1. Introduction
Some classical works by Boltzmann, Gibbs and Maxwell have defined entropy under a statistical framework. A useful entropy concept is the Shannon entropy since it is a basic tool to quantify the amount of uncertainty in many kinds of physical or biological processes [
1,
2,
3,
4,
5,
6]. It may be interpreted as a quantification of information loss [
1,
2,
3,
7,
8,
9]. On the other hand, entropy-based tools have been also proposed to evaluate the propagation of epidemics and related public control interventions (see, for instance, [
10,
11,
12,
13,
14,
15,
16,
17] and some of the references therein). There are also models whose basic framework relies on the use of entropy tools, as for instance [
13,
14,
15,
16]. It can be also pointed out that the control designs might be incorporated to some epidemic propagation and other biological problems, see, for instance, [
18,
19,
20,
21,
22,
23,
24,
25,
26,
27], and, in particular, for the synthesis of decentralized control in patchy (or network node-based) interlaced environments [
24,
27]. A typical situation is that of several towns each with its own health center, whose susceptible and infectious populations, apart from their coupled self-dynamics among their integrating subpopulations, might also mutually interact with the subpopulations of the neighboring nodes through in-coming and out-coming fluxes.
It can be pointed out that the knowledge or estimation of the transient behavior of the infection is very relevant for the hospital management of the disease since it is necessary to manage the availability of beds and other sanitary utensils and sanitary means, in general. The work by Wang et al. in [
11] pays mainly attention to the description of the transient behavior of the evolution of epidemics rather than to the equilibrium states. The main purpose in that paper was to formulate the time interval occurring between the time instant of the maximum of the infection, which gives a relative maximum of the infection evolution through time (and which zeroes the first time-derivative of the infection function), and the time instant giving its previous inflection time instant. It turns out that the knowledge of the first part of the transient evolution is very relevant to fight against the initial exploding of the illness since any eventual control intervention is typically much more efficient as far as it is taken as quickly as possible. The model proposed in [
11] is a time-varying differential equation of first-order describing the infectious population which is the unique explicit one in the model. It is also pointed out in that paper that the time-varying coefficient might potentially contain the supplementary environment information to make such an equation well-posed to practically describe a concrete disease evolution. An interesting point of that work is that the infection evolution is identified with a log-normal distribution whose parameterization is selected in such a way that the entropy production rate is maximized. The above proposed theoretical first-order model has been proved to be very efficient to describe the data of SARS 2003. Alternative interpretations of the entropy in terms of maximum entropy or maximum entropy rate are given, for instance, in [
12,
13,
14] and some references therein.
This paper studies how to link the extension of the first-order differential system proposed in [
11] for the study of infection propagations to epidemic models with more integrated coupled subpopulations (such as susceptible, immune, vaccinated etc.) by introducing the coupling and control information through the time-varying coefficient which drives the basic differential equation model. It is considered relevant the control of the infection along its transient to fight more efficiently against a potential initial exploding transmission. Note that the disease-free and endemic equilibrium points and their stability properties depend on the concrete parameterization while they admit a certain design monitoring by the choice of the control and treatment gains and the use of feedback information in the corresponding controls. See, for instance [
19,
27]. Therefore, special attention is paid to the transients of the infection curve evolution in terms of the time instants of its first relative maximum towards its previous inflection time instant since there is a certain gap in the background literature concerning the study of such transients. The ratio of such time instants is later on considered subject to some worst-case uncertainty relations via the calculation and analysis of an “ad hoc” Shannon’s entropy. Note that entropy issues have been considered in the study of biological, evolution and epidemic models by incorporating techniques of information theory. See, for instance [
11,
12,
13,
28,
29,
30,
31,
32]. It is well-known that the entropy production theorems might be classified according to a generalized sequence of stable thermodynamic states. Also, the thermodynamic equilibrium, which is characterized by the absence of gradients of state or kinematic variables, is in a state of maximum entropy and zero entropy production [
33,
34]. Furthermore, linear non-equilibrium processes are associated with entropy production so that the entropy concept may be also invoked in transient processes [
35]. On the other hand, it may be pointed out that uncertainties can appear in the characterization of the infection evolution through time, even in deterministic models, due to parameterization uncertainties, fluxes of populations or existing uncertainties in the initial conditions. Other mathematical techniques of interest which combine analytical and numerical issues have been also been applied to the analysis and discussion of epidemic models with eventual support of mathematical techniques on homotopy analysis and distribution functions as, forinstance, the log-normal distribution [
36,
37]. For instance, in [
38], the SIR and SIS epidemic models are solved through the homotopy analysis method. A one-parameter family of series solutions is obtained which gives a method to ensure convergent series solutions for those kinds of models. On the other hand, in [
39], the analytic solutions of an SIR epidemic model are investigated in parametric form. It is also found that the generalization of a SIR model including births and mortality with vital dynamics might be reduced to an Abel-type which greatly simplify the analysis.
