
A Model of Hepatitis B Viral Dynamics with Delays


Abstract
:1. Introduction
2. Materials and Methods
2.1. Basic Model
2.2. Model 2
- Similarly, the fourth equation implies
2.3. Basic Model with Delay
2.4. Model 2 with Delay
3. Results
3.1. Threshold Indices
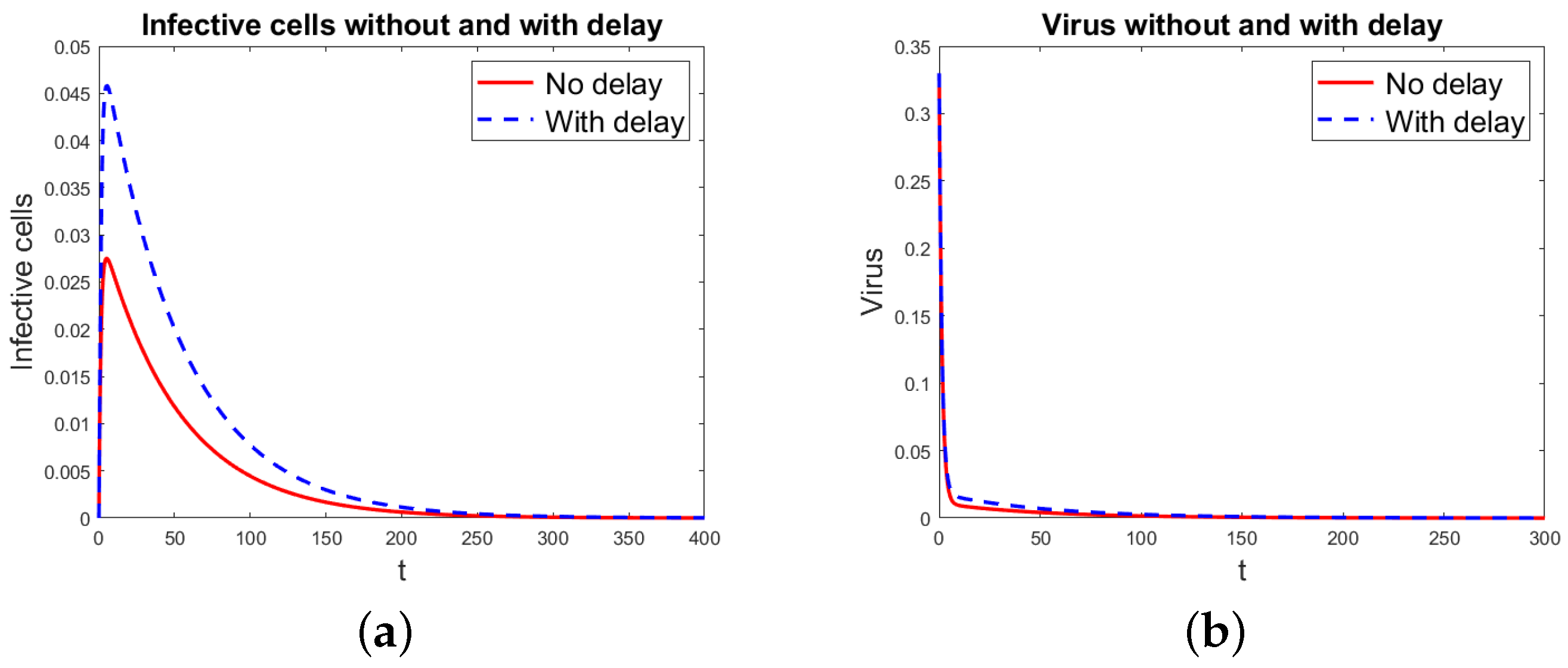
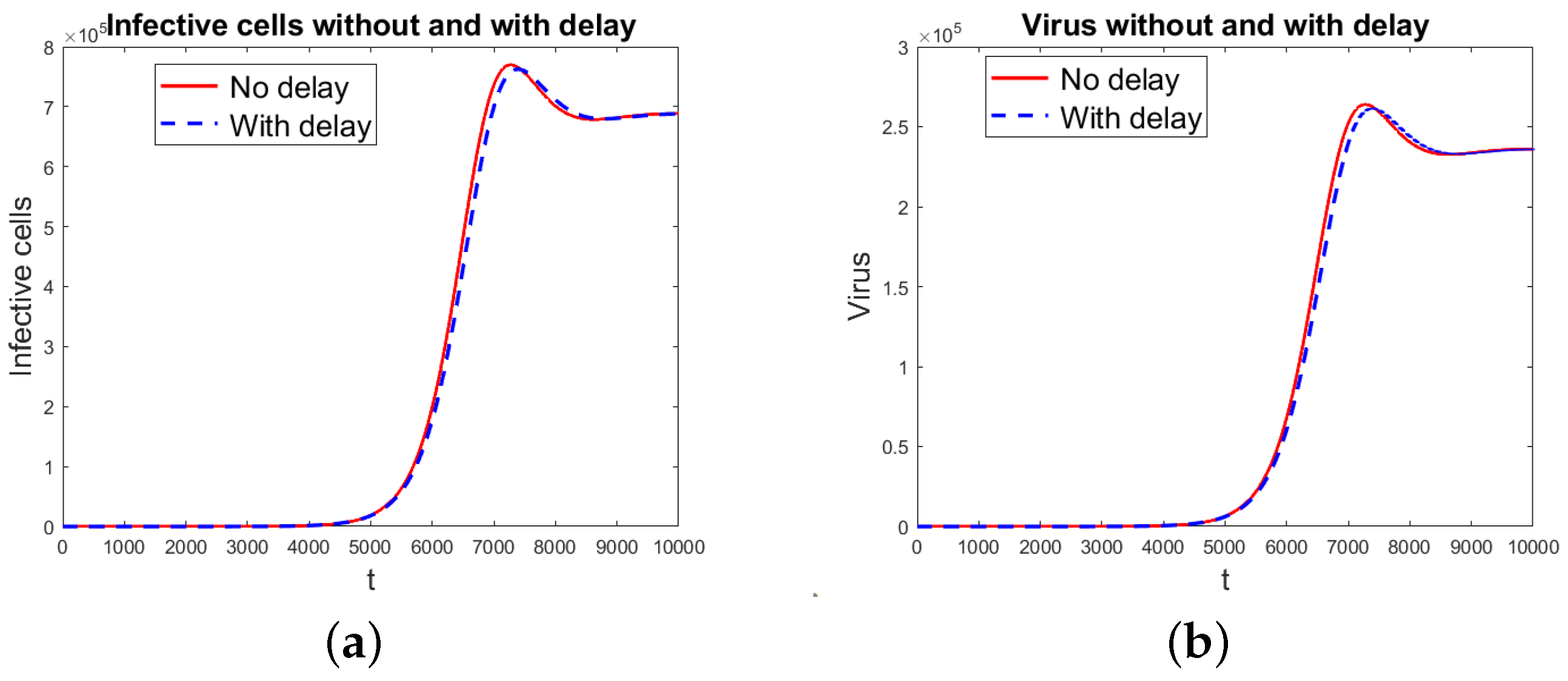
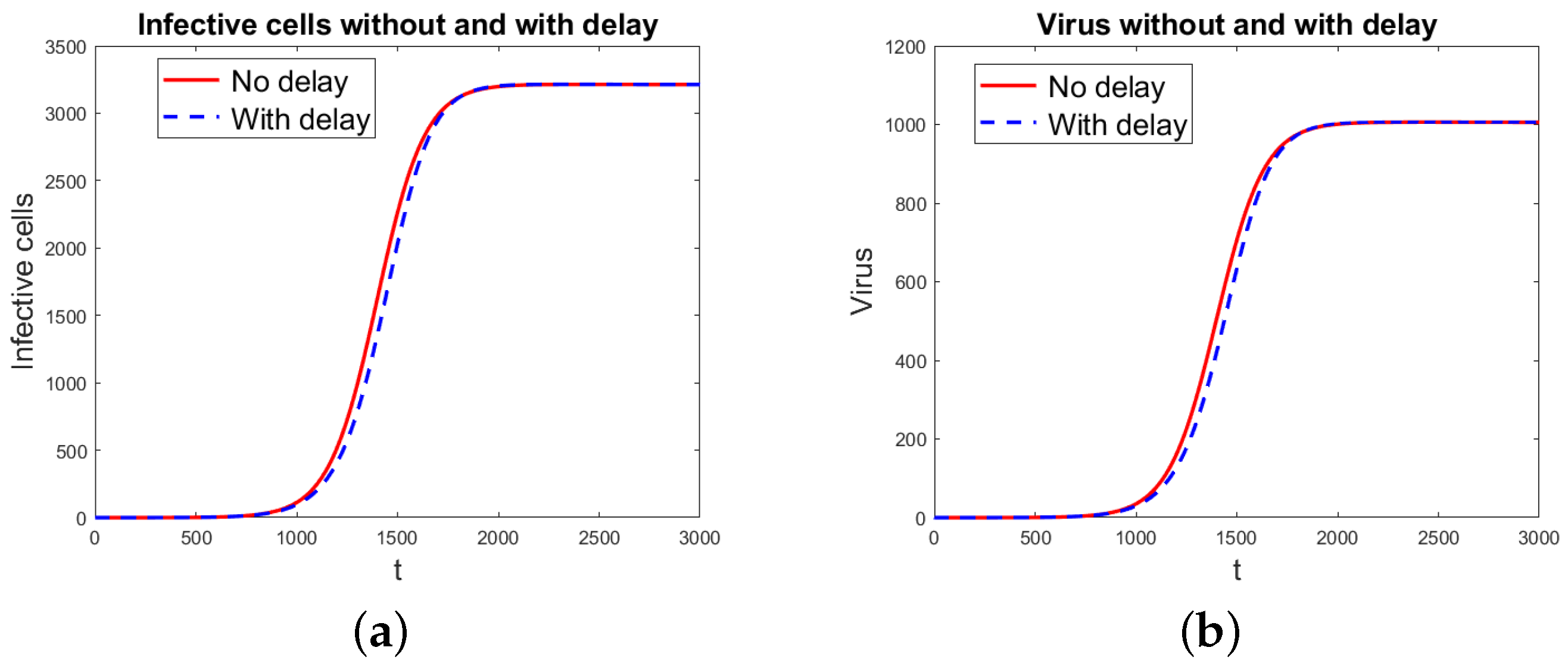
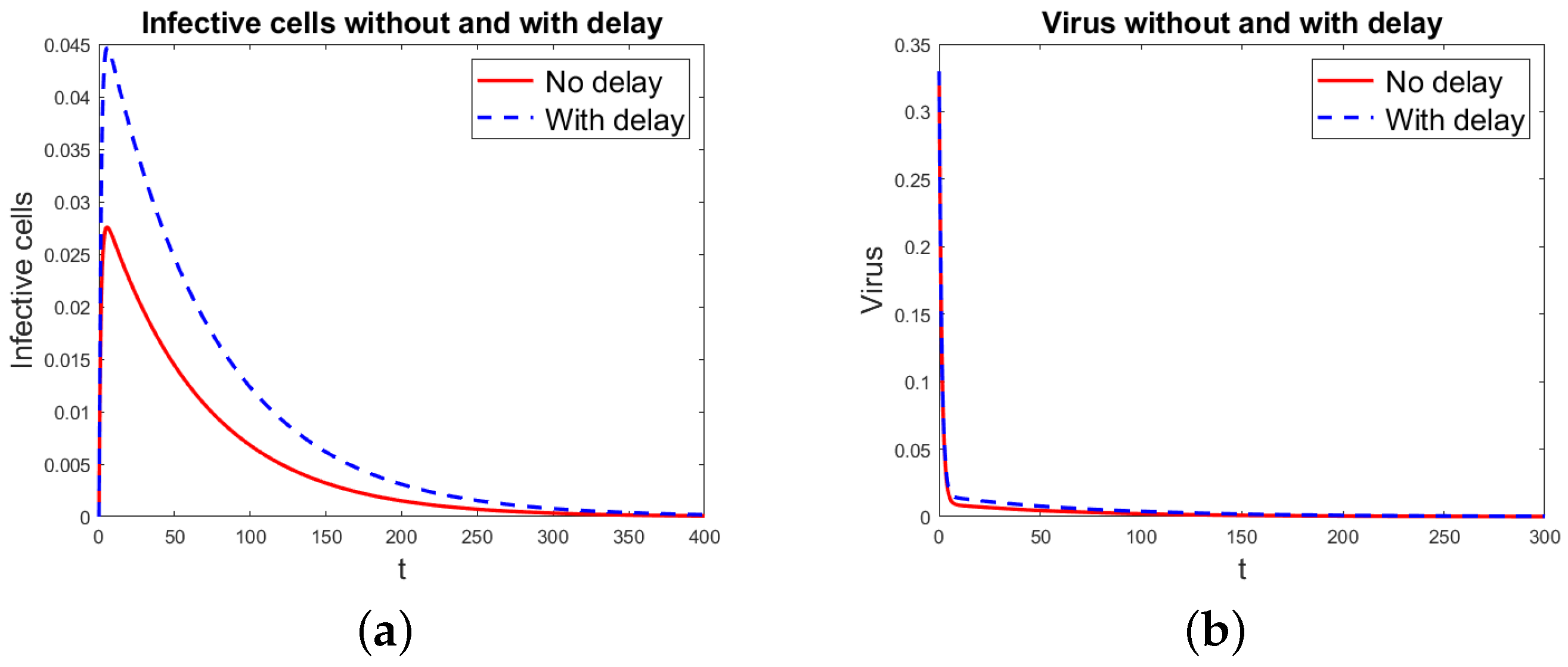
Numerical Simulations
4. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ODE | Ordinary differential equation |
| DDE | Delay differential equation |
| DFE | Disease-free equilibrium |
| CE | Chronic equilibrium |
References
- Center for Disease Control and Prevention Web Page for Hepatitis B. Available online: https://www.cdc.gov/hepatitis/hbv/index.htm (accessed on 17 October 2023).
