Entropy Production and Its Application to the Coupled Nonequilibrium Processes of ATP Synthesis

Abstract
:1. Introduction
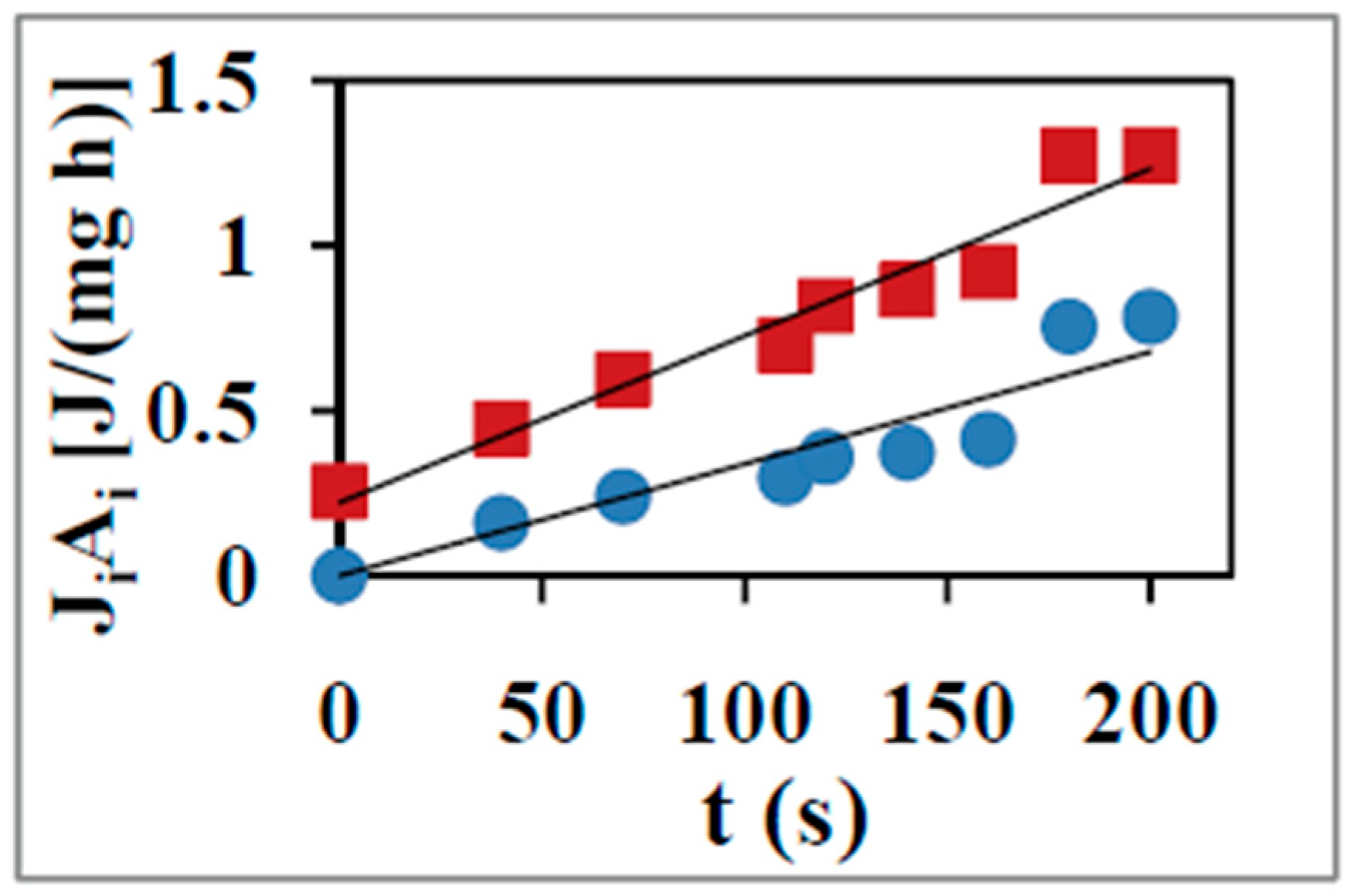
2. Entropy Production During Redox-Coupled ATP Synthesis
3. Calculation of the Thermodynamic Efficiency, η, from the Rate of Entropy Production
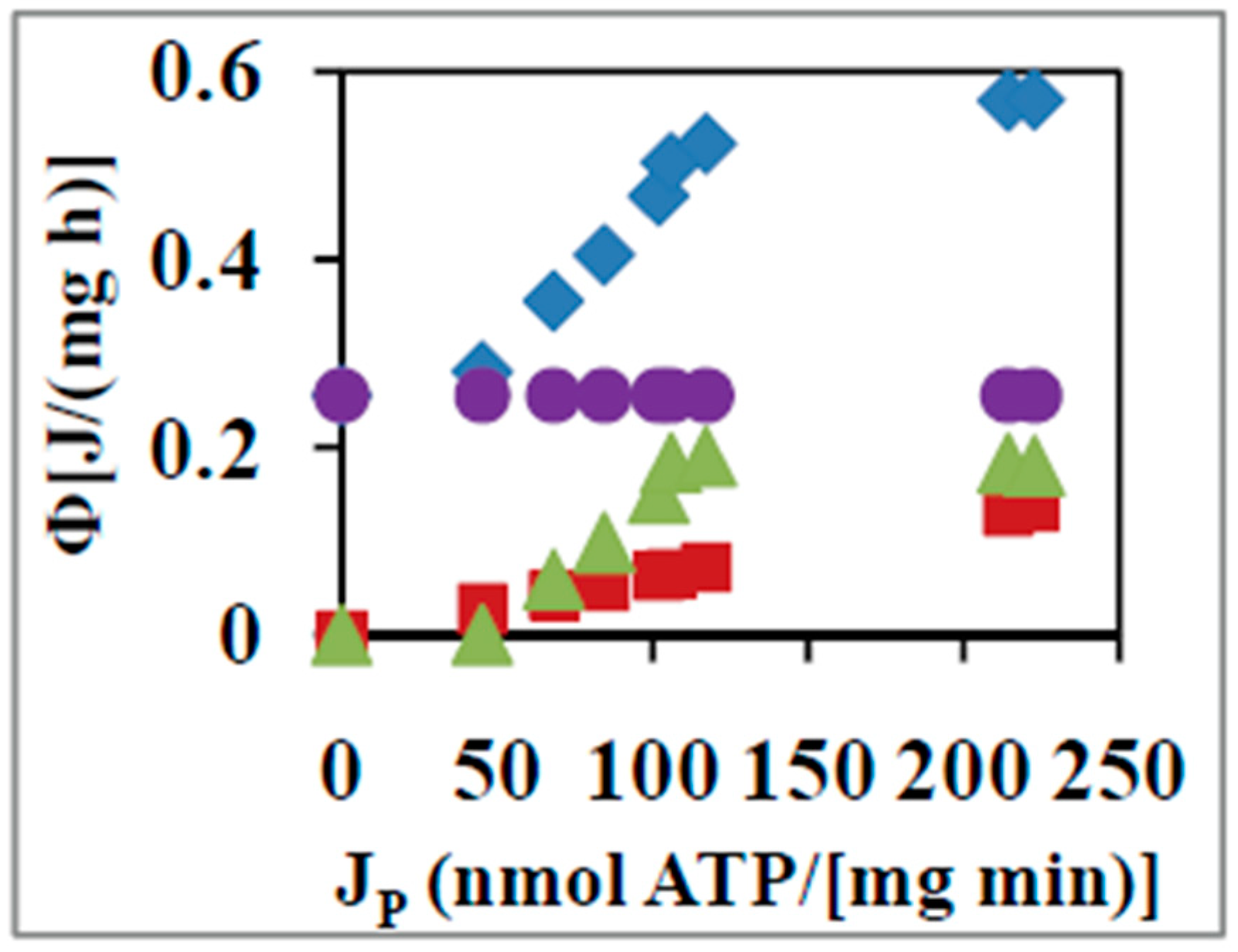
4. Linear Nonequilibrium Thermodynamic Analysis of ATP Synthesis
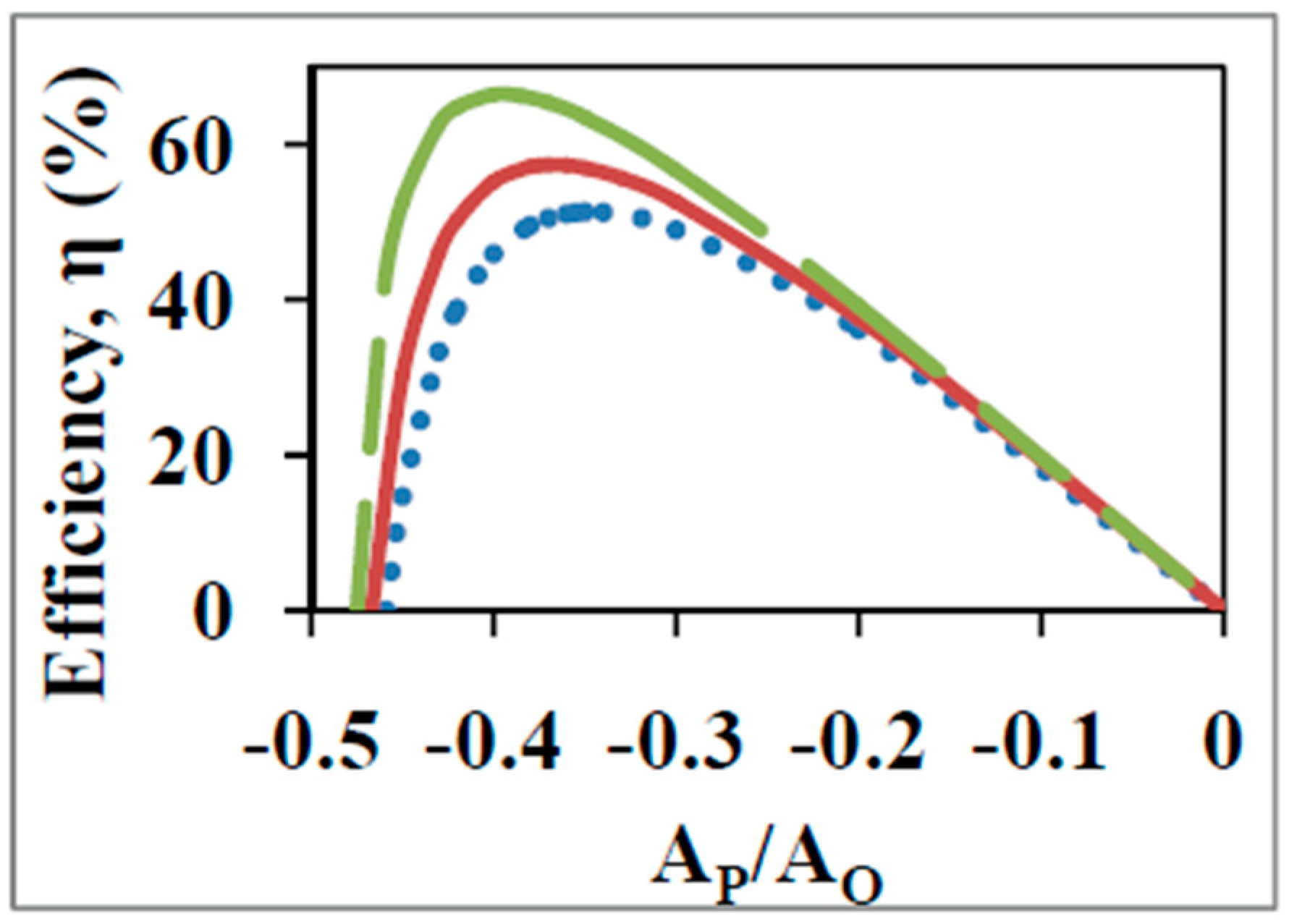
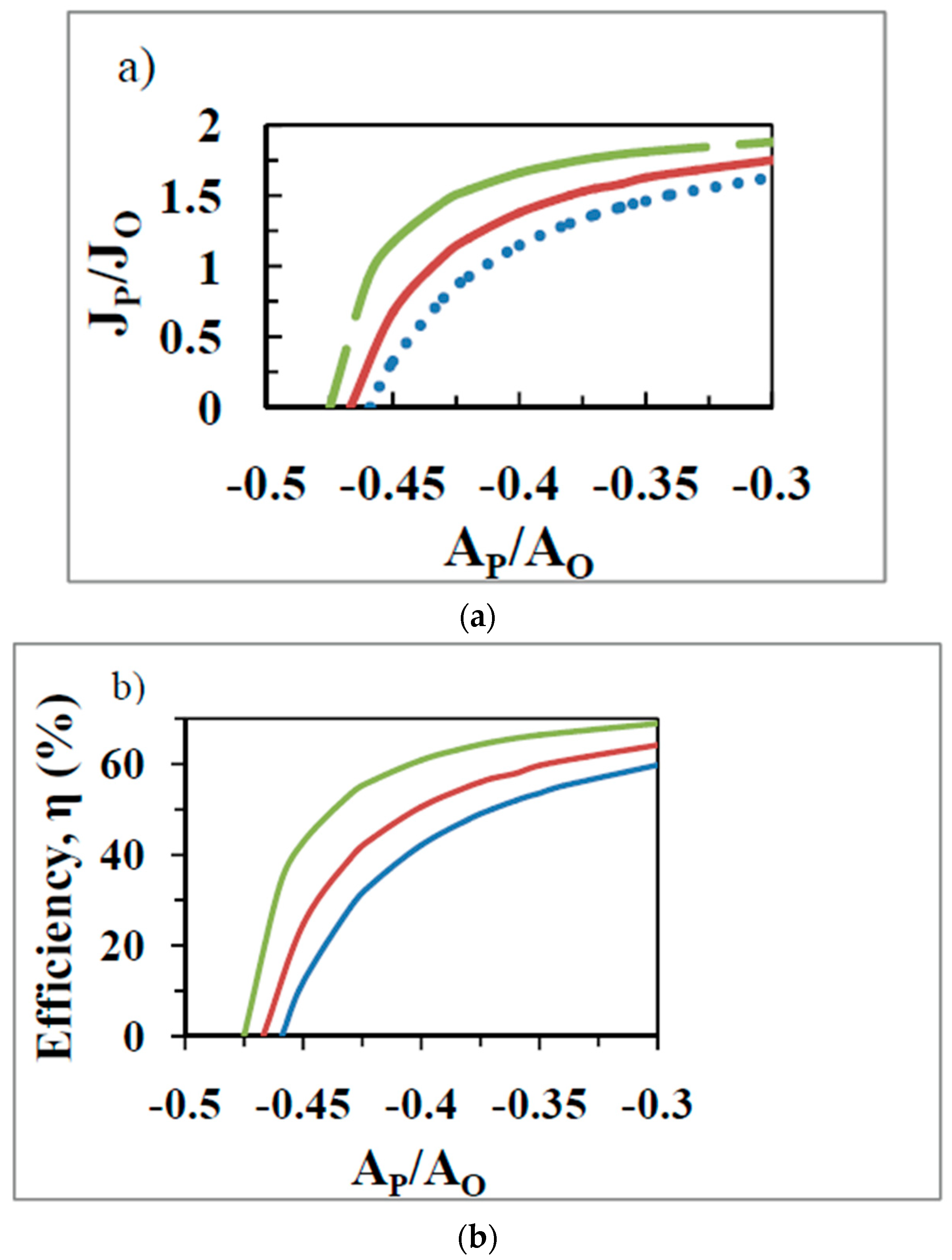
5. Derivation of the Degree of Coupling, , between Oxidation and ATP Synthesis from Experimental Data Using Nonequilibrium Thermodynamics
6. Derivation of a Relationship for ADP/([Extra O]) in Terms of Kinetic Parameters and Comparison with Experimental Results
7. Optimality of Biological Free Energy Converters in OX PHOS
7.1. Biothermodynamics Approach to Optimality Criteria for Free Energy Converters in OX PHOS
7.2. Biothermokinetics Approach to Optimality Criteria for Free Energy Converters in OX PHOS
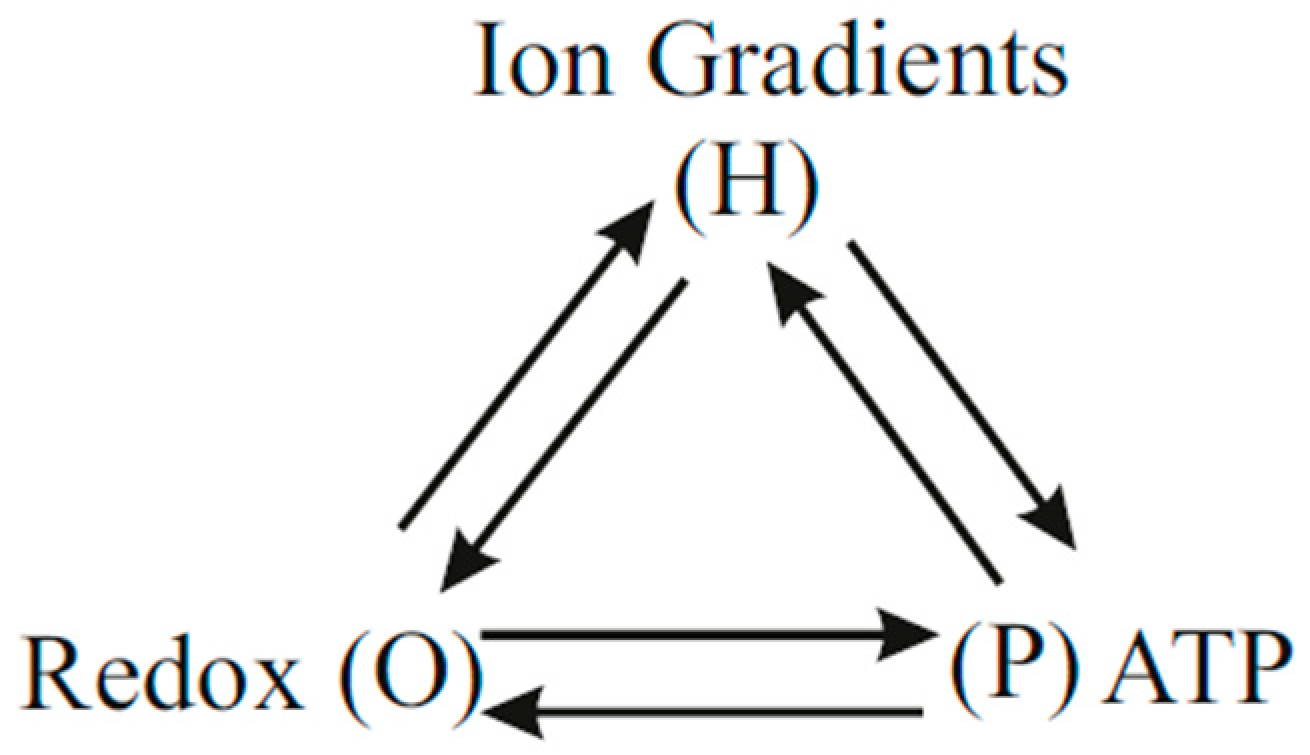
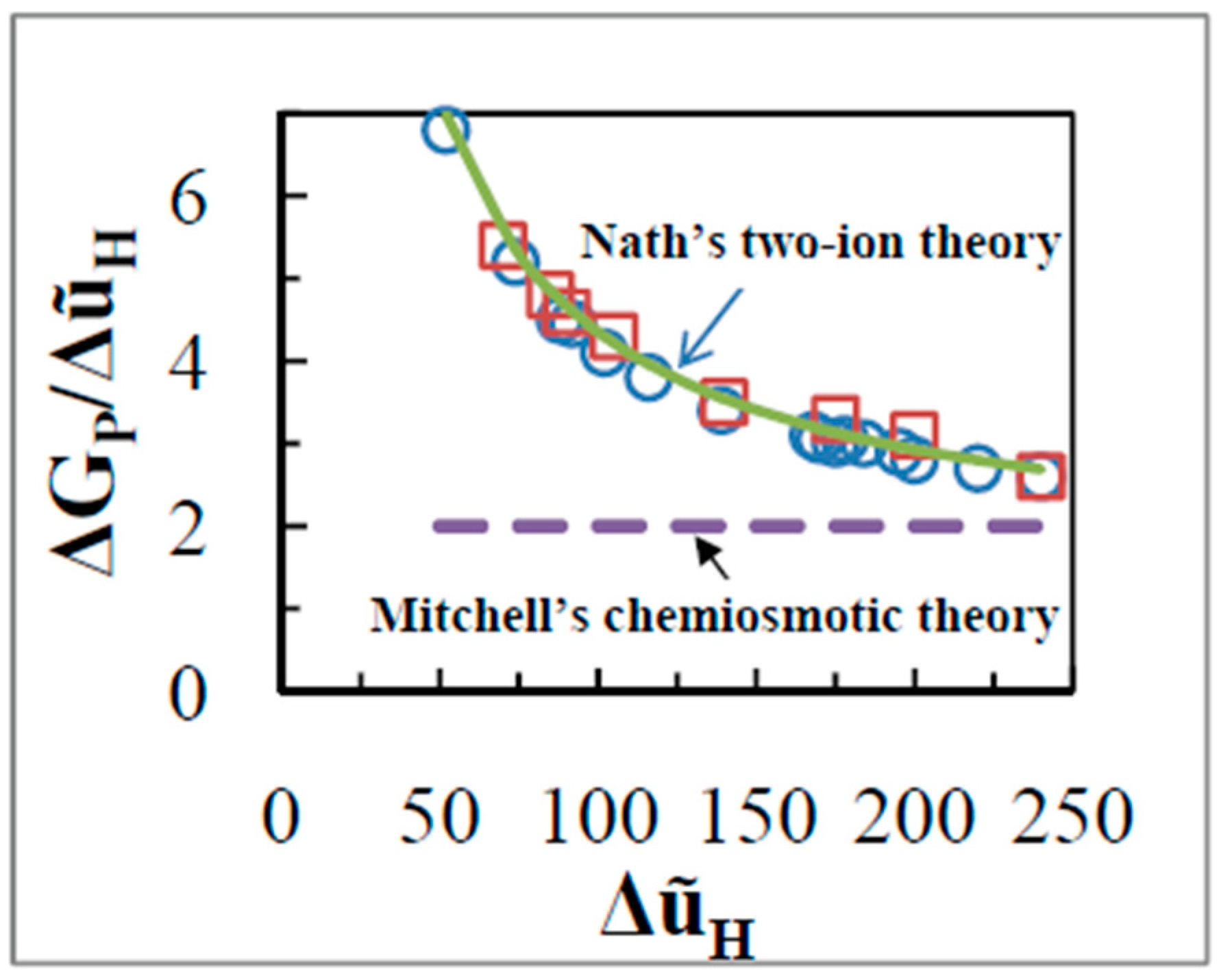
8. Molecular Mechanisms Beyond the Chemiosmotic Theory
8.1. Experiments on ADP Phosphorylation During the Transition from State 4 to State 3 or in State 3
8.2. Experiments Probing State 4
9. Experimental Methods
10. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Roels, J.A. Energetics and Kinetics in Biotechnology; Elsevier Biomedical Press: Amsterdam, The Netherlands, 1983. [Google Scholar]
- Jou, D.; Ferrer, F. A simple nonequilibrium thermodynamic description of some inhibitors of oxidative phosphorylation. J. Theor. Biol. 1985, 117, 471–488. [Google Scholar] [CrossRef]
- Kondepudi, D.; Prigogine, I. Modern Thermodynamics: From Heat Engines to Dissipative Structures; John Wiley & Sons: New York, NY, USA, 1998. [Google Scholar]