The paper is organized as follows:
Section 2 gives an extension of the basic model of [
11] to be then compared in subsequent sections with some existing models with several subpopulations. Such a model only considers the infection evolution through time and it is based on the action of two auxiliary non-negative functions which define appropriately the time-varying coefficient which defines the first-order differential equation of the infection evolution. The model includes, as particular case, that of the abovementioned reference where both such auxiliary functions are identical to the time argument. Particular choices of those functions make it possible to consider alternative effects linked to the basic model like, for instance, the influence on the infectious subpopulation of other coupled subpopulations in more general models like, for instance, the susceptible, exposed, recovered or vaccinated ones. It is also possible to include the control effects through such a varying coefficient, if any, like for instance, the vaccination and treatment controls. Some basic formal results are stated and proved mainly concerning with the first relative maxima and inflection time instants of the infection curve through time. The above two time instants are relevant to take appropriate control interventions to fight against an initially exploding infectious disease.
Section 3 links the basic model of
Section 2 with some known epidemic models which integrate more subpopulations than just the infectious one, like for instance, the susceptible and recovered subpopulations, The time-varying coefficient driving the infection evolution is defined explicitly for each of the discussed epidemic models. Basically, it is taken in mind that some relevant information of higher-order differential epidemic models concerning the transient trajectory solution can be captured by a parameter-dependent and time-varying coefficient which drives a first-order differential equation to the light of the basic model of
Section 2. So, the time-varying coefficient describing the infection evolution depends in those cases of the remaining subpopulations integrated in the model. The maximum and inflection time instants are characterized for some given examples involving epidemic models of several subpopulations. In particular, the last one of the discussed theoretical examples includes the effects of vaccination and treatment intervention controls generated by linear feedback of the susceptible and infectious subpopulations, respectively. Later on,
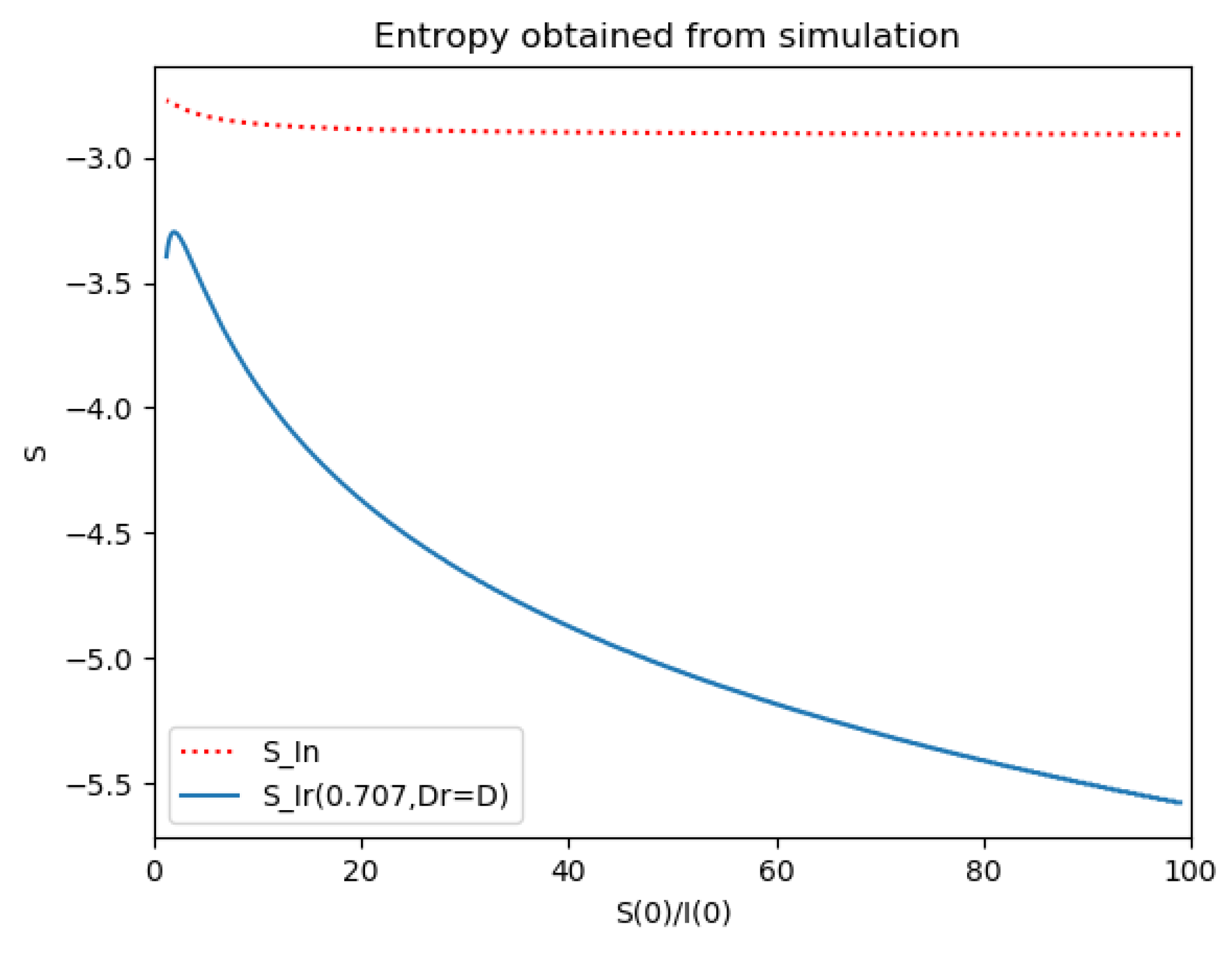
