


Comparative Study of Cis- and Trans-Priming Effect of PEG and BABA in Cowpea Seedlings on Exposure to PEG-Induced Osmotic Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Priming
2.2. Experimental Design
2.3. Estimation of Chlorophyll Content
2.4. Estimation of Malondialdehyde Content and Reactive Oxygen Species
2.5. Determination of Leaf Osmolality
2.6. Estimation of Non-Enzymatic Antioxidants
2.7. Determination of Enzymatic Antioxidants
2.8. Measurements of Chlorophyll a Fluorescence Parameters
2.9. Statistical Analysis
3. Results
3.1. Selection of Stress Imparting Concentration of PEG and Standardisation of Effective Concentrations of Priming Agents
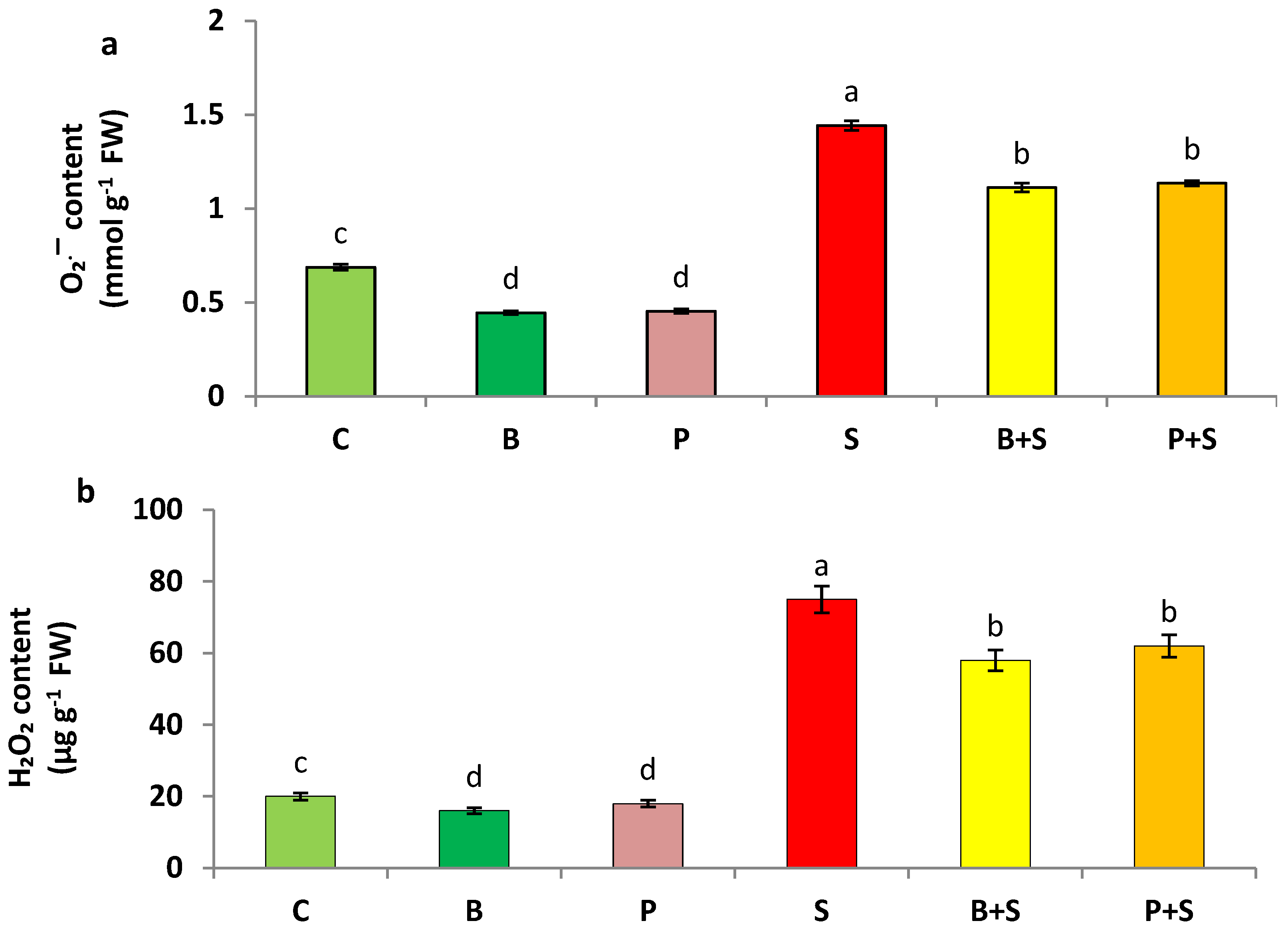
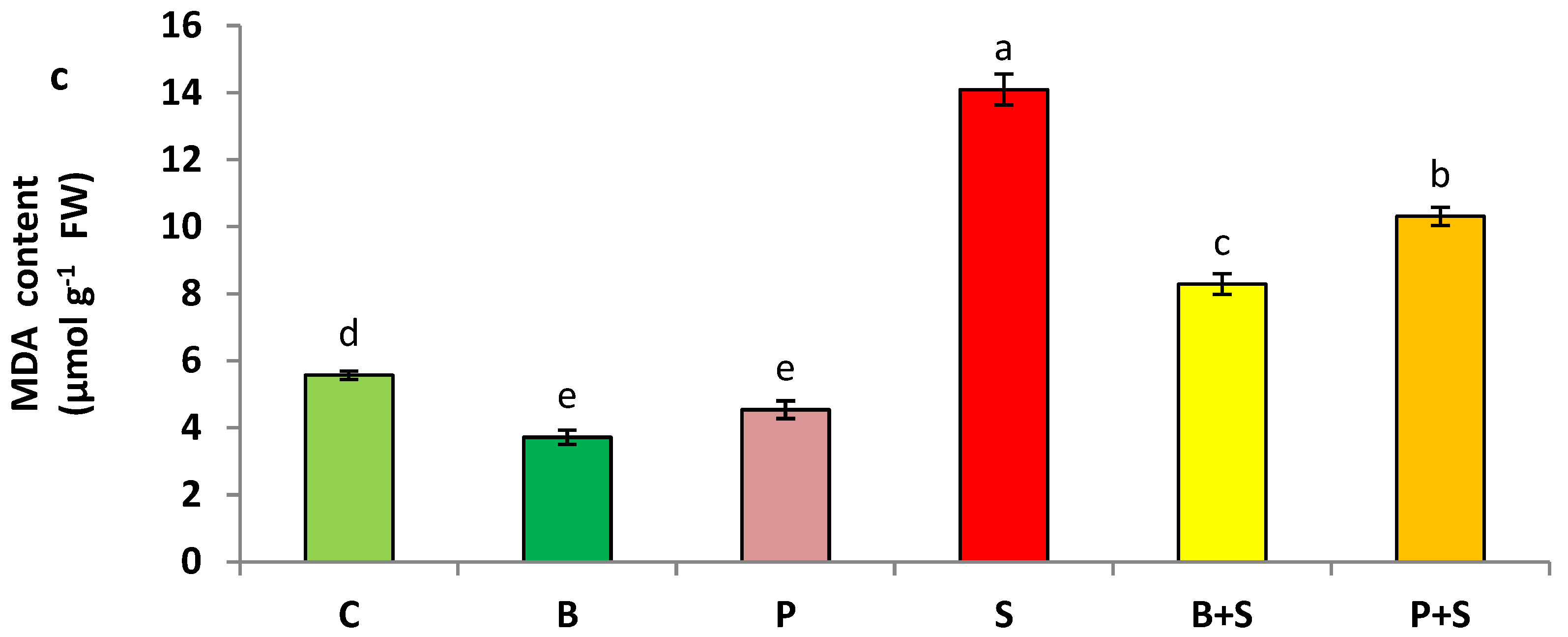
3.2. Oxidative Stress Markers
3.3. Osmolality
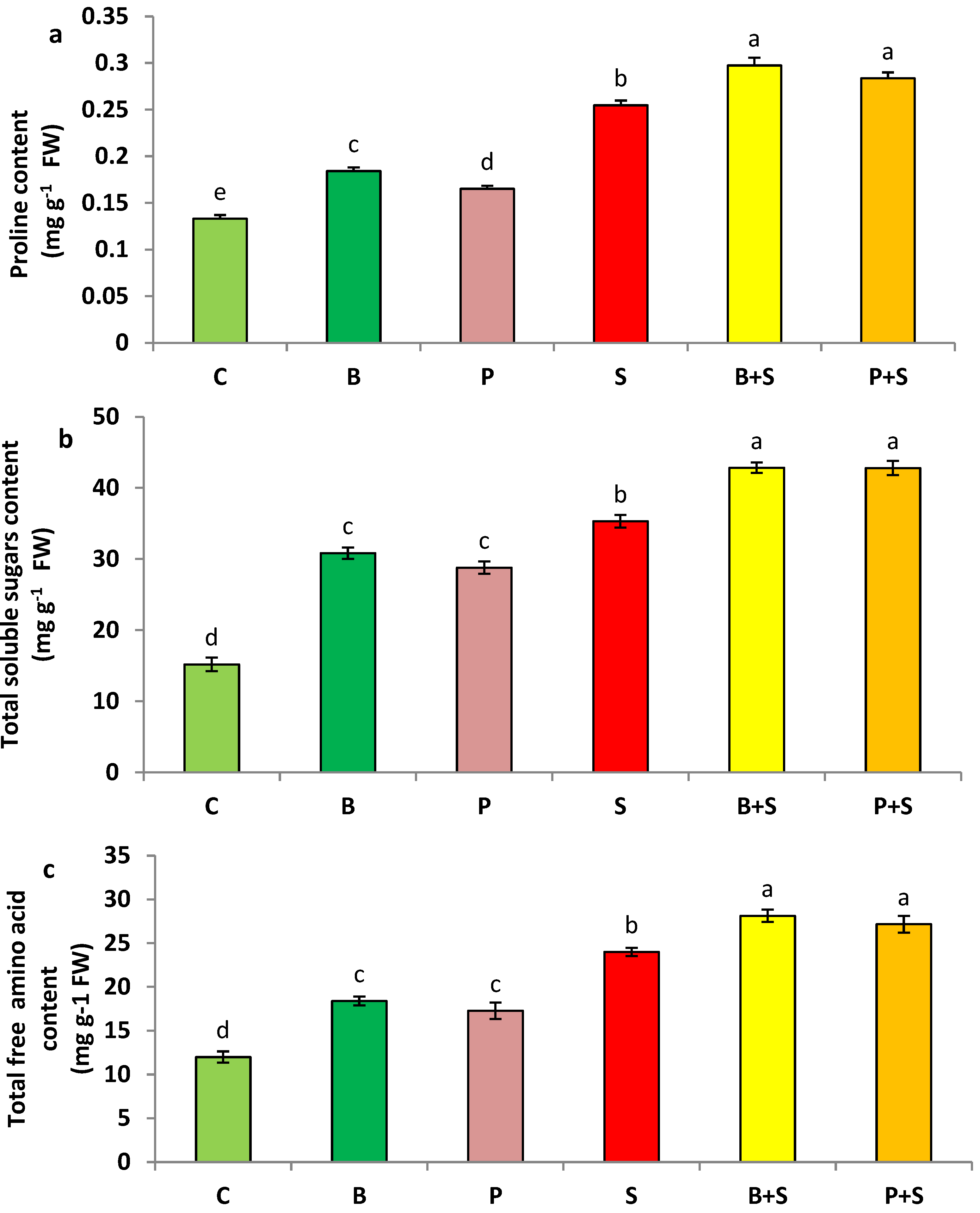
3.4. Non-Enzymatic Antioxidants
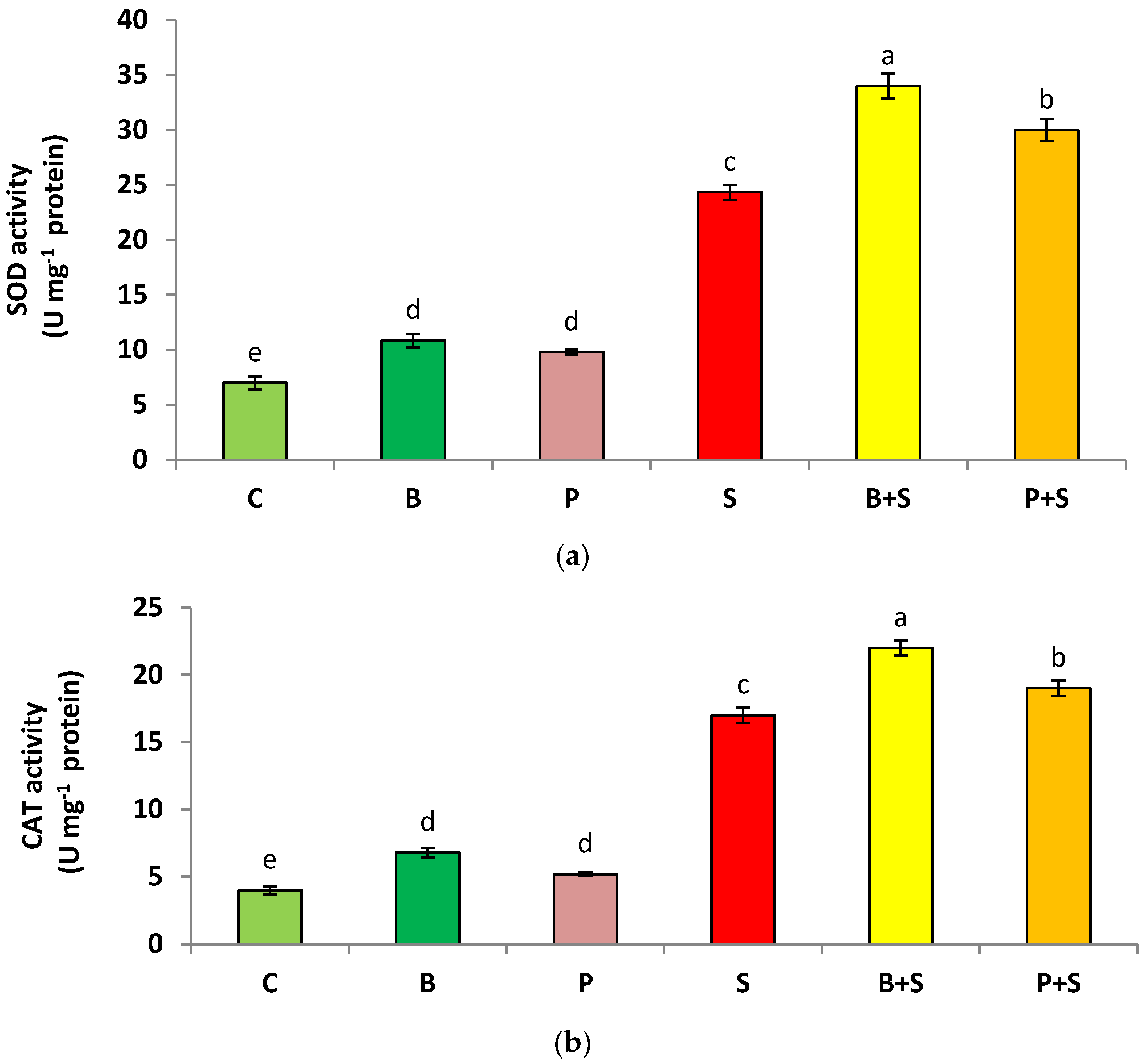
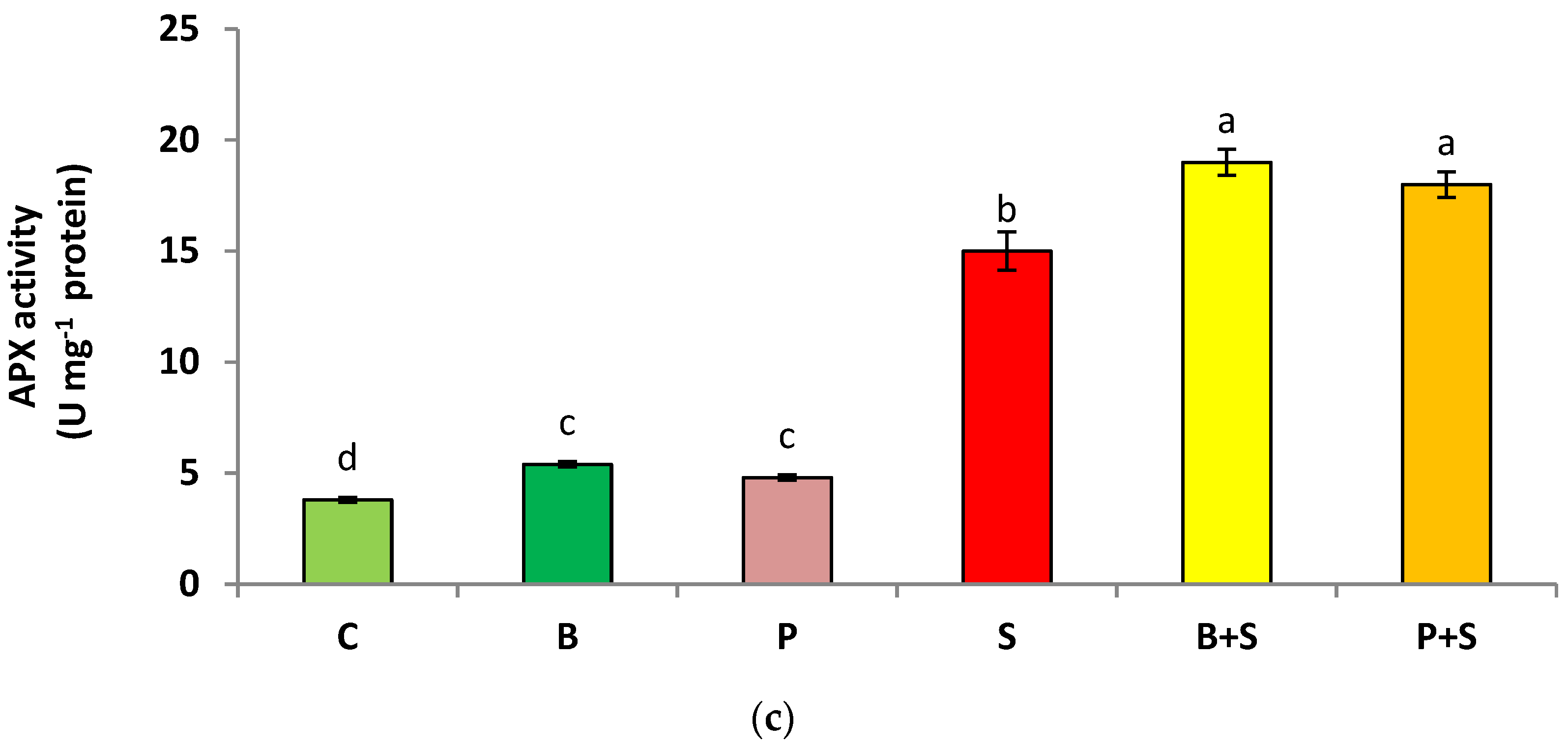
3.5. Enzymatic Antioxidants
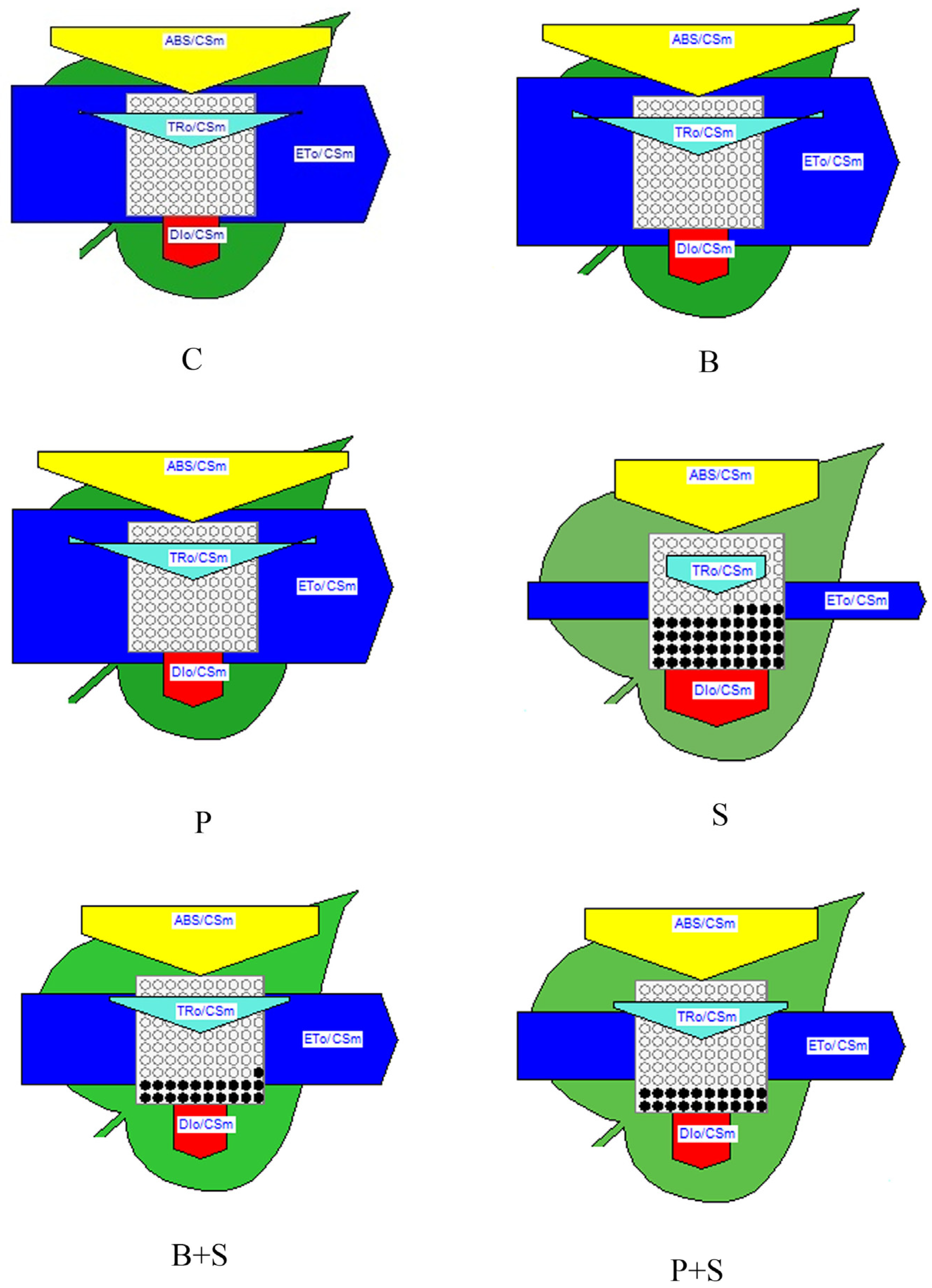
3.6. Photobiology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Basal, O.; Szabó, A.; Veres, S. Physiology of soybean as affected by PEG-induced drought stress. Curr. Plant Biol. 2020, 22, 100135. [Google Scholar] [CrossRef]
- Ru, C.; Wang, K.; Hu, X.; Chen, D.; Wang, W.; Yang, H. Nitrogen modulates the effects of heat, drought, and combined stresses on photosynthesis, antioxidant capacity, cell osmoregulation, and grain yield in winter wheat. J. Plant Growth Regul. 2022, 1–23. [Google Scholar] [CrossRef]
- Osmolovskaya, N.; Shumilina, J.; Kim, A.; Didio, A.; Grishina, T.; Bilova, T.; Keltsieva, O.A.; Zhukov, V.; Tikhonovich, I.; Tarakhovskaya, E.; et al. Methodology of drought stress research: Experimental setup and physiological characterization. Int. J. Mol. Sci. 2018, 19, 4089. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Luo, X.; Wei, M.; Khan, A.; Munsif, F.; Huang, T.; Pan, X.; Shan, Z. Antioxidant enzymatic activity and its related genes expression in cassava leaves at different growth stages play key roles in sustaining yield and drought tolerance under moisture stress. J. Plant Growth Regul. 2020, 39, 594–607. [Google Scholar] [CrossRef]