- World Heath Organization Web Page for Hepatitis B. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-b (accessed on 17 October 2023).
- Din, A.; Li, Y. Mathematical analysis of a new nonlinear stochastic hepatitis B epidemic model with vaccination effect and a case study. Eur. Phys. J. Plus 2022, 137, 1–24. [Google Scholar] [CrossRef]
- Wodajo, F.A.; Gebru, D.M.; Alemneh, H.T. Mathematical model analysis of effective intervention strategies on transmission dynamics of hepatitis B virus. Sci. Rep. 2023, 13, 8737. [Google Scholar] [CrossRef]
- Zada, I.; Naeem Jan, M.; Ali, N.; Alrowail, D.; Sooppy Nisar, K.; Zaman, G. Mathematical analysis of hepatitis B epidemic model with optimal control. Adv. Differ. Equ. 2021, 2021, 1–29. [Google Scholar] [CrossRef]
- Oludoun, O.; Adebimpe, O.; Ndako, J.; Adeniyi, M.; Abiodun, O.; Gbadamosi, B. The impact of testing and treatment on the dynamics of Hepatitis B virus. F1000Research 2021, 10, 936. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Infectious Diseases of Humans: Dynamics and Control; Oxford University Press: Oxford, UK, 1991. [Google Scholar]
- Nowak, M.A.; Bangham, C.R. Population dynamics of immune responses to persistent viruses. Science 1996, 272, 74–79. [Google Scholar] [CrossRef]
- Bonhoeffer, S.; May, R.M.; Shaw, G.M.; Nowak, M.A. Virus dynamics and drug therapy. Proc. Natl. Acad. Sci. USA 1997, 94, 6971–6976. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.; May, R.M. Virus Dynamics: Mathematical Principles of Immunology and Virology: Mathematical Principles of Immunology and Virology; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Regoes, R.R.; Wodarz, D.; Nowak, M.A. Virus dynamics: The effect of target cell limitation and immune responses on virus evolution. J. Theor. Biol. 1998, 191, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Cui, J.A.; Hui, J. The importance of immune responses in a model of hepatitis B virus. Nonlinear Dyn. 2012, 67, 723–734. [Google Scholar] [CrossRef]
- Ciupe, S.M.; Ribeiro, R.M.; Perelson, A.S. Antibody responses during hepatitis B viral infection. PLoS Comput. Biol. 2014, 10, e1003730. [Google Scholar] [CrossRef]
- Wodarz, D.; Nowak, M.A. Mathematical models of HIV pathogenesis and treatment. BioEssays 2002, 24, 1178–1187. [Google Scholar] [CrossRef]
- Nowak, M.A.; Bonhoeffer, S.; Hill, A.M.; Boehme, R.; Thomas, H.C.; McDade, H. Viral dynamics in hepatitis B virus infection. Proc. Natl. Acad. Sci. USA 1996, 93, 4398–4402. [Google Scholar] [CrossRef]
- Kuang, Y. Delay Differential Equations: With Applications in Population Dynamics; Academic Press: Cambridge, MA, USA, 1993. [Google Scholar]
- Erneux, T. Applied Delay Differential Equations; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009; Volume 3. [Google Scholar]
- Ruan, S. Delay differential equations in single species dynamics. In Delay Differential Equations and Applications; Springer: Berlin/Heidelberg, Germany, 2006; pp. 477–517. [Google Scholar]
- Ciupe, S.M.; Ribeiro, R.M.; Nelson, P.W.; Dusheiko, G.; Perelson, A.S. The role of cells refractory to productive infection in acute hepatitis B viral dynamics. Proc. Natl. Acad. Sci. USA 2007, 104, 5050–5055. [Google Scholar] [CrossRef]
