
An Indo-West Pacific Distribution for the Coral-Dwelling Gall Crab Lithoscaptus doughnut (Decapoda: Cryptochiridae)



Abstract
:1. Introduction
2. Material and Methods
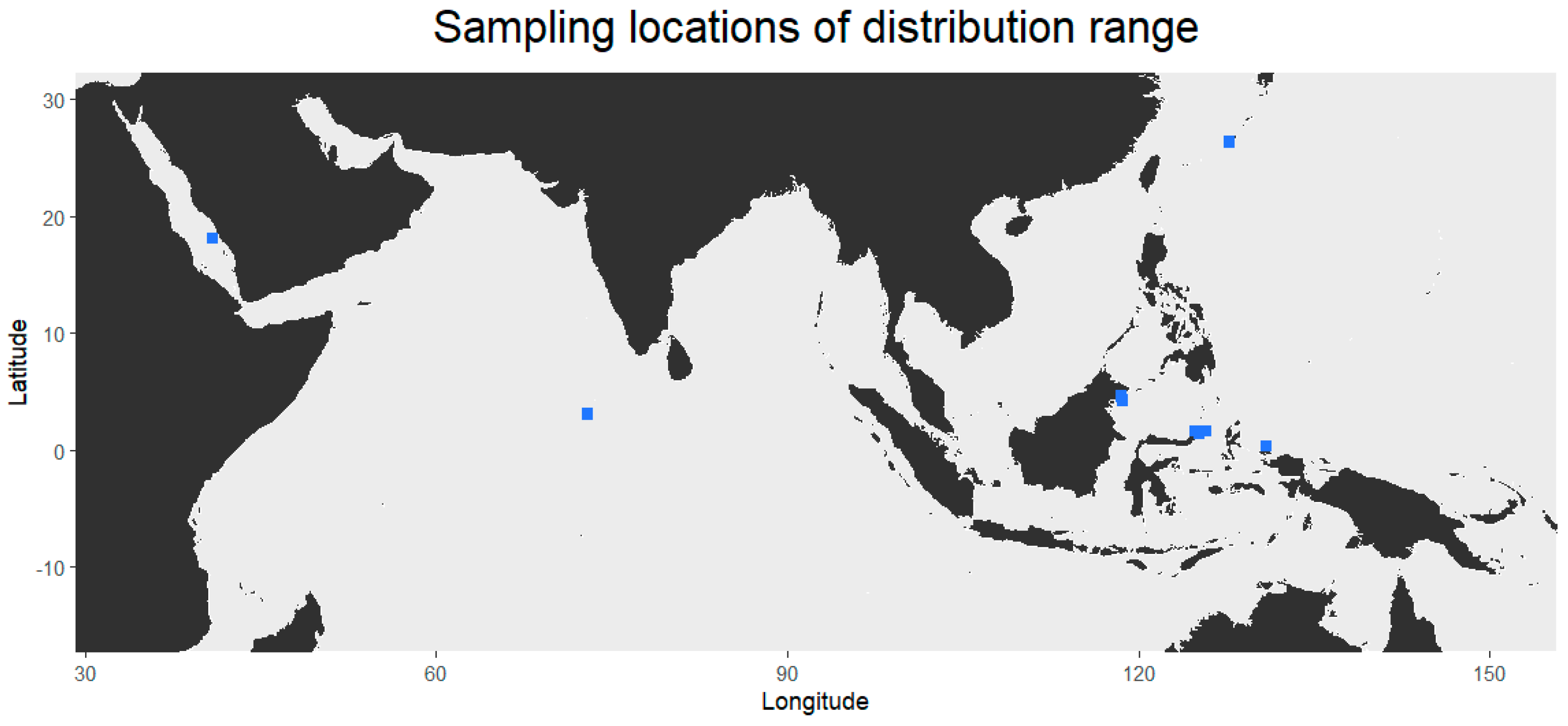
2.1. Sampling
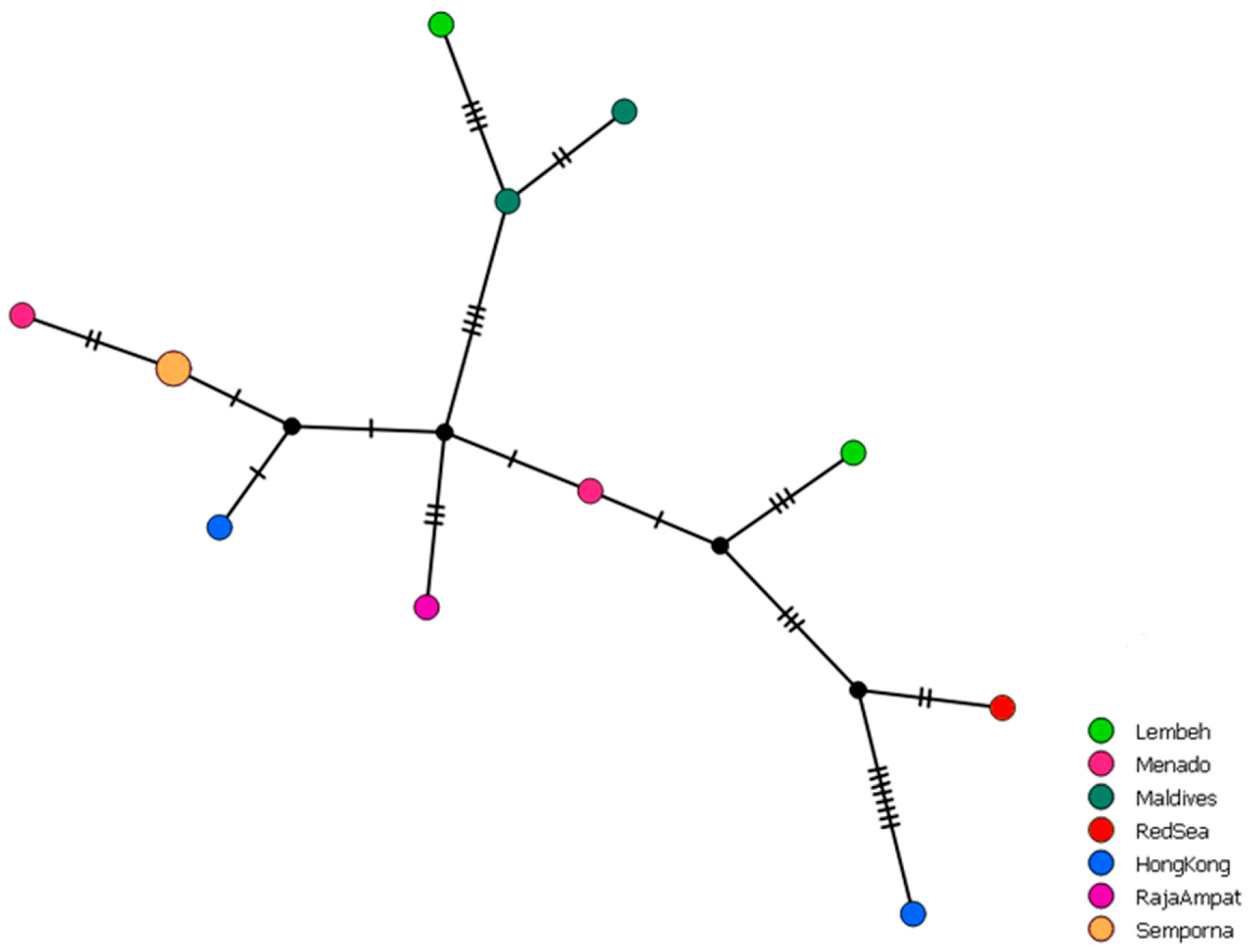
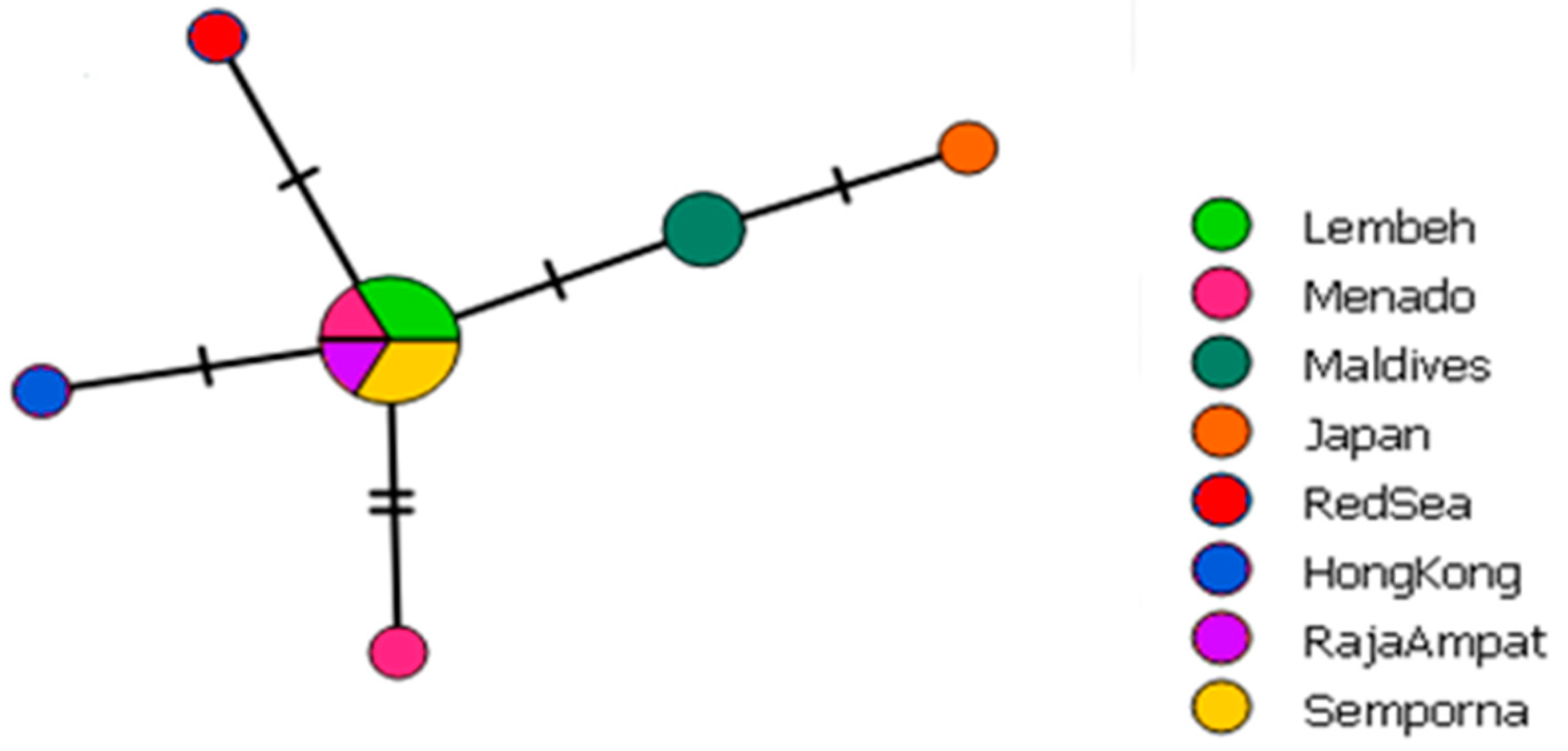
2.2. Haplotype Networks
2.3. Intraspecific Divergence
3. Results and Discussion
3.1. Distribution and Haplotype Networks
3.2. Coral Host
3.3. Phylogenetic Position
3.4. Lithoscaptus cf. doughnut
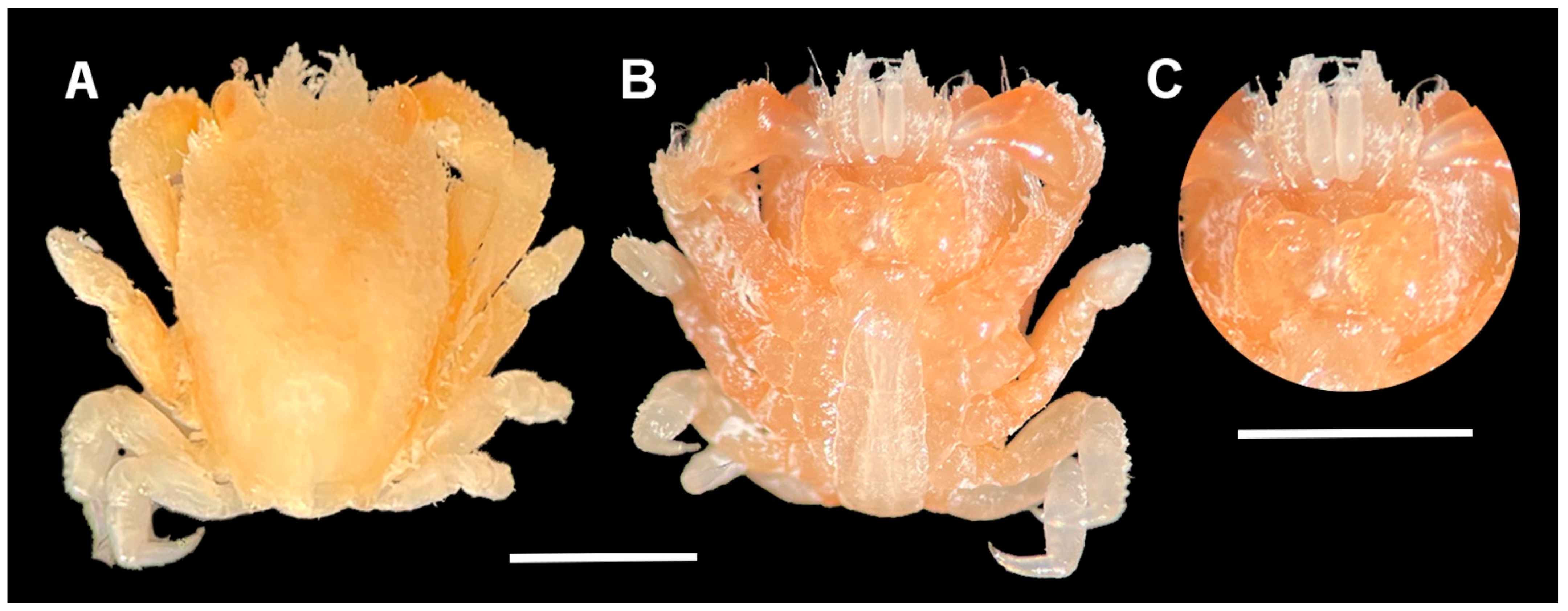
3.5. Colour in Life
3.6. Description of Male L. doughnut
3.7. Parasites
3.8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reaka-Kudla, M.L. The Global Biodiversity of Coral Reefs: A Comparison with Rain Forests. In Biodiversity II: Understanding and Protecting Our Biological Resources; Reaka-Kudla, M.L., Wilson, D.E., Wilson, E.O., Eds.; Joseph Henry Press: Washington, WA, USA, 1997; pp. 83–108. [Google Scholar]
- Hoeksema, B.W.; van der Meij, S.E.T.; Fransen, C.H.J.M. The Mushroom Coral as a Habitat. J. Mar. Biol. Assoc. 2012, 92, 647–663. [Google Scholar] [CrossRef]
- Stella, J.S.; Pratchett, M.S.; Hutchings, P.A.; Jones, G.P. Coral-Associated Invertebrates: Diversity, Ecological Importance and Vulnerability to Disturbance. Oceanogr. Mar. Biol. Annu. Rev. 2011, 49, 43–104. [Google Scholar]
- Vehof, J.; van der Meij, S.E.T.; Türkay, M.; Becker, C. Female Reproductive Morphology of Coral-Inhabiting Gall Crabs (Crustacea: Decapoda: Brachyura: Cryptochiridae). Acta Zool. 2016, 97, 117–126. [Google Scholar] [CrossRef]
