirect Antioxidants. Molecules 2010, 15, 7266–7291. [Google Scholar] [CrossRef]
- Kobayashi, M.; Li, L.; Iwamoto, N.; Nakajima-Takagi, Y.; Kaneko, H.; Nakayama, Y.; Eguchi, M.; Wada, Y.; Kumagai, Y.; Yamamoto, M. The Antioxidant Defense System Keap1-Nrf2 Comprises a Multiple Sensing Mechanism for Responding to a Wide Range of Chemical Compounds. Mol. Cell. Biol. 2009, 29, 493–502. [Google Scholar] [CrossRef]
- Vriend, J.; Reiter, R.J. The Keap1-Nrf2-antioxidant response element pathway: A review of its regulation by melatonin and the proteasome. Mol. Cell. Endocrinol. 2015, 401, 213–220. [Google Scholar] [CrossRef]
- Zhou, Y.; Jiang, Z.; Lu, H.; Xu, Z.; Tong, R.; Shi, J.; Jia, G. Recent Advances of Natural Polyphenols Activators for Keap1-Nrf2 Signaling Pathway. Chem. Biodivers. 2019, 16, e1900400. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Barber, A.J.; Spagnuolo, C.; Russo, G.L.; Daglia, M.; Nabavi, S.M.; Sobarzo-Sánchez, E. Nrf2 as molecular target for polyphenols: A novel therapeutic strategy in diabetic retinopathy. Crit. Rev. Clin. Lab. Sci. 2016, 53, 293–312. [Google Scholar] [CrossRef]
- Kohandel, Z.; Farkhondeh, T.; Aschner, M.; Samarghandian, S. Nrf2 a molecular therapeutic target for Astaxanthin. Biomed. Pharmacother. 2021, 137, 111374. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, Y.; You, Q.; Jiang, Z. Recent progress in the development of small molecule Nrf2 activators: A patent review (2017-present). Expert Opin. Ther. Pat. 2020, 30, 209–225. [Google Scholar] [CrossRef]
- Klomparens, E.A.; Ding, Y. The neuroprotective mechanisms and effects of sulforaphane. Brain Circ. 2019, 5, 74–83. [Google Scholar]
- Cleasby, A.; Yon, J.; Day, P.J.; Richardson, C.; Tickle, I.J.; Williams, P.A.; Callahan, J.F.; Carr, R.; Concha, N.; Kerns, J.K. Structure of the BTB domain of Keap1 and its interaction with the triterpenoid antagonist CDDO. PLoS ONE 2014, 9, e98896. [Google Scholar] [CrossRef]
- Zhuang, C.; Miao, Z.; Sheng, C.; Zhang, W. Updated research and applications of small molecule inhibitors of Keap1-Nrf2 protein-protein interaction: A review. Curr. Med. Chem. 2014, 21, 1861–1870. [Google Scholar] [CrossRef]
- Li, L.; Liu, J.; Nie, S.; Ding, L.; Wang, L.; Liu, J.; Liu, W.; Zhang, T. Direct inhibition of Keap1–Nrf2 interaction by egg-derived peptides DKK and DDW revealed by molecular docking and fluorescence polarization. RSC Adv. 2017, 7, 34963–34971. [Google Scholar] [CrossRef]
- Vitale, G.A.; Coppola, D.; Palma Esposito, F.; Buonocore, C.; Ausuri, J.; Tortorella, E.; de Pascale, D. Antioxidant Molecules from Marine Fungi: Methodologies and Perspectives. Antioxidants 2020, 9, 1183. [Google Scholar] [CrossRef]
- Kim, K.W.; Kim, H.J.; Sohn, J.H.; Yim, J.H.; Kim, Y.C.; Oh, H. Terrein suppressed lipopolysaccharide-induced neuroinflammation through inhibition of NF-κB pathway by activating Nrf2/HO-1 signaling in BV2 and primary microglial cells. J. Pharmacol. Sci. 2020, 143, 209–218. [Google Scholar] [CrossRef]