- Caplan, S.R.; Essig, A. Bioenergetics and Linear Nonequilibrium Thermodynamics: The Steady State; Harvard University Press: Cambridge, MA, USA, 1999. [Google Scholar]
- Hansen, L.D.; Criddle, R.S.; Smith, B.N.; Macfarlane, C.; Church, J.N.; Thygerson, T.; Jovanovic, T.; Booth, T. Thermodynamic law for adaptation of plants to environmental temperatures. Pure Appl. Chem. 2005, 77, 1425–1444. [Google Scholar]
- Lebon, G.; Jou, D.; Casas-Vázquez, J. Understanding Non-equilibrium Thermodynamics: Foundations, Applications, Frontiers; Springer: Berlin, Germany, 2008. [Google Scholar]
- Andresen, B. Metrics and energy landscapes in irreversible thermodynamics. Entropy 2015, 17, 6304–6317. [Google Scholar] [CrossRef]
- Dobovišek, D.; Juretić, A. Enzyme kinetics and the maximum entropy production principle. Biophys. Chem. 2011, 154, 49–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nath, S. The thermodynamic efficiency of ATP synthesis in oxidative phosphorylation. Biophys. Chem. 2016, 219, 69–74. [Google Scholar] [CrossRef]
- Nath, S. Optimality principle for the coupled chemical reactions of ATP synthesis and its molecular interpretation. Chem. Phys. Lett. 2018, 699, 212–217. [Google Scholar] [CrossRef]
- Nath, S. Coupling in ATP synthesis: Test of thermodynamic consistency and formulation in terms of the principle of least action. Chem. Phys. Lett. 2019, 723, 118–122. [Google Scholar] [CrossRef]
- Lucia, U.; Deisboeck, T.S. The importance of ion fluxes for cancer proliferation and metastasis: A thermodynamic analysis. J. Theor. Biol. 2018, 445, 1–8. [Google Scholar] [CrossRef]
- Stucki, J.W. The optimal efficiency and economic degrees of coupling of oxidative phosphorylation. Eur. J. Biochem. 1980, 109, 269–283. [Google Scholar] [CrossRef]
- Dewar, R.C.; Juretić, D.; Županović, P. The functional design of the rotary enzyme ATP synthase is consistent with maximum entropy production. Chem. Phys. Lett. 2006, 430, 177–182. [Google Scholar] [CrossRef]