- van Tienderen, K.M.; van der Meij, S.E.T. Extreme Mitochondrial Variation in the Atlantic Gall Crab Opecarcinus hypostegus (Decapoda Cryptochiridae) Reveals Adaptive Genetic Divergence over Agaricia Coral Hosts. Sci. Rep. 2017, 7, 39461. [Google Scholar] [CrossRef] [PubMed]
- Bähr, S.; van der Meij, S.E.T. Red Fluorescence in Symbiotic Coral-Dwelling Gall Crabs. Galaxea J. Coral Reef Stud. 2019, 21, 27–28. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Bravo, H.; Paulay, G.; van der Meij, S.E.T. Diversification and Distribution of Gall Crabs (Brachyura: Cryptochiridae: Opecarcinus) Associated with Agariciidae Corals. Coral Reefs 2022, 41, 699–709. [Google Scholar] [CrossRef]
- Bähr, S.; van der Meij, S.E.T.; Terraneo, T.I.; Xu, T.; Benzoni, F. Interspecific Coral Competition Does Not Affect the Symbiosis of Gall Crabs (Decapoda: Cryptochiridae) and Their Scleractinian Hosts. Ecol. Evol. 2023, 13, e10051. [Google Scholar] [CrossRef]
- Kropp, R.K.; Manning, R.B. The Atlantic Gall Crabs, Family Cryptochiridae (Crustacea: Decapoda: Brachyura). Smithson. Contrib. Zool. 1987, 462, 1–11. [Google Scholar] [CrossRef]
- DecaNet Eds. DecaNet. 2023. Available online: https://www.decanet.info (accessed on 3 October 2023). [CrossRef]
- Bähr, S.; Johnson, M.L.; Berumen, M.L.; Hardenstine, R.S.; Rich, W.A.; van der Meij, S.E.T. Morphology and Reproduction in the Hapalocarcinus marsupialis Stimpson, 1859 Species Complex (Decapoda: Brachyura: Cryptochiridae). J. Crustac. Biol. 2021, 41, ruab052. [Google Scholar] [CrossRef]