- Kim, H.Y.; Kwon, H.D.; Jang, T.S.; Lim, J.; Lee, H.S. Mathematical modeling of triphasic viral dynamics in patients with HBeAg-positive chronic hepatitis B showing response to 24-week clevudine therapy. PLoS ONE 2012, 7, e50377. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, J.; Li, Y.; Jiang, Z.; Han, X. Dynamics analysis of a delayed virus model with two different transmission methods and treatments. Adv. Differ. Equ. 2020, 2020, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Dagasso, G.; Urban, J.; Kwiatkowska, M. Incorporating time delays in the mathematical modelling of the human immune response in viral infections. Procedia Comput. Sci. 2021, 185, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Yosyingyong, P.; Viriyapong, R. Global dynamics of multiple delays within-host model for a hepatitis B virus infection of hepatocytes with immune response and drug therapy. Math. Biosci. Eng. 2023, 20, 7349–7386. [Google Scholar] [CrossRef] [PubMed]
- Rihan, F.A.; Alsakaji, H.J. Analysis of a stochastic HBV infection model with delayed immune response. Math. Biosci. Eng. 2021, 18, 5194–5220. [Google Scholar] [CrossRef] [PubMed]
- Li, D.M.; Chai, B.; Wang, Q. A model of hepatitis B virus with random interference infection rate. Math. Biosci. Eng. 2021, 18, 8257–8297. [Google Scholar] [CrossRef]
- Goyal, A.; Liao, L.E.; Perelson, A.S. Within-host mathematical models of hepatitis B virus infection: Past, present, and future. Curr. Opin. Syst. Biol. 2019, 18, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Pourbashash, H.; Pilyugin, S.S.; De Leenheer, P.; McCluskey, C. Global analysis of within-host virus models with cell-to-cell viral transmission. Discret. Contin. Dyn. Syst. Ser. B 2014, 19, 3341–3357. [Google Scholar] [CrossRef]
- Elbaz, I.M.; El-Metwally, H.; Sohaly, M. Viral kinetics, stability and sensitivity analysis of the within-host COVID-19 model. Sci. Rep. 2023, 13, 11675. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Liu, A.; Foryś, U. Global stability analysis of some nonlinear delay differential equations in population dynamics. J. Nonlinear Sci. 2016, 26, 27–41. [Google Scholar] [CrossRef]
- Lv, J.; Ma, W. Global asymptotic stability of a delay differential equation model for SARS-CoV-2 virus infection mediated by ACE2 receptor protein. Appl. Math. Lett. 2023, 142, 108631. [Google Scholar] [CrossRef] [PubMed]
- Orosz, G. Hopf bifurcation calculations in delayed systems. Period. Polytech. Mech. Eng. 2004, 48, 189–200. [Google Scholar]
- Jiang, X.; Chen, X.; Chi, M.; Chen, J. On Hopf bifurcation and control for a delay systems. Appl. Math. Comput. 2020, 370, 124906. [Google Scholar] [CrossRef]
- Engelborghs, K.; Luzyanina, T.; Roose, D. Numerical bifurcation analysis of delay differential equations using DDE-BIFTOOL. ACM Trans. Math. Softw. 2002, 28, 1–21. [Google Scholar] [CrossRef]
- Cui, Q.; Xu, C.; Ou, W.; Pang, Y.; Liu, Z.; Li, P.; Yao, L. Bifurcation Behavior and Hybrid Controller Design of a 2D Lotka–Volterra Commensal Symbiosis System Accompanying Delay. Mathematics 2023, 11, 4808. [Google Scholar] [CrossRef]
- Ou, W.; Xu, C.; Cui, Q.; Pang, Y.; Liu, Z.; Shen, J.; Baber, M.Z.; Farman, M.; Ahmad, S. Hopf bifurcation exploration and control technique in a predator-prey system incorporating delay. AIMS Math. 2024, 9, 1622–1651. [Google Scholar] [CrossRef]
- Tang, L.S.; Covert, E.; Wilson, E.; Kottilil, S. Chronic hepatitis B infection: A review. JAMA 2018, 319, 1802–1813. [Google Scholar] [CrossRef]
- Volinsky, I. Mathematical Model of Hepatitis B Virus Treatment with Support of Immune System. Mathematics 2022, 10, 2821. [Google Scholar] [CrossRef]
- Hu, J.; Liu, K. Complete and incomplete hepatitis B virus particles: Formation, function, and application. Viruses 2017, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Tu, T.; Zhang, H.; Urban, S. Hepatitis B Virus DNA Integration: In Vitro Models for Investigating Viral Pathogenesis and Persistence. Viruses 2021, 13, 180. [Google Scholar] [CrossRef] [PubMed]
- Van den Driessche, P.; Watmough, J. Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 2002, 180, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Van den Driessche, P. Reproduction numbers of infectious disease models. Infect. Dis. Model. 2017, 2, 288–303. [Google Scholar] [CrossRef] [PubMed]