- Mitchell, P. Chemiosmotic coupling in oxidative and photosynthetic phosphorylation. Biol. Rev. 1966, 41, 445–502. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P. Bioenergetic aspects of unity in biochemistry: Evolution of the concept of ligand conduction in chemical, osmotic and chemiosmotic reaction mechanisms. In Oxygen, Fuels and Living Matter, Part 1; Semenza, G., Ed.; Wiley: New York, NY, USA, 1981; pp. 30–56. [Google Scholar]
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature 1961, 191, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Nath, S. Beyond the chemiosmotic theory: Analysis of key fundamental aspects of energy coupling in oxidative phosphorylation in the light of a torsional mechanism of energy transduction and ATP synthesis − Invited review part 1. J. Bioenerg. Biomembr. 2010, 42, 293–300. [Google Scholar] [CrossRef]
- Nath, S. Beyond the chemiosmotic theory: Analysis of key fundamental aspects of energy coupling in oxidative phosphorylation in the light of a torsional mechanism of energy transduction and ATP synthesis − Invited review part 2. J. Bioenerg. Biomembr. 2010, 42, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Nath, S. Two-ion theory of energy coupling in ATP synthesis rectifies a fundamental flaw in the governing equations of the chemiosmotic theory. Biophys. Chem. 2017, 230, 45–52. [Google Scholar] [CrossRef]
- Noji, H.; Yasuda, R.; Yoshida, M.; Kinosita, K. Direct observation of the rotation of F1-ATPase. Nature 1997, 386, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Stock, D.; Leslie, A.G.W.; Walker, J.E. Molecular architecture of the rotary motor in ATP synthase. Science 1999, 286, 1700–1705. [Google Scholar] [CrossRef] [PubMed]
- Nath, S.; Rohatgi, H.; Saha, A. The torsional mechanism of energy transfer in ATP synthase. Curr. Sci. 1999, 77, 167–169. [Google Scholar]