- Fize, A.; Serène, R. Les Hapalocarcinidés du Viet-Nam. Archives du Museum National d’Histoire Naturelle; Sèptieme Série. 5; Archives de France: Paris, France, 1957; pp. 1–202. [Google Scholar]
- Kropp, R.K. Revision of the Genera of Gall Crabs (Crustacea: Cryptochiridae) Occurring in the Pacific Ocean. Pac. Sci. 1990, 44, 417–448. [Google Scholar]
- van der Meij, S.E.T. Host Relations and DNA Reveal a Cryptic Gall Crab Species (Crustacea: Decapoda: Cryptochiridae) Associated with Mushroom Corals (Scleractinia: Fungiidae). Contrib. Zool. 2015, 84, 39–57. [Google Scholar] [CrossRef]
- van der Meij, S.E.T. Host Species, Range Extensions, and an Observation of the Mating System of Atlantic Shallow-Water Gall Crabs (Decapoda: Cryptochiridae). Bull. Mar. Sci. 2014, 90, 1001–1010. [Google Scholar] [CrossRef]
- Huang, D.; Goldberg, E.E.; Chou, L.M.; Roy, K. The Origin and Evolution of Coral Species Richness in a Marine Biodiversity Hotspot. Evolution 2018, 72, 288–302. [Google Scholar] [CrossRef]
- Briggs, J.C.; Bowen, B.W. A Realignment of Marine Biogeographic Provinces with Particular Reference to Fish Distributions. J. Biogeogr. 2012, 39, 12–30. [Google Scholar] [CrossRef]
- van der Meij, S.E.T.; Berumen, M.L.; Paulay, G. A New Species of Fizesereneia Takeda & Tamura, 1980 (Crustacea: Brachyura: Cryptochiridae) from the Red Sea and Oman. Zootaxa 2015, 3931, 585–595. [Google Scholar] [CrossRef] [PubMed]
- van der Meij, S.E.T. A New Gall Crab Species (Brachyura, Cryptochiridae) Associated with the Free-Living Coral Trachyphyllia geoffroyi (Scleractinia, Merulinidae). Zookeys 2015, 500, 61–72. [Google Scholar] [CrossRef]