- Diekmann, O.; Heesterbeek, J.; Roberts, M.G. The construction of next-generation matrices for compartmental epidemic models. J. R. Soc. Interface 2010, 7, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Hefferman, J.; Smith, R.; Wahl, L. Perspectives on the basic reproduction ratio. J. R. Soc. Interface 2005, 2, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.D. Mathematical Biology I. An Introduction; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Allen, L. An Introduction to Mathematical Biology; Pearson-Prentice Hall: Old Bridge, NJ, USA, 2007. [Google Scholar]
- Wei, H.M.; Li, X.Z.; Martcheva, M. An epidemic model of a vector-borne disease with direct transmission and time delay. J. Math. Anal. Appl. 2008, 342, 895–908. [Google Scholar] [CrossRef]
- Al Basir, F.; Takeuchi, Y.; Ray, S.A. Dynamics of a delayed plant disease model with Beddington-DeAngelis disease transmission. Math. Biosci. Eng. 2021, 18, 583–599. [Google Scholar] [CrossRef]
- Wodarz, D. Mathematical models of immune effector responses to viral infections: Virus control versus the development of pathology. J. Comput. Appl. Math. 2005, 184, 301–319. [Google Scholar] [CrossRef]
- Zhang, S.; Li, F.; Xu, X. Dynamics and control strategy for a delayed viral infection model. J. Biol. Dyn. 2022, 16, 44–63. [Google Scholar] [CrossRef]
- Bauer, T.; Sprinzl, M.; Protzer, U. Immune control of hepatitis B virus. Dig. Dis. 2011, 29, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.L. An Introduction to Delay Differential Equations with Applications to the Life Sciences; Springer: New York, NY, USA, 2011; Volume 57. [Google Scholar]
- Lok, A.S.; McMahon, B.J. Chronic hepatitis B. Hepatology 2007, 45, 507–539. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | Description |
|---|---|---|
| 5 × cells/mL 1/d | recruitment rate of susceptible cells | |
| 0.003/d | death rate of susceptible cells | |
| 4 × mL/(cells d) | infection rate of susceptible cells by virus | |
| 0.043/d | death rate of infected cells | |
| B | 5.58 | number of virions produce by 1 infected cell |
| 0.7 | death rate of virus | |
| 1 d | delay in time of infection |
| Parameter | Value | Description |
|---|---|---|
| 4 × mL/(cells d) | infection rate of susceptible cells by virus | |
| 0.6 × mL/(cells d) | elimination rate of infected cells by effector cells | |
| 0.6 × mL/(cells d) | removal rate of effector cells after elimination of infected cells | |
| 4 × mL/(cells d) | elimination rate of virus by effector cells | |
| 4 × mL/(cells d) | removal rate of effector cells after elimination of virus | |
| s | 24 cells/mL 1/d | recruitment rate of effector cells |
| 2.2 × /d | recruitment rate of effector cells due to infected cells | |
| 0.5/d | death rate of effector cells | |
| 1 d | delay in time of infection | |
| 24 d | delay in recruiting of effector cells due to infected cells |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen-Charpentier, B. A Model of Hepatitis B Viral Dynamics with Delays. AppliedMath 2024, 4, 182-196. https://doi.org/10.3390/appliedmath4010009
Chen-Charpentier B. A Model of Hepatitis B Viral Dynamics with Delays. AppliedMath. 2024; 4(1):182-196. https://doi.org/10.3390/appliedmath4010009
Chicago/Turabian StyleChen-Charpentier, Benito. 2024. "A Model of Hepatitis B Viral Dynamics with Delays" AppliedMath 4, no. 1: 182-196. https://doi.org/10.3390/appliedmath4010009
APA StyleChen-Charpentier, B. (2024). A Model of Hepatitis B Viral Dynamics with Delays. AppliedMath, 4(1), 182-196. https://doi.org/10.3390/appliedmath4010009