- Nath, S.; Jain, S. BREAKTHROUGHS AND VIEWS: Kinetic modeling of ATP synthesis by ATP synthase and its mechanistic implications. Biochem. Biophys. Res. Commun. 2000, 272, 629–633. [Google Scholar] [CrossRef]
- Nath, S.; Rohatgi, H.; Saha, A. The catalytic cycle of ATP synthesis by means of a torsional mechanism. Curr. Sci. 2000, 78, 23–27. [Google Scholar]
- Nath, S. Molecular mechanisms of energy transduction in cells: Engineering applications and biological implications. Adv. Biochem. Eng. Biotechnol. 2003, 85, 125–180. [Google Scholar] [PubMed]
- Nath, S. The torsional mechanism of energy transduction and ATP synthesis as a breakthrough in our understanding of the mechanistic, kinetic and thermodynamic details. Thermochim. Acta 2004, 422, 5–17. [Google Scholar] [CrossRef]
- Nath, S. The new unified theory of ATP synthesis/hydrolysis and muscle contraction, its manifold fundamental consequences and mechanistic implications and its applications in health and disease. Int. J. Mol. Sci. 2008, 9, 1784–1840. [Google Scholar] [CrossRef] [PubMed]
- Nath, S.S.; Nath, S. Energy transfer from adenosine triphosphate: Quantitative analysis and mechanistic insights. J. Phys. Chem. B 2009, 113, 1533–1537. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Murugavel, R.; Hansen, L.D. ATP synthase and the torsional mechanism: Resolving a 50-year-old mystery. Curr. Sci. 2004, 87, 16–19. [Google Scholar]
- Iotti, S.; Sabatini, S.; Vacca, A. Chemical and biochemical thermodynamics: From ATP hydrolysis to a general reassessment. J. Phys. Chem. B 2010, 114, 1985–1993. [Google Scholar] [CrossRef] [PubMed]
- Channakeshava, C. New paradigm for ATP synthesis and consumption. J. Biosci. 2011, 36, 3–4. [Google Scholar] [CrossRef]
- Agarwal, B. A role for anions in ATP synthesis and its molecular mechanistic interpretation. J Bioenerg. Biomembr. 2011, 43, 299–310. [Google Scholar] [CrossRef]
