Characterizing Complex Networks Using Entropy-Degree Diagrams: Unveiling Changes in Functional Brain Connectivity Induced by Ayahuasca

 and
and 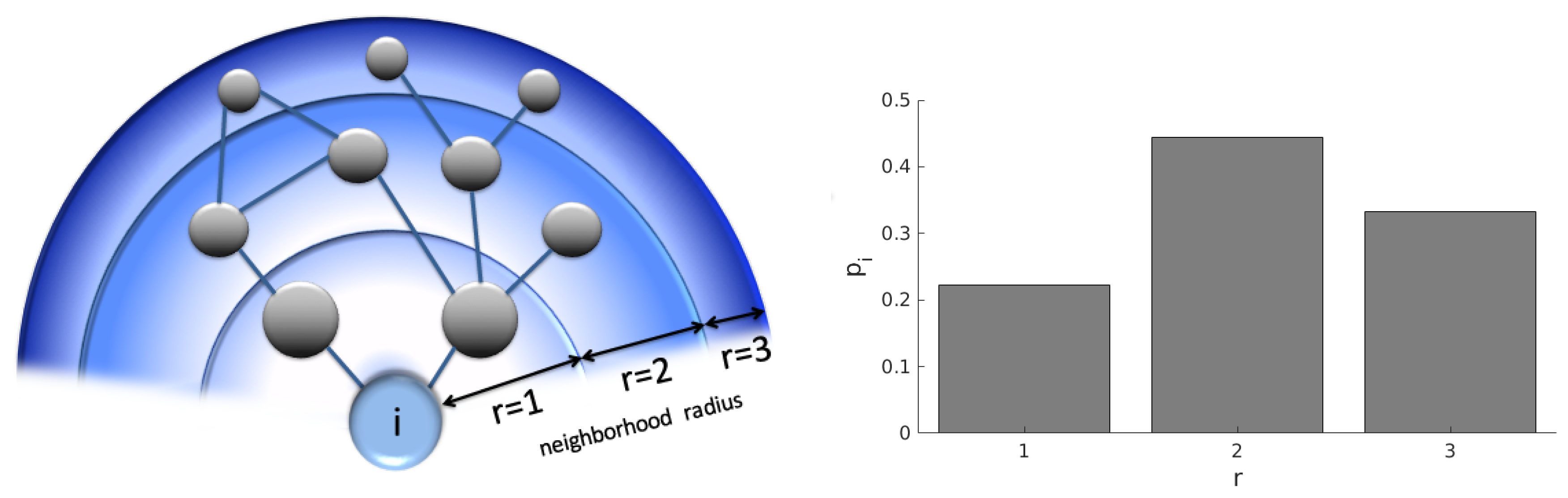{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
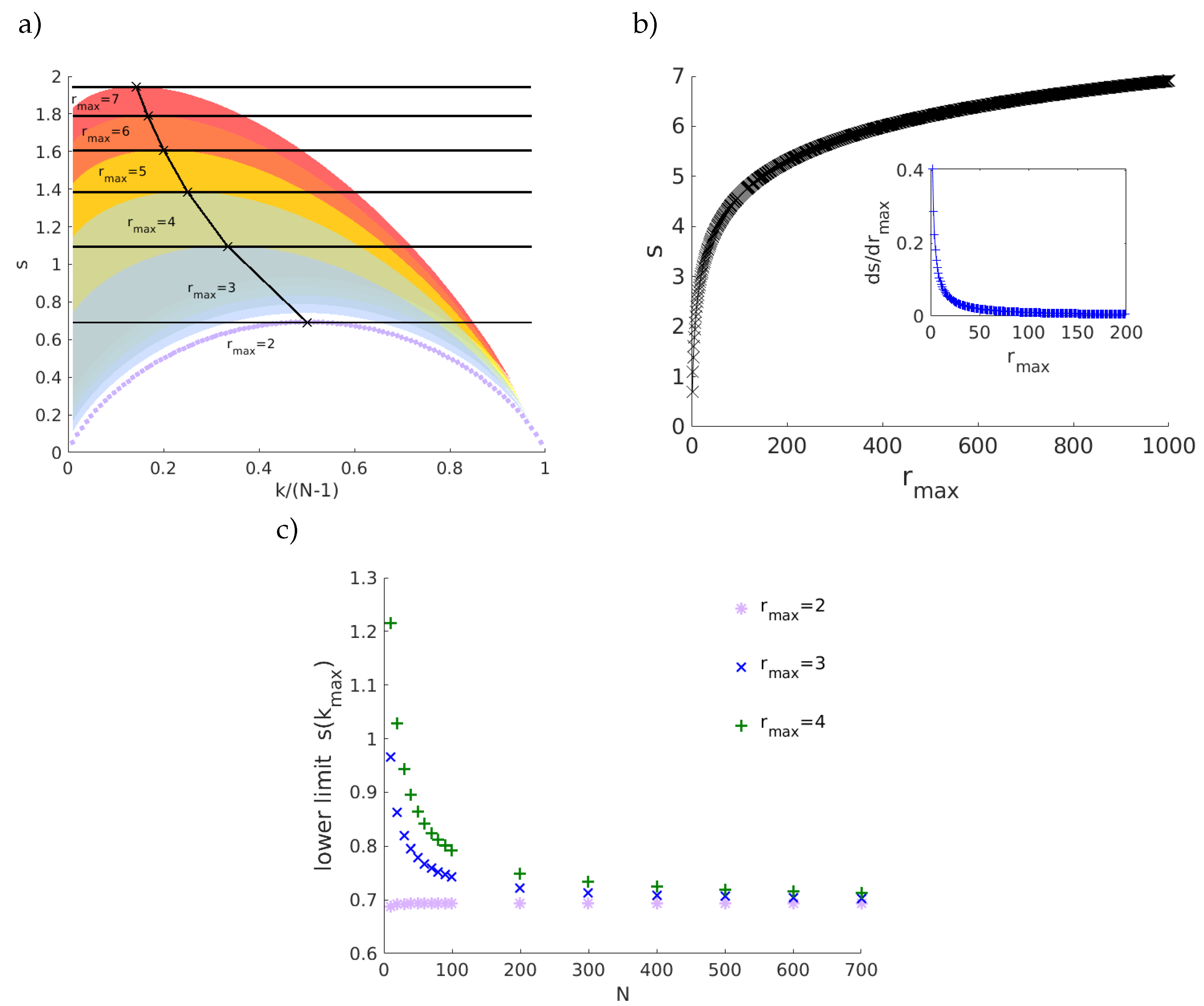
2. Geodesic Entropy
Entropy-Degree Diagram
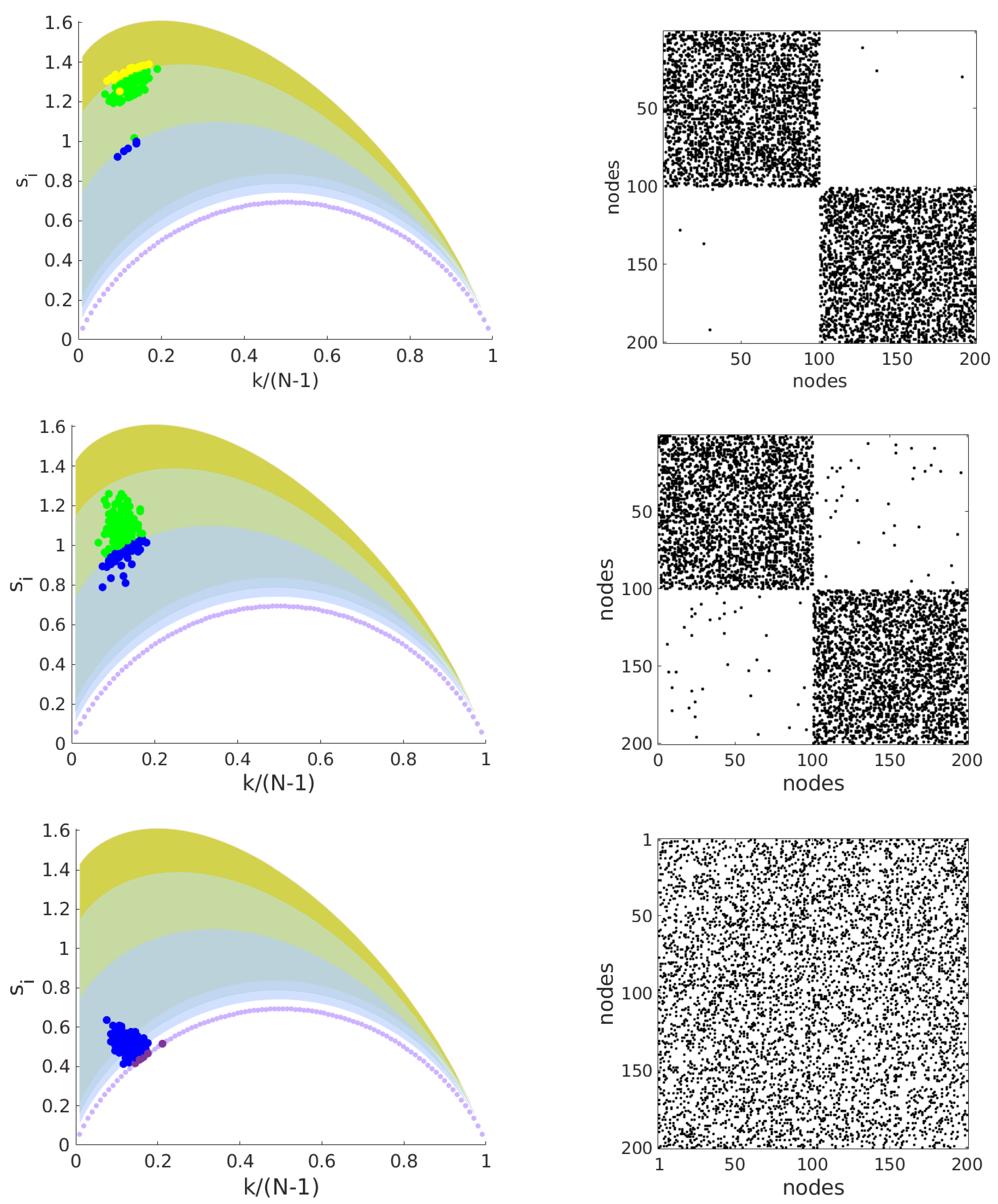
3. Results
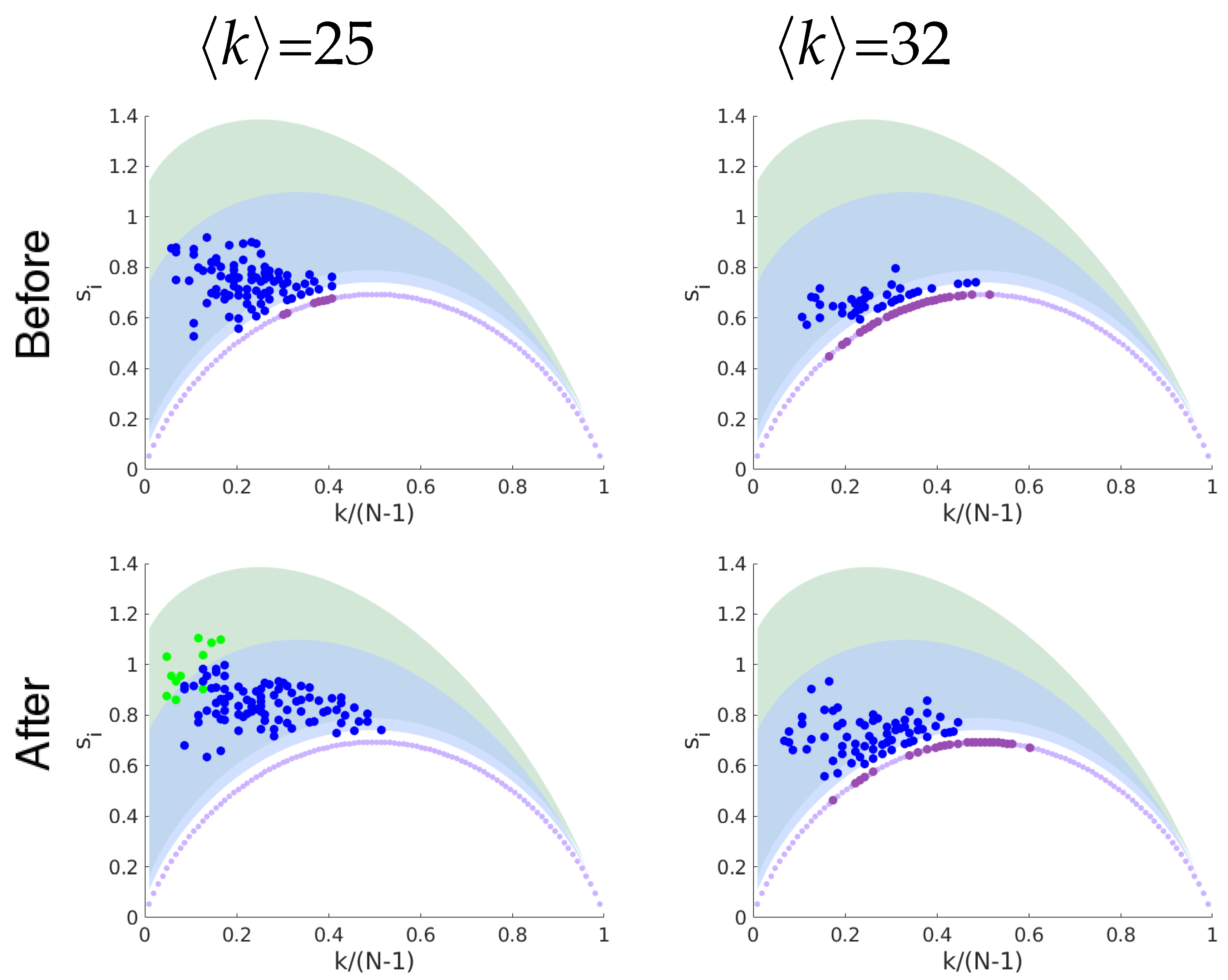
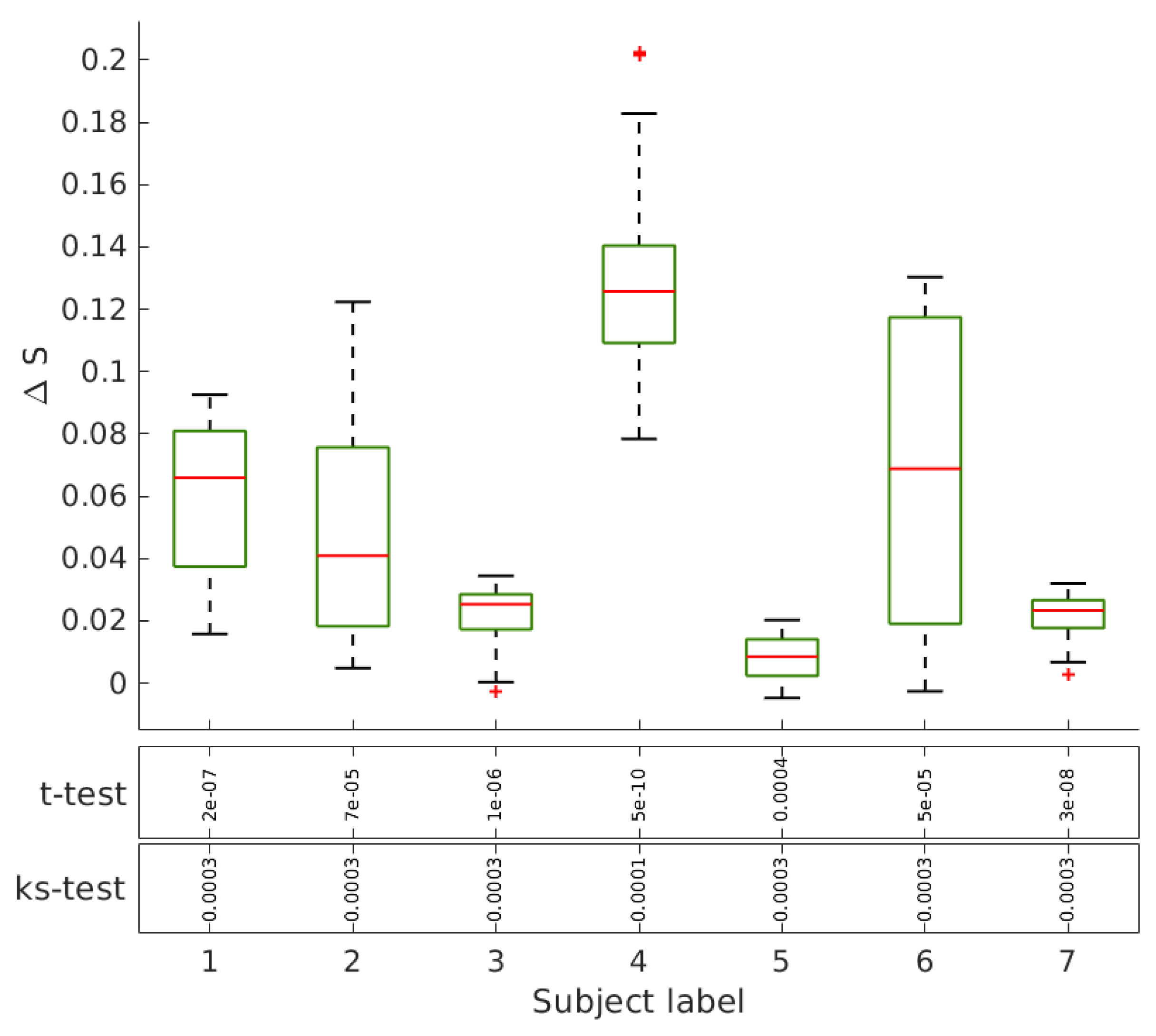
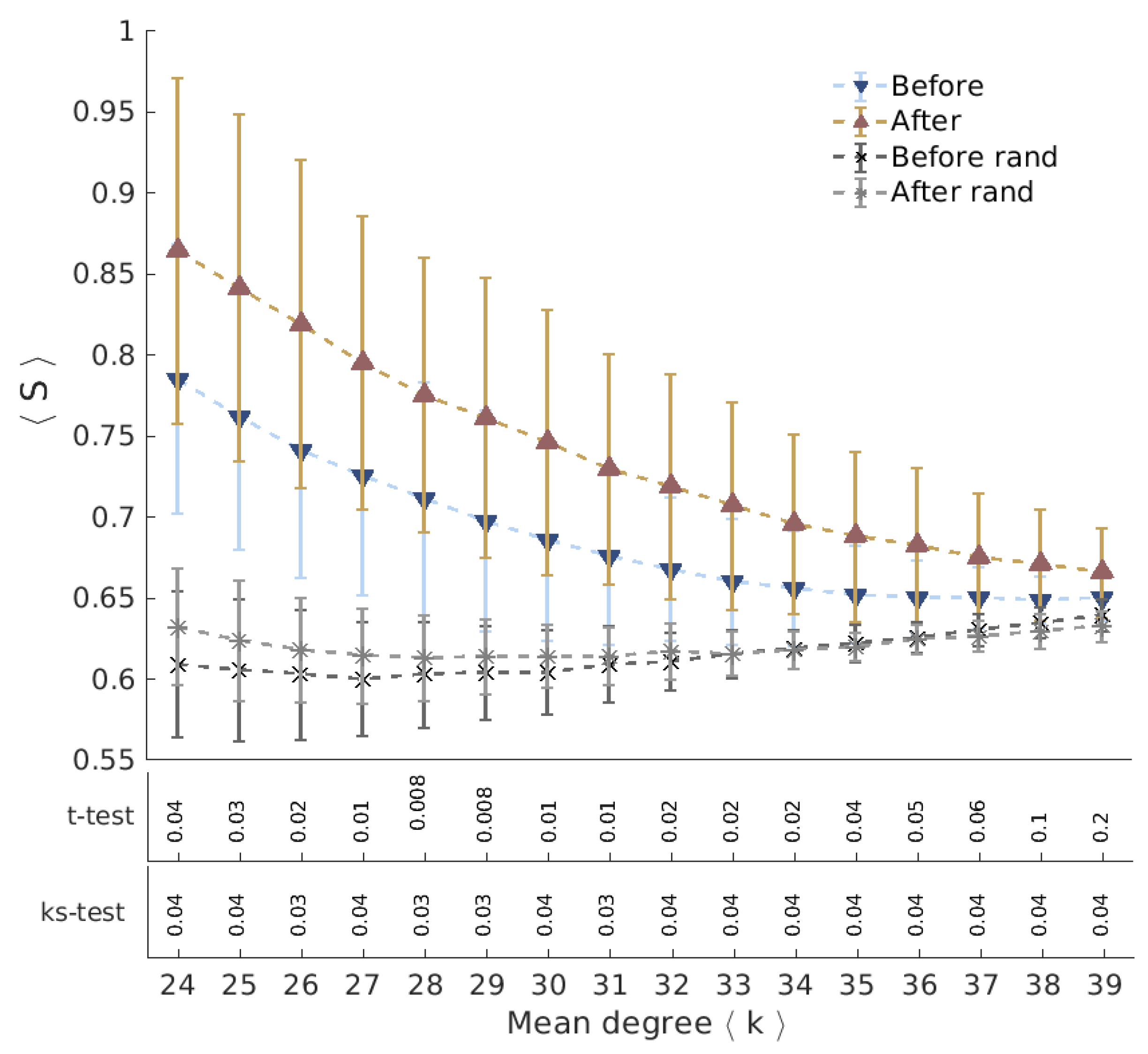
Geodesic Entropy of Functional Brain Networks under Ayahuasca Influence
4. Discussion and Conclusions
5. Materials and Methods
5.1. Data
5.2. Obtaining Functional Networks from fMRI Data
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bassett, D.S.; Sporns, O. Network neuroscience. Nat. Neurosci. 2017, 20, 353. [Google Scholar] [CrossRef] [PubMed]
- Borge-Holthoefer, J.; Baños, R.A.; González-Bailón, S.; Moreno, Y. Cascading behaviour in complex socio-technical networks. J. Complex Netw. 2013, 1, 3–24. [Google Scholar] [CrossRef] [Green Version]