- van der Meij, S.E.T. The Coral Genus Caulastraea Dana, 1846 (Scleractinia, Merulinidae) as a New Host for Gall Crabs (Decapoda, Cryptochiridae), with the Description of Lithoscaptus tuerkayi sp. nov. Crustaceana 2017, 90, 1027–1038. [Google Scholar] [CrossRef]
- Kropp, R.K. The Status of Cryptochirus coralliodytes Heller and Lithoscaptus paradoxus Milne-Edwards (Brachyura: Cryptochiridae). Proc. Biol. Soc. Wash. 1988, 101, 872–882. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Schubart, C.D. Mitochondrial DNA and Decapod Phylogenies: The Importance of Pseudogenes and Primer Optimization. In Crustacean Issues 18: Decapod Crustacean Phylogenetics; Martin, J.W., Crandall, K.A., Felder, D.L., Eds.; CRC Press: Boca Raton, FL, USA, 2009; pp. 47–65. [Google Scholar]
- Sequencher; Version 4.10.1 DNA Sequence Analysis Software; Gene Codes Coorporation: Ann Arbor, MI, USA, 2010; Available online: http://www.genecodes.com/ (accessed on 21 December 2023).
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Wong, K.J.H.; Tsao, Y.F.; Qiu, J.W.; Chan, B.K.K. Diversity of Coral-Associated Pit Crabs (Crustacea: Decapoda: Cryptochiridae) from Hong Kong, with Description of Two New Species of Lithoscaptus A. Milne-Edwards, 1862. Front. Mar. Sci. 2023, 9, 1003321. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-Feature Software for Haplotype Network Construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Juszkiewicz, D.J.; White, N.E.; Stolarski, J.; Benzoni, F.; Arrigoni, R.; Hoeksema, B.W.; Wilson, N.G.; Bunce, M.; Richards, Z.T. Phylogeography of Recent Plesiastrea (Scleractinia: Plesiastreidae) Based on an Integrated Taxonomic Approach. Mol. Phylogenetics Evol. 2022, 172, 107469. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; van der Meij, S.E.T. Cryptic Marine Biota of the Raja Ampat Island Group; Naturalis Biodiversity Center: Leiden, The Netherlands, 2008. [Google Scholar]
- Boyko, C.B.; van der Meij, S.E.T. A Trifecta of Swiftian Symbioses: Stony Corals, Gall Crabs and Their Parasites (Scleractinia; Brachyura: Cryptochiridae; Isopoda: Epicaridea and Cirripedia: Rhizocephala). Zool. J. Linn. Soc. 2018, 184, 304–329. [Google Scholar] [CrossRef]
- Norton, D.A.; Carpenter, M.A. Mistletoes as Parasites: Host Specificity and Speciation. Trends Ecol. Evol. 1998, 13, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Quek, Z.B.R.; Jain, S.S.; Richards, Z.T.; Arrigoni, R.; Benzoni, F.; Hoeksema, B.W.; Carvajal, J.I.; Wilson, N.G.; Baird, A.H.; Kitahara, M.V.; et al. A Hybrid-Capture Approach to Reconstruct the Phylogeny of Scleractinia (Cnidaria: Hexacorallia). Mol. Phylogenetics Evol. 2023, 186, 107867. [Google Scholar] [CrossRef] [PubMed]
- van der Meij, S.E.T.; Klaus, S. Origin and Diversification of Coral-Dwelling Gall Crabs (Decapoda: Cryptochiridae) In Evolutionary Diversification of Coral-dwelling Gall Crabs (Cryptochiridae). Ph.D. Thesis, Naturalis Biodiversity Center & Leiden University, Leiden, The Netherlands, 2015; pp. 79–86. [Google Scholar]