- Goufo, P.; Moutinho-Pereira, J.M.; Jorge, T.F.; Correia, C.M.; Oliveira, M.R.; Rosa, E.A.S.; António, C.; Trindade, H. Cowpea (Vigna unguiculata L. Walp.) metabolomics: Osmoprotection as a physiological strategy for drought stress resistance and improved yield. Front. Plant Sci. 2017, 8, 586. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Gogoi, N.; Barthakur, S.; Baroowa, B.; Bharadwaj, N.; Alghamdi, S.S.; Siddique, K.H.M. Drought stress in grain legumes during reproduction and grain filling. J. Agron. Crop Sci. 2017, 203, 81–102. [Google Scholar] [CrossRef]
- Paparella, S.; Araújo, S.S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed priming: State of the art and new perspectives. Plant Cell Rep. 2015, 34, 1281–1293. [Google Scholar] [CrossRef]
- Tania, S.S.; Rhaman, M.S.; Rauf, F.; Rahaman, M.M.; Kabir, M.H.; Hoque, M.A.; Murata, Y. Alleviation of salt-inhibited germination and seedling growth of kidney bean by seed priming and exogenous application of salicylic acid (SA) and hydrogen peroxide (H2O2). Seeds 2022, 1, 87–98. [Google Scholar] [CrossRef]
- Khan, M.N.; Khan, Z.; Luo, T.; Liu, J.; Rizwan, M.; Zhang, J.; Xu, Z.; Wu, H.; Hu, L. Seed Priming with gibberellic acid and melatonin in rapeseed: Consequences for improving yield and seed quality under drought and non-stress conditions. Ind. Crops Prod. 2020, 156, 112850. [Google Scholar] [CrossRef]
- Farooq, M.; Romdhane, L.; Al Sulti, M.K.R.A.; Rehman, A.; Al-Busaidi, W.M.; Lee, D. Morphological, physiological and biochemical aspects of osmopriming-induced drought tolerance in lentil. J. Agron. Crop Sci. 2020, 206, 176–186. [Google Scholar] [CrossRef]