- Villadsen, J.; Nielsen, J.; Lidén, G. Bioreaction Engineering Principles, 3rd ed.; Springer: New York, NY, USA, 2011; Chapter 4. [Google Scholar]
- Xu, L.; Liu, F. The chemo-mechanical coupled model for F1F0-motor. Progress Biophys. Mol. Biol. 2012, 108, 139–148. [Google Scholar] [CrossRef]
- Agarwal, B. Revisiting the ‘chemiosmotic theory’: Coupled transport of anion and proton for ATP synthesis. Bioenergetics 2012, 2, e116. [Google Scholar] [CrossRef]
- Rao, N.M. Medical Biochemistry, 2nd ed.; New Age International: New Delhi, India, 2014; pp. 280–281. [Google Scholar]
- Wray, V. Commentary on “Oxidative phosphorylation revisited”. Biotechnol. Bioeng. 2015, 112, 1984–1985. [Google Scholar] [CrossRef] [PubMed]
- Ivanischev, V.V. Problems of bioenergy in the light of new ideas in biology. News Tula State Univ. Natl. Sci. 2017, 1, 98–109. [Google Scholar]
- Ivanischev, V.V. Problems of the mathematical description of the chemiosmotic theory. News Tula State Univ. Natl. Sci. 2018, 3, 129–135. [Google Scholar]
- Ji, S. The Cell Language Theory: Connecting Mind and Matter; World Scientific: Singapore, 2018; Chapter 3. [Google Scholar]
- Watt, I.N.; Montgomery, M.G.; Runswick, M.J.; Leslie, A.G.W.; Walker, J.E. Bioenergetic cost of making an adenosine triphosphate molecule in animal mitochondria. Proc. Natl. Acad. Sci. USA 2010, 107, 16823–16827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Bueler, S.A.; Rubinstein, J.L. Atomic model for the dimeric FO region of mitochondrial ATP synthase. Science 2017, 358, 936–940. [Google Scholar] [CrossRef] [PubMed]
- Nath, S. Integration of demand and energy sides in the ATP energy economics of cells. Biophys. Chem. 2019, 253, 1–8. [Google Scholar] [CrossRef]