- van der Meij, S.E.T. Lithoscaptus aquarius sp. nov (Decapoda: Cryptochiridae) Described from a Catalaphyllia jardinei (Scleractinia) out of the Aquarium Trade. Arthropoda 2023, 1, 350–359. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Voucher Code | COI | 16S | Host Coral | Country | Locality |
|---|---|---|---|---|---|
| CEL-Hapa-040/ASIZCR | OP103644 | n/a | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Hong Kong | Basalt Island, Sai Kung |
| CEL-Hapa-006 | OP103613 | OP114856 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Hong Kong | Long Ke Tsai, Sai Kung |
| RMNH.CRUS.D.54169 | OR710957 | OR711069 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Indonesia | Tanjung Nanas II, Lembeh Isl., N Sulawesi |
| RMNH.CRUS.D.54172 | OR710956 | OR711070 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Indonesia | Pulau Abadi, Lembeh Isl., N Sulawesi |
| RMNH.CRUS.D.54106 | OR710954 | OR711068 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Indonesia | Lekuan III, Bunaken Isl., N Sulawesi |
| RMNH.CRUS.D.54063 | OR710953 | OR711067 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Indonesia | Tiwoho, Bunaken Isl., N Sulawesi |
| RMNH.CRUS.D.53890 | OR710955 | OR711066 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Indonesia | Mayalibit Bay, E Manil Isl., Raja Ampat, W Papua |
| RMNH.CRUS.D.57236 | NA | OR711074 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Japan | Mizugama, Okinawa |
| RMNH.CRUS.D.53725 | OR710951 | OR711064 | Cyphastrea chalcidicum (Forskål, 1775) | Malaysia | Mabul Isl., Eel Garden, Semporna |