- Abid, M.; Hakeem, A.; Shao, Y.; Liu, Y.; Zahoor, R.; Fan, Y.; Suyu, J.; Ata-Ul-Karim, S.T.; Tian, Z.; Jiang, D.; et al. Seed osmopriming invokes stress memory against post-germinative drought stress in wheat (Triticum aestivum L.). Environ. Exp. Bot. 2018, 145, 12–20. [Google Scholar] [CrossRef]
- Aswathi, K.P.R.; Kalaji, H.M.; Puthur, J.T. seed priming of plants aiding in drought stress tolerance and faster recovery: A review. Plant Growth Regul. 2022, 97, 235–253. [Google Scholar] [CrossRef]
- Jisha, K.C.; Puthur, J.T. Seed priming with BABA (β-amino butyric acid): A cost-effective method of abiotic stress tolerance in Vigna radiata (L.) Wilczek. Protoplasma 2016, 253, 277–289. [Google Scholar] [CrossRef]
- Ferreira Ribas, A.; Volpi e Silva, N.; dos Santos, T.B.; Lima Abrantes, F.; Castilho Custódio, C.; Barbosa Machado-Neto, N.; Esteves Vieira, L.G. Regulation of α-expansins genes in Arabidopsis Thaliana seeds during post-osmopriming germination. Physiol. Mol. Biol. Plants 2019, 25, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Macarisin, D.; Wisniewski, M.E.; Bassett, C.; Thannhauser, T.W. Proteomic analysis of β-aminobutyric acid priming and abscisic acid–induction of drought resistance in crab apple (Malus pumila): Effect on general metabolism, the phenylpropanoid pathway and cell wall enzymes. Plant Cell Environ. 2009, 32, 1612–1631. [Google Scholar] [CrossRef]
- Abid, G.; Ouertani, R.N.; Jebara, S.H.; Boubakri, H.; Muhovski, Y.; Ghouili, E.; Abdelkarim, S.; Chaieb, O.; Hidri, Y.; Kadri, S.; et al. Alleviation of drought stress in faba bean (Vicia faba L.) by exogenous application of β-aminobutyric acid (BABA). Physiol. Mol. Biol. Plants 2020, 26, 1173–1186. [Google Scholar] [CrossRef]