- Nath, S. Analysis of molecular mechanisms of ATP synthesis from the standpoint of the principle of electrical neutrality. Biophys. Chem. 2017, 224, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Kalckar, H. Phosphorylation in kidney tissue. Enzymologia 1937, 2, 47–52. [Google Scholar]
- Ochoa, S. Efficiency of aerobic phosphorylation in cell-free heart extracts. J. Biol. Chem. 1943, 151, 493–505. [Google Scholar]
- Chance, B.; Williams, G.R. Respiratory enzymes in oxidative phosphorylation: I. Kinetics of oxygen utilization. J. Biol. Chem. 1955, 217, 383–394. [Google Scholar]
- Chance, B.; Williams, G.R. The respiratory chain and oxidative phosphorylation. Adv. Enzymol. 1956, 17, 65–134. [Google Scholar]
- Ferguson, S.J. The ups and downs of P/O ratios (and the question of non-integral coupling stoichiometries for oxidative phosphorylation and related processes). Trends Biochem. Sci. 1986, 11, 351–353. [Google Scholar] [CrossRef]
- Hinkle, P.C.; Kumar, M.A.; Resetar, A.; Harris, D.L. Mechanistic stoichiometry of mitochondrial oxidative phosphorylation. Biochemistry 1991, 30, 3576–3582. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, D.G. The influence of respiration and ATP hydrolysis on the proton-electrochemical gradient across the inner membrane of rat-liver mitochondria as determined by ion distribution. Eur. J. Biochem. 1974, 50, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.P.; Gu, Q.; Xiong, Y.; Mitchell, R.A.; Ernster, L. P/O ratios reassessed: Mitochondrial P/O ratios consistently exceed 1.5 with succinate and 2.5 with NAD-linked substrates. FASEB J 1996, 10, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Kedem, O.; Caplan, S.R. Degree of coupling and its relation to efficiency of energy conversion. Trans. Faraday Soc. 1965, 21, 1897–1911. [Google Scholar] [CrossRef]
- Juretić, D.; Westerhoff, H.V. Variation of efficiency with free-energy dissipation in models of biological energy transduction. Biophys. Chem. 1987, 28, 21–34. [Google Scholar] [CrossRef]