| RMNH.CRUS.D.53740 | OR710952 | OR711065 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Malaysia | Mata Pahi Isl., Semporna |
| RMNH.CRUS.D.58330 | OR710958 | OR711072 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Maldives | Free Climbing, Faafu Atoll |
| RMNH.CRUS.D.58331 | OR710959 | OR711073 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Maldives | Route 66, Faafu Atoll |
| RMNH.CRUS.D.54404 | OR710960 | OR711071 | Plesiastrea peroni, Milne Edwards & Haime, 1857 | Saudi Arabia | Shib Radib, offshore of Farasan Banks |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Claassen, J.R.; Tuti, Y.; van der Meij, S.E.T. An Indo-West Pacific Distribution for the Coral-Dwelling Gall Crab Lithoscaptus doughnut (Decapoda: Cryptochiridae). Arthropoda 2024, 2, 66-75. https://doi.org/10.3390/arthropoda2010005
Claassen JR, Tuti Y, van der Meij SET. An Indo-West Pacific Distribution for the Coral-Dwelling Gall Crab Lithoscaptus doughnut (Decapoda: Cryptochiridae). Arthropoda. 2024; 2(1):66-75. https://doi.org/10.3390/arthropoda2010005
Chicago/Turabian StyleClaassen, Jorn R., Yosephine Tuti, and Sancia E. T. van der Meij. 2024. "An Indo-West Pacific Distribution for the Coral-Dwelling Gall Crab Lithoscaptus doughnut (Decapoda: Cryptochiridae)" Arthropoda 2, no. 1: 66-75. https://doi.org/10.3390/arthropoda2010005
APA StyleClaassen, J. R., Tuti, Y., & van der Meij, S. E. T. (2024). An Indo-West Pacific Distribution for the Coral-Dwelling Gall Crab Lithoscaptus doughnut (Decapoda: Cryptochiridae). Arthropoda, 2(1), 66-75. https://doi.org/10.3390/arthropoda2010005