- Shaw, A.K.; Bhardwaj, P.K.; Ghosh, S.; Roy, S.; Saha, S.; Sherpa, A.R.; Saha, S.K.; Hossain, Z. β-aminobutyric acid mediated drought stress alleviation in maize (Zea mays L.). Environ. Sci. Pollut. Res. Int. 2016, 23, 2437–2453. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Doke, N. Involvement of superoxide anion generation in the hypersensitive response of potato tuber tissues to infection with an incompatible race of Phytophthora infestans and to the hyphal wall components. Physiol. Plant Pathol. 1983, 23, 345–357. [Google Scholar] [CrossRef]
- Junglee, S.; Urban, L.; Sallanon, H.; Lopez-Lauri, F. Optimized assay for hydrogen peroxide determination in plant tissue using potassium iodide. Am. J. Anal. Chem. 2014, 5, 730–736. [Google Scholar] [CrossRef]
- Hura, T.; Grzesiak, S.; Hura, K.; Thiemt, E.; Tokarz, K.; Wedzony, M. Physiological and biochemical tools useful in drought-tolerance detection in genotypes of winter triticale: Accumulation of ferulic acid correlates with drought tolerance. Ann. Bot. 2007, 100, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.K. Rapid determination of free proline for water studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Moore, S.; Stein, W.H. Photometric ninhydrin method for use in the chromatography of amino acids. J. Biol. Chem. 1948, 176, 367–388. [Google Scholar] [CrossRef]
- Chen, J.X.; Wang, X.F. Guide to Plant Physiological Experiments; South China University of Technology Press: Guangzhou, China, 2002; pp. 123–127. [Google Scholar]
- Folin, O.; Denis, W. A Colorimetric method for the determination of phenols (and phenol derivatives) in urine. J. Biol. Chem. 1915, 22, 305–308. [Google Scholar] [CrossRef]
- Yin, D.; Chen, S.; Chen, F.; Guan, Z.; Fang, W. Morphological and physiological responses of two chrysanthemum cultivars differing in their tolerance to waterlogging. Environ. Exp. Bot. 2009, 67, 87–93. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of proteins utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence; Papageorgiou, G.C., Govindjee, G., Eds.; Springer Science+Business Media: Berlin/Heidelberg, Germany, 2004; pp. 321–362. [Google Scholar] [CrossRef]