- Pilosof, S.; Porter, M.A.; Pascual, M.; Kéfi, S. The multilayer nature of ecological networks. Nat. Ecol. Evol. 2017, 1, 0101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haynes, J.D.; Rees, G. Decoding mental states from brain activity in humans. Nat. Rev. Neurosci. 2006, 7, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.E.J. A measure of betweenness centrality based on random walks. Soc. Netw. 2005, 27, 39–54. [Google Scholar] [CrossRef] [Green Version]
- van den Heuvel, M.P.; Sporns, O. Network hubs in the human brain. Trends Cogn. Sci. 2013, 17, 683–696. [Google Scholar] [CrossRef]
- Hou, B.; Yao, Y.; Liao, D. Identifying all-around nodes for spreading dynamics in complex networks. Physica A 2012, 391, 4012–4017. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Riba, J.; Valle, M.; Urbano, G.; Yritia, M.; Morte, A.; Barbanoj, M.J. Human Pharmacology of Ayahuasca: Subjective and Cardiovascular Effects, Monoamine Metabolite Excretion, and Pharmacokinetics. J. Pharmacol. Exp. Ther. 2003, 306, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, A.; Ott, J.; Feilding, A. LSD: My Problem Child; OUP Oxford: Oxford, UK, 2013. [Google Scholar]
- Passie, T.; Halpern, J.H.; Stichtenoth, O.D.; Emrich, H.M.; Hintzen, A. The Pharmacology of Lysergic Acid Diethylamide: A Review. CNS Neurosci. Ther. 2018, 14, 295–314. [Google Scholar] [CrossRef] [PubMed]
- Shanon, B. The Antipodes of the Mind: Charting the Phenomenology of the Ayahuasca Experience; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Labate, B.C.; Cavnar, C. Ayahuasca Shamanism in the Amazon and Beyond; Oxford Ritual Studies Series; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Labate, B.C.; Cavnar, C. Prohibition, Religious Freedom, and Human Rights: Regulating Traditional Drug Use; Springer: Berlin, Germany, 2014. [Google Scholar]
- Labate, B.C.; Cavnar, C. The Therapeutic Use of Ayahuasca; Springer: Berlin, Germany, 2013. [Google Scholar]