- Sciubba, E. What did Lotka really say? A critical reassessment of the “maximum power principle”. Ecol. Modell. 2011, 222, 1347–1353. [Google Scholar] [CrossRef]
- Nath, S. A thermodynamic principle for the coupled bioenergetic processes of ATP synthesis. Pure Appl. Chem. 1998, 70, 639–644. [Google Scholar] [CrossRef] [Green Version]
- Lemasters, J.J. The ATP-to-oxygen stoichiometries of oxidative phosphorylation by rat liver mitochondria. J. Biol. Chem. 1984, 259, 13123–13130. [Google Scholar]
- Westerhoff, H.A.; van Dam, K. Thermodynamics and Control of Biological Free-Energy Transduction; Elsevier: Amsterdam, The Netherlands, 1987. [Google Scholar]
- Martyushev, L.M. The maximum entropy production principle: Two basic questions. Phil. Trans. R Soc. B 2010, 365, 1333–1334. [Google Scholar] [CrossRef] [PubMed]
- Martyushev, L.M. Entropy and entropy production: Old misconceptions and new breakthroughs. Entropy 2013, 15, 1152–1170. [Google Scholar] [CrossRef]
- Azzone, G.F.; Pozzan, T.; Massari, S. Proton electrochemical gradient and phosphate potential in mitochondria. Biochim. Biophys. Acta 1978, 501, 307–316. [Google Scholar] [CrossRef]
- Allegretti, M.; Klusch, N.; Mills, D.J.; Vonck, J.; Kühlbrandt, W.; Davies, K.M. Horizontal membrane-intrinsic α-helices in the stator a-subunit of an F-type ATP synthase. Nature 2015, 521, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Schneider, W.C.; Hogeboom, G.H. Intracellular distribution of enzymes: Further studies on the distribution of cytochrome c in rat liver homogenates. J. Biol. Chem. 1950, 183, 123–128. [Google Scholar]
- Myers, D.K.; Slater, E.C. The enzymic hydrolysis of adenosine triphosphate by liver mitochondria. I. Activities at different pH values. Biochem. J. 1957, 67, 558–572. [Google Scholar] [CrossRef] [PubMed]
- Fiske, C.H.; Subbarow, Y. The colorimetric determination of phosphorus. J. Biol. Chem. 1925, 66, 375–400. [Google Scholar]