- Liu, H.; Able, A.J.; Able, J.A. Priming crops for the future: Rewiring stress memory. Trends Plant Sci. 2022, 27, 699–716. [Google Scholar] [CrossRef] [PubMed]
- Jisha, K.C.; Puthur, J.T. Seed halopriming outdo hydropriming in enhancing seedling vigor and osmotic stress tolerance potential of rice varieties. J. Crop Sci. Biotechnol. 2014, 17, 209–219. [Google Scholar] [CrossRef]
- Thomas, D.T.; Puthur, J.T. Amplification of abiotic stress tolerance potential in rice seedlings with a low dose of UV-B seed priming. Funct. Plant Biol. 2019, 46, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; Challabathula, D.; Puthur, J.T. UV-B priming of Oryza sativa seeds augments the innate tolerance potential in a tolerant variety more effectively toward NaCl and PEG stressors. J. Plant Growth Regul. 2021, 40, 1166–1180. [Google Scholar] [CrossRef]
- Dhanya Thomas, T.T.; Dinakar, C.; Puthur, J.T. Effect of UV-B priming on the abiotic stress tolerance of stress-sensitive rice seedlings: Priming imprints and cross-tolerance. Plant Physiol. Biochem. 2020, 147, 21–30. [Google Scholar] [CrossRef]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical priming of plants against multiple abiotic stresses: Mission possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, X.; Dong, Y.; Zhang, F.; He, Q.; Chen, J.; Zhu, S.; Zhao, T. Seed priming with melatonin improves salt tolerance in cotton through regulating photosynthesis, scavenging reactive oxygen species and coordinating with phytohormone signal pathways. Ind. Crops Prod. 2021, 169, 113671. [Google Scholar] [CrossRef]
- Mostek, A.; Börner, A.; Weidner, S. comparative proteomic analysis of β-aminobutyric acid-mediated alleviation of salt stress in barley. Plant Physiol. Biochem. 2016, 99, 150–161. [Google Scholar] [CrossRef]