- Riba, J.; Rodriguez-Fornells, A.; Urbano, G.; Morte, A.; Antonijoan, R.; Montero, M.; Callaway, C.J.; Barbanoj, J.M. Subjective effects and tolerability of the South American psychoactive beverage Ayahuasca in healthy volunteers. Psychopharmacology 2001, 154, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Shanon, B. Antipodes of the Mind; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Newman, M.E.J. Networks: An Introduction; Oxford University Press, Inc.: New York, NY, USA, 2010. [Google Scholar]
- Albert, R.; Barabasi, A.L. Statistical mechanics of complex networks. Rev. Mod. Phys. 2002, 74, 47–97. [Google Scholar] [CrossRef] [Green Version]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. NeuroImage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- Watts, D.J.; Strogatz, S.H. Collective dynamics of ’small-world’ networks. Nature 1998, 393, 440. [Google Scholar] [CrossRef] [PubMed]
- Viol, A.; Palhano-Fontes, F.; Onias, H.; de Araujo, D.B.; Viswanathan, G.M. Shannon entropy of brain functional complex networks under the influence of the psychedelic Ayahuasca. Sci. Rep. 2017, 7, 7388. [Google Scholar] [CrossRef] [PubMed]
- Maslov, S.; Sneppen, K. Specificity and Stability in Topology of Protein Networks. Science 2002, 296, 910–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z. Pattern Synchronization of Nonlinear Heterogeneous Multiagent Networks With Jointly Connected Topologies. IEEE Trans. Control Netw. Syst. 2014, 1, 349–359. [Google Scholar] [CrossRef]
- Stella, M.; De Domenico, M. Distance Entropy Cartography Characterises Centrality in Complex Networks. Entropy 2018, 20, 268. [Google Scholar] [CrossRef]
- Carhart-Harris, R.; Leech, R.; Hellyer, P.J.; Shanahan, M.; Feilding, A.; Tagliazucchi, E.; Chialvo, D.R.; Nutt, D. The entropic brain: A theory of conscious states informed by neuroimaging research with psychedelic drugs. Front. Hum. Neurosci. 2014, 8, 20. [Google Scholar]
- Papo, D. Commentary: The entropic brain: a theory of conscious states informed by neuroimaging research with psychedelic drugs. Front. Hum. Neurosci. 2016, 10, 423. [Google Scholar] [CrossRef]