- Mitchell, P.; Moyle, J. Estimation of membrane potential and pH difference across the cristae membrane of rat liver mitochondria. Eur. J. Biochem. 1969, 7, 471–484. [Google Scholar] [CrossRef]
- Bergmeyer, H.U. Methods of Enzymatic Analysis; Elsevier: New York, NY, USA, 1974. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Optimization | Maximum Output Flow, JP | Maximum Output Power, JPAP | Maximum Economic Output Flow, JPη | Maximum Economic Output Power, JPAPη |
|---|---|---|---|---|
| Single variable with q = 1 | <0 | 50 | 50 | 67 |
| Two variable [13,59] | 23.6 | 41.4 | 53.5 | 61.8 |
| Output Function | Normalized (As in Stucki [13]) | Leak | Redox Slip | ATPase Slip |
|---|---|---|---|---|
| (Without Normalization [9,10,11,57,59]) | ||||
| JP | 0.786 | 0.612 | 0.100 | 0.289 |
| JPAP | 0.909 | 0.882 | 0.864 | 0.886 |
| JPη | 0.953 | 0.946 | 0.941 | 0.944 |
| JPAPη | 0.973 | 0.969 | 0.969 | 0.969 |
| 2,4-dinitrophenol (μM) | JO(natom O/(mg min)) | JP(nmol ATP/(mg min)) | ΔũH (mV) |
|---|---|---|---|
| 0 | 98 | 152 | 233 ± 5 |
| 3 | 97 | 105 | 200 ± 5 |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nath, S. Entropy Production and Its Application to the Coupled Nonequilibrium Processes of ATP Synthesis. Entropy 2019, 21, 746. https://doi.org/10.3390/e21080746
Nath S. Entropy Production and Its Application to the Coupled Nonequilibrium Processes of ATP Synthesis. Entropy. 2019; 21(8):746. https://doi.org/10.3390/e21080746
Chicago/Turabian StyleNath, Sunil. 2019. "Entropy Production and Its Application to the Coupled Nonequilibrium Processes of ATP Synthesis" Entropy 21, no. 8: 746. https://doi.org/10.3390/e21080746
APA StyleNath, S. (2019). Entropy Production and Its Application to the Coupled Nonequilibrium Processes of ATP Synthesis. Entropy, 21(8), 746. https://doi.org/10.3390/e21080746