- Zhang, F.; Yu, J.; Johnston, C.R.; Wang, Y.; Zhu, K.; Lu, F.; Zhang, Z.; Zou, J. Seed priming with polyethylene glycol induces physiological changes in sorghum (Sorghum bicolor L. Moench) seedlings under suboptimal soil moisture environments. PLoS ONE 2015, 10, e0140620. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Zhi, W.; Feng, Y.; Zhou, G.; Zhu, G. Seed priming improved salt-stressed sorghum growth by enhancing antioxidative defense. PLoS ONE 2022, 17, e0263036. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Shi, S.; Wang, B.; Zhao, J. Physiological and biochemical changes in different drought-tolerant alfalfa (Medicago sativa L.) varieties under PEG-induced drought stress. Acta Physiol. Plant. 2018, 40, 1–15. [Google Scholar] [CrossRef]
- Hosseini Boldaji, S.A.; Khavari-Nejad, R.A.; Hassan Sajedi, R.; Fahimi, H.; Saadatmand, S. Water availability effects on antioxidant enzyme activities, lipid peroxidation, and reducing sugar contents of alfalfa (Medicago sativa L.). Acta Physiol. Plant. 2012, 34, 1177–1186. [Google Scholar] [CrossRef]
- Penella, C.; Nebauer, S.G.; Bautista, A.S.; López-Galarza, S.; Calatayud, Á. Rootstock alleviates PEG-induced water stress in grafted pepper seedlings: Physiological responses. J. Plant Physiol. 2014, 171, 842–851. [Google Scholar] [CrossRef]
- Hannachi, S.; Werbrouck, S.; Bahrini, I.; Abdelgadir, A.; Siddiqui, H.A.; Van Labeke, M.C. Obtaining salt stress-tolerant eggplant somaclonal variants from in vitro selection. Plants 2021, 10, 2539. [Google Scholar] [CrossRef]
- Koh, Y.S.; Wong, S.K.; Ismail, N.H.; Zengin, G.; Duangjai, A.; Saokaew, S.; Phisalprapa, P.; Tan, K.W.; Goh, B.H.; Tang, S.Y. Mitigation of environmental stress-impacts in plants: Role of sole and combinatory exogenous application of glutathione. Front. Plant Sci. 2021, 12, 791205. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- ElSayed, A.I.; El-Hamahmy, M.A.M.; Rafudeen, M.S.; Mohamed, A.H.; Omar, A.A. The impact of drought stress on antioxidant responses and accumulation of flavonolignans in milk thistle (Silybum marianum (L.) Gaertn). Plants 2019, 8, 611. [Google Scholar] [CrossRef]
- Batool, M.; El-Badri, A.M.; Hassan, M.U.; Haiyun, Y.; Chunyun, W.; Zhenkun, Y.; Jie, K.; Wang, B.; Zhou, G. Drought Stress in Brassica napus: Effects, tolerance mechanisms, and management strategies. J. Plant Growth Regul. 2022, 42, 1–25. [Google Scholar] [CrossRef]
- Zhang, K.; Khan, Z.; Wu, H.; Khan, M.N.; Hu, L. Gibberellic acid priming improved rapeseed drought tolerance by modulating root morphology, ROS homeostasis, and chloroplast autophagy. J. Plant Growth Regul. 2022, 1–14. [Google Scholar] [CrossRef]
- Ma, X.; Gu, J.; Luo, Q.; Wen, M.; Li, H.; Wang, Z. The synergistic benefits of β—Aminobutyric acid and γ-aminobutyrate on salt and drought tolerance in cassava. Plant Biotechnol. Rep. 2022, 16, 29–41. [Google Scholar] [CrossRef]
- Mathur, S.; Mehta, P.; Jajoo, A. Effects of dual stress (high salt and high temperature) on the photochemical efficiency of wheat leaves (Triticum aestivum). Physiol. Mol. Biol. Plants 2013, 19, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Szafrańska, K.; Reiter, R.J.; Posmyk, M.M. Melatonin application to Pisum sativum L. seeds positively influences the function of the photosynthetic apparatus in growing seedlings during paraquat-induced oxidative stress. Front. Plant Sci. 2016, 7, 1663. [Google Scholar] [CrossRef] [PubMed]
- Ambreen, S.; Athar, H.; Khan, A.; Zafar, Z.U.; Ayyaz, A.; Kalaji, H.M. Seed priming with proline improved photosystem II efficiency and growth of wheat (Triticum aestivum L.). BMC Plant Biol. 2021, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, T.; Farooq, M.; Ahmad, R.; Zohaib, A.; Wahid, A. Seed Priming and transgenerational drought memory improves tolerance against salt stress in bread wheat. Plant Physiol. Biochem. 