- Carhart-Harris, R.L. The entropic brain—Revisited. Neuropharmacology 2018, 142, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Association, A.P.; DSM-IV-TR, American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-IV-TR. In Diagnostic and Statistical Manual of Mental Disorders; American Psychiatric Association Publishing: Washington, DC, USA, 2000. [Google Scholar]
- De Araujo, D.B.; Ribeiro, S.; Cecchi, G.A.; Carvalho, F.M.; Sanchez, T.A.; Pinto, J.P.; de Martinis, B.S.; Crippa, J.A.; Hallak, J.E.C.; Santos, A.C. Seeing with the eyes shut: Neural basis of enhanced imagery following ayahuasca ingestion. Hum. Brain Mapp. 2012, 33, 2550–2560. [Google Scholar] [CrossRef] [PubMed]
- Fransson, P. Spontaneous low-frequency BOLD signal fluctuations: An fMRI investigation of the resting-state default mode of brain function hypothesis. Hum. Brain Mapp. 2005, 26, 15–29. [Google Scholar] [CrossRef]
- Onias, H.; Viol, A.; Palhano-Fontes, F.; Andrade, K.C.; Sturzbecher, M.; Viswanathan, G.; de Araujo, D.B. Brain complex network analysis by means of resting state fMRI and graph analysis: Will it be helpful in clinical epilepsy? Epilepsy Behav. 2014, 38, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Schroter, M.S.; Spoormaker, V.I.; Schorer, A.; Wohlschlager, A.; Czisch, M.; Kochs, E.F.; Zimmer, C.; Hemmer, B.; Schneider, G.; Jordan, D.; et al. Spatiotemporal Reconfiguration of Large-Scale Brain Functional Networks during Propofol-Induced Loss of Consciousness. J. Neurosci. 2012, 32, 12832–12840. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liang, M.; Zhou, Y.; He, Y.; Hao, Y.; Song, M.; Yu, C.; Liu, H.; Liu, Z.; Jiang, T. Disrupted small-world networks in schizophrenia. Brain 2008, 131, 945–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viol, A.; Palhano-Fontes, F.; Onias, H.; de Araujo, D.B.; Hövel, P.; Viswanathan, G.M. Characterizing Complex Networks Using Entropy-Degree Diagrams: Unveiling Changes in Functional Brain Connectivity Induced by Ayahuasca. Entropy 2019, 21, 128. https://doi.org/10.3390/e21020128
Viol A, Palhano-Fontes F, Onias H, de Araujo DB, Hövel P, Viswanathan GM. Characterizing Complex Networks Using Entropy-Degree Diagrams: Unveiling Changes in Functional Brain Connectivity Induced by Ayahuasca. Entropy. 2019; 21(2):128. https://doi.org/10.3390/e21020128
Chicago/Turabian StyleViol, Aline, Fernanda Palhano-Fontes, Heloisa Onias, Draulio B. de Araujo, Philipp Hövel, and Gandhi M. Viswanathan. 2019. "Characterizing Complex Networks Using Entropy-Degree Diagrams: Unveiling Changes in Functional Brain Connectivity Induced by Ayahuasca" Entropy 21, no. 2: 128. https://doi.org/10.3390/e21020128
APA StyleViol, A., Palhano-Fontes, F., Onias, H., de Araujo, D. B., Hövel, P., & Viswanathan, G. M. (2019). Characterizing Complex Networks Using Entropy-Degree Diagrams: Unveiling Changes in Functional Brain Connectivity Induced by Ayahuasca. Entropy, 21(2), 128. https://doi.org/10.3390/e21020128