- Yang, Z.; Zhu, M.L.; Li, D.H.; Zeng, R.; Han, B.N. N-Me-trichodermamide B isolated from Penicillium janthinellum, with antioxidant properties through Nrf2-mediated signaling pathway. Bioorg. Med. Chem. 2017, 25, 6614–6622. [Google Scholar] [CrossRef]
- Trinh, P.T.; Yurchenko, A.N.; Khmel, O.O.; Dieu, T.V.; Ngoc, N.T.; Girich, E.V.; Menshov, A.S.; Kim, N.Y.; Chingizova, E.A.; Van, T.T.; et al. Cytoprotective Polyketides from Sponge-Derived Fungus Lopadostoma pouzarii. Molecules 2022, 27, 7650. [Google Scholar] [CrossRef]
- Yurchenko, E.A.; Menchinskaya, E.S.; Pislyagin, E.A.; Chingizova, E.A.; Girich, E.V.; Yurchenko, A.N.; Aminin, D.L.; Mikhailov, V.V. Cytoprotective Activity of p-Terphenyl Polyketides and Flavuside B from Marine-Derived Fungi against Oxidative Stress in Neuro-2a Cells. Molecules 2021, 26, 3618. [Google Scholar] [CrossRef]
- Yurchenko, A.N.; Smetanina, O.F.; Ivanets, E.V.; Phan, T.T.H.; Ngo, N.T.D.; Zhuravleva, O.I.; Rasin, A.B.; Dyshlovoy, S.A.; Menchinskaya, E.S.; Pislyagin, E.A.; et al. Auroglaucin-related neuroprotective compounds from Vietnamese marine sediment-derived fungus Aspergillus niveoglaucus. Nat. Prod. Res. 2020, 34, 2589–2594. [Google Scholar] [CrossRef] [PubMed]
- Girich, E.V.; Yurchenko, A.N.; Smetanina, O.F.; Trinh, P.T.H.; Ngoc, N.T.D.; Pivkin, M.V.; Popov, R.S.; Pislyagin, E.A.; Menchinskaya, E.S.; Chingizova, E.A.; et al. Neuroprotective Metabolites from Vietnamese Marine Derived Fungi of Aspergillus and Penicillium Genera. Mar. Drugs 2020, 18, 608. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, E.; Menchinskaya, E.; Pislyagin, E.; Trinh, P.; Ivanets, E.; Smetanina, O.; Yurchenko, A. Neuroprotective Activity of Some Marine Fungal Metabolites in the 6-Hydroxydopamin- and Paraquat-Induced Parkinson’s Disease Models. Mar. Drugs 2018, 16, 457. [Google Scholar] [CrossRef] [PubMed]
- Grosdidier, A.; Zoete, V.; Michielin, O. SwissDock, a protein-small molecule docking web service based on EADock DSS. Nucleic Acids Res. 2011, 39, W270–W277. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Brooks Iii, C.L.; Mackerell Jr, A.D.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef]
- Haberthür, U.; Caflisch, A. FACTS: Fast analytical continuum treatment of solvation. J. Comput. Chem. 2008, 29, 701–715. [Google Scholar] [CrossRef]
- Grosdidier, A.; Zoete, V.; Michielin, O. Fast docking using the CHARMM force field with EADock DSS. J. Comput. Chem. 2011, 32, 2149–2159. [Google Scholar] [CrossRef]
- Grosdidier, A.; Zoete, V.; Michielin, O. EADock: Docking of small molecules into protein active sites with a multiobjective evolutionary optimization. Proteins Struct. Funct. Bioinform. 2007, 67, 1010–1025. [Google Scholar] [CrossRef]
- Cianciullo, P.; Maresca, V.; Sorbo, S.; Basile, A. Antioxidant and Antibacterial Properties of Extracts and Bioactive Compounds in Bryophytes. Appl. Sci. 2022, 12, 160. [Google Scholar] [CrossRef]
- Jnoff, E.; Albrecht, C.; Barker, J.J.; Barker, O.; Beaumont, E.; Bromidge, S.; Brookfield, F.; Brooks, M.; Bubert, C.; Ceska, T. Binding mode and structure–activity relationships around direct inhibitors of the Nrf2–Keap1 complex. ChemMedChem 2014, 9, 699–705. [Google Scholar] [CrossRef]
- Herrera-Arozamena, C.; Estrada-Valencia, M.; Pérez, C.; Lagartera, L.; Morales-García, J.A.; Pérez-Castillo, A.; Franco-Gonzalez, J.F.; Michalska, P.; Duarte, P.; León, R.; et al. Tuning melatonin receptor subtype selectivity in oxadiazolone-based analogues: Discovery of QR2 ligands and NRF2 activators with neurogenic properties. Eur. J. Med. Chem. 2020, 190, 112090. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, L.-Q.; Long, H.-P.; Liu, J.; Jiang, Y.-P.; Xue, Y.; Wang, W.-X.; Tan, G.-S.; Gong, Z.-C.; Liu, J.-K. Xylarinaps A–E, five pairs of naphthalenone derivatives with neuroprotective activities from Xylaria nigripes. Phytochemistry 2021, 186, 112729. [Google Scholar] [CrossRef] [PubMed]
- Inoyama, D.; Chen, Y.; Huang, X.; Beamer, L.J.; Kong, A.-N.T.; Hu, L. Optimization of fluorescently labeled Nrf2 peptide probes and the development of a fluorescence polarization assay for the discovery of inhibitors of Keap1-Nrf2 interaction. J. Biomol. Screen. 2012, 17, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Inoyama, D.; Kong, A.-N.T.; Beamer, L.J.; Hu, L. Kinetic Analyses of Keap1–Nrf2 Interaction and Determination of the Minimal Nrf2 Peptide Sequence Required for Keap1 Binding Using Surface Plasmon Resonance. Chem. Biol. Drug Des. 2011, 78, 1014–1021. [Google Scholar] [CrossRef]
- Smirnova, N.A.; Haskew-Layton, R.E.; Basso, M.; Hushpulian, D.M.; Payappilly, J.B.; Speer, R.E.; Ahn, Y.-H.; Rakhman, I.; Cole, P.A.; Pinto, J.T. Development of Neh2-luciferase reporter and its application for high throughput screening and real-time monitoring of Nrf2 activators. Chem. Biol. 2011, 18, 752–765. [Google Scholar] [CrossRef]
- Cuadrado, A.; Rojo, A.I.; Wells, G.; Hayes, J.D.; Cousin, S.P.; Rumsey, W.L.; Attucks, O.C.; Franklin, S.; Levonen, A.-L.; Kensler, T.W. Therapeutic targeting of the NRF2 and KEAP1 partnership in chronic diseases. Nat. Rev. Drug Discov. 2019, 18, 295–317. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ΔG (kcal/mol) | Full Fitness (kcal/mol) | H-Binding Residue/ H-Bonding Distance, A° |
|---|---|---|
| −14.760137 | −900.57 | Arg 415 ⟷ O23/2.674 |
| O19 ⟷ Arg 415/2.682 | ||
| Arg 415 ⟷ O23/2.068 | ||
| Arg 415 ⟷ O19/2.072 | ||
| Arg 483 ⟷ O14/2.386 | ||
| O5 ⟷ Ser 439/2.445 | ||
| Arg 459 ⟷ O3/1.835 | ||
| O6 ⟷ Hsd 436/2.105 | ||
| O8 ⟷ Thr 481/2.021 |
| Compound | Cluster | ΔG (kcal/mol) | Full Fitness (kcal/mol) | H-Binding Residue/ H-Bonding Distance, A° | Location in Nrf2-Binding Site of Keap1 |
|---|---|---|---|---|---|
| Gliorosein (1) | 2 | −6.909368 | −1111.3994 | Val606 ⟷ O/2.049 Val418 ⟷ O1/2.309 Val465 ⟷ O1/2.543 | no |
| 1 | −6.7755146 | −1112.5961 | Arg415 ⟷ O/2.004 Arg415 ⟷ O/2.497 | yes | |
| 4″-Hydroxycandidusine A (2) | 26 | −8.116934 | −1084.1508 | Val606 ⟷ O4/2.123 H8 ⟷ Val463/2.382 Val465 ⟷ O3/2.215 | no |
| 2 | −7.9192557 | −1090.6539 | Val606 ⟷ O4/2.486 Val465 ⟷ O3/2.202 | no | |
| Niveoglaucin A (3) | 4 | −7.22126 | −1138.7821 | H3 ⟷ Ala510/1.975 H8 ⟷ Val418/1.894 Val418 ⟷ O2/2.080 | no |
| 3 | −6.7057815 | −1139.0631 | Arg415 ⟷ O2/2.704 H3 ⟷ Leu365/2.164 | yes | |
| 13 | −6.941605 | −6.941605 | H3 ⟷ Ser363/2.201 | yes | |
| Terphenyllin (4) | 18 | −6.979565 | −1051.2461 | Val418 ⟷ O3/1.976 Gly367 ⟷ O2/1.940 H11 ⟷ Gly367/2.021 Val606 ⟷ O2/2.176 Val606 ⟷ O4/2.621 | no |
| 0 | −7.871525 | −1079.2283 | Val465 ⟷ O2/2.322 H11 ⟷ Val465/1.927 | no | |
| 3′-Hydroxyterphenyllin (5) | 6 | −8.2964945 | −1072.0981 | H10 ⟷ Val418/1.719 Val606 ⟷ O4/2.171 Val465 ⟷ O3/2.433 | no |
| 0 | −7.9774666 | −1076.5175 | H11 ⟷ Val606/1.803 Val465 ⟷ O4/2.394 H10 ⟷ Leu365/2.110 Val606 ⟷ O3/2.478 | no | |
| 3″-Hydroxyterphenyllin (6) | 0 | −7.9407578 | −1075.1831 | H8 ⟷ Val463/2.042 H10 ⟷ Ile559/2.486 Thr560 ⟷O2/2.469 | no |
| 26 | −7.0460024 | −1068.861 | Thr560 ⟷ O1/2.321 H9 ⟷ Val606/1.950 Val369 ⟷ O2/2.526 Gly423 ⟷ O3/2.232 Asp422 ⟷ O/2.324 | no | |
| 6-Hydroxy-N-acetyl-β-oxotryptamin (7) | 1 | −7.4022284 | −1135.5000 | H5 ⟷ Val606/1.901 H3 ⟷ Val512/1.949 | no |
| 23 | −6.5958056 | −1131.7983 | Arg483 ⟷ O2/2.564 Arg415 ⟷ O1/2.703 | Yes | |
| 4-Hydroxyscytalone (8) | 1 | −7.197147 | −1133.7689 | Val606 ⟷ O3/2.464 Val512 ⟷ O2/2.414 H9 ⟷ Val604/2.132 | no |
| 5 | −6.555147 | −1132.1863 | Arg483 ⟷ O3/2.294 H6 ⟷ Ser555/2.154 | yes | |
| 12 | −6.406861 | −1133.5903 | Arg483 ⟷ O1/2.400 Arg483 ⟷ O2/2.801 H9 ⟷ Ser555/2.364 Gln530 ⟷ O4/2.445 | yes | |
| 4-Hydroxy-6-dehydroxyscytalone (9) | 15 | −7.199379 | −1114.5966 | H7 ⟷ Gly367/2.664 Gly367 ⟷ O1/2.155 Val606 ⟷ O1/2.084 H9 ⟷ Val465/2.201 H7 ⟷ Val606/1.114 | no |
| 7 | −6.592193 | −1121.9755 | no | yes | |
| 21 | −6.27284 | −1117.4307 | no | yes | |
| Terrein (10) | 19 | −5.9071436 | −1112.7003 | Arg483 ⟷ O1/2.084 Arg483 ⟷ O2/2.215 | yes |
| 12 | −5.8997755 | −1106.2816 | Ser508 ⟷ H4/2.276 | yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yurchenko, E.A.; Khmel, O.O.; Nesterenko, L.E.; Aminin, D.L. The Kelch/Nrf2 Antioxidant System as a Target for Some Marine Fungal Metabolites. Oxygen 2023, 3, 374-385. https://doi.org/10.3390/oxygen3040024
Yurchenko EA, Khmel OO, Nesterenko LE, Aminin DL. The Kelch/Nrf2 Antioxidant System as a Target for Some Marine Fungal Metabolites. Oxygen. 2023; 3(4):374-385. https://doi.org/10.3390/oxygen3040024
Chicago/Turabian StyleYurchenko, Ekaterina A., Olga O. Khmel, Liliana E. Nesterenko, and Dmitry L. Aminin. 2023. "The Kelch/Nrf2 Antioxidant System as a Target for Some Marine Fungal Metabolites" Oxygen 3, no. 4: 374-385. https://doi.org/10.3390/oxygen3040024
APA StyleYurchenko, E. A., Khmel, O. O., Nesterenko, L. E., & Aminin, D. L. (2023). The Kelch/Nrf2 Antioxidant System as a Target for Some Marine Fungal Metabolites. Oxygen, 3(4), 374-385. https://doi.org/10.3390/oxygen3040024