2017, 118, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; Puthur, J.T.; Challabathula, D.; Brestič, M. Transgenerational effect of UV-B priming on photochemistry and associated metabolism in rice seedlings subjected to PEG-induced osmotic stress. Photosynthetica 2022, 60, 219–229. [Google Scholar] [CrossRef]
- Jisha, K.C.; Puthur, J.T. Seed priming with beta-amino butyric acid improves abiotic stress tolerance in rice seedlings. Rice Sci. 2016, 23, 242–254. [Google Scholar] [CrossRef]
- Vijayakumari, K.; Jisha, K.C.; Puthur, J.T. GABA/BABA priming: A means for enhancing abiotic stress tolerance potential of plants with less energy investments on defence cache. Acta Physiol. Plant. 2016, 38, 230. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Osmolality (mmol kg−1) |
|---|---|
| C | 292 ± 7.05 |
| B | 335 ± 9.01 |
| P | 338 ± 10.40 |
| S | 382 ± 9.17 |
| B + S | 425 ± 7.64 |
| P + S | 428 ± 8.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aswathi, K.P.R.; Sen, A.; Puthur, J.T. Comparative Study of Cis- and Trans-Priming Effect of PEG and BABA in Cowpea Seedlings on Exposure to PEG-Induced Osmotic Stress. Seeds 2023, 2, 85-100. https://doi.org/10.3390/seeds2010007
Aswathi KPR, Sen A, Puthur JT. Comparative Study of Cis- and Trans-Priming Effect of PEG and BABA in Cowpea Seedlings on Exposure to PEG-Induced Osmotic Stress. Seeds. 2023; 2(1):85-100. https://doi.org/10.3390/seeds2010007
Chicago/Turabian StyleAswathi, K. P. Raj, Akhila Sen, and Jos T. Puthur. 2023. "Comparative Study of Cis- and Trans-Priming Effect of PEG and BABA in Cowpea Seedlings on Exposure to PEG-Induced Osmotic Stress" Seeds 2, no. 1: 85-100. https://doi.org/10.3390/seeds2010007
APA StyleAswathi, K. P. R., Sen, A., & Puthur, J. T. (2023). Comparative Study of Cis- and Trans-Priming Effect of PEG and BABA in Cowpea Seedlings on Exposure to PEG-Induced Osmotic Stress. Seeds, 2(1), 85-100. https://doi.org/10.3390/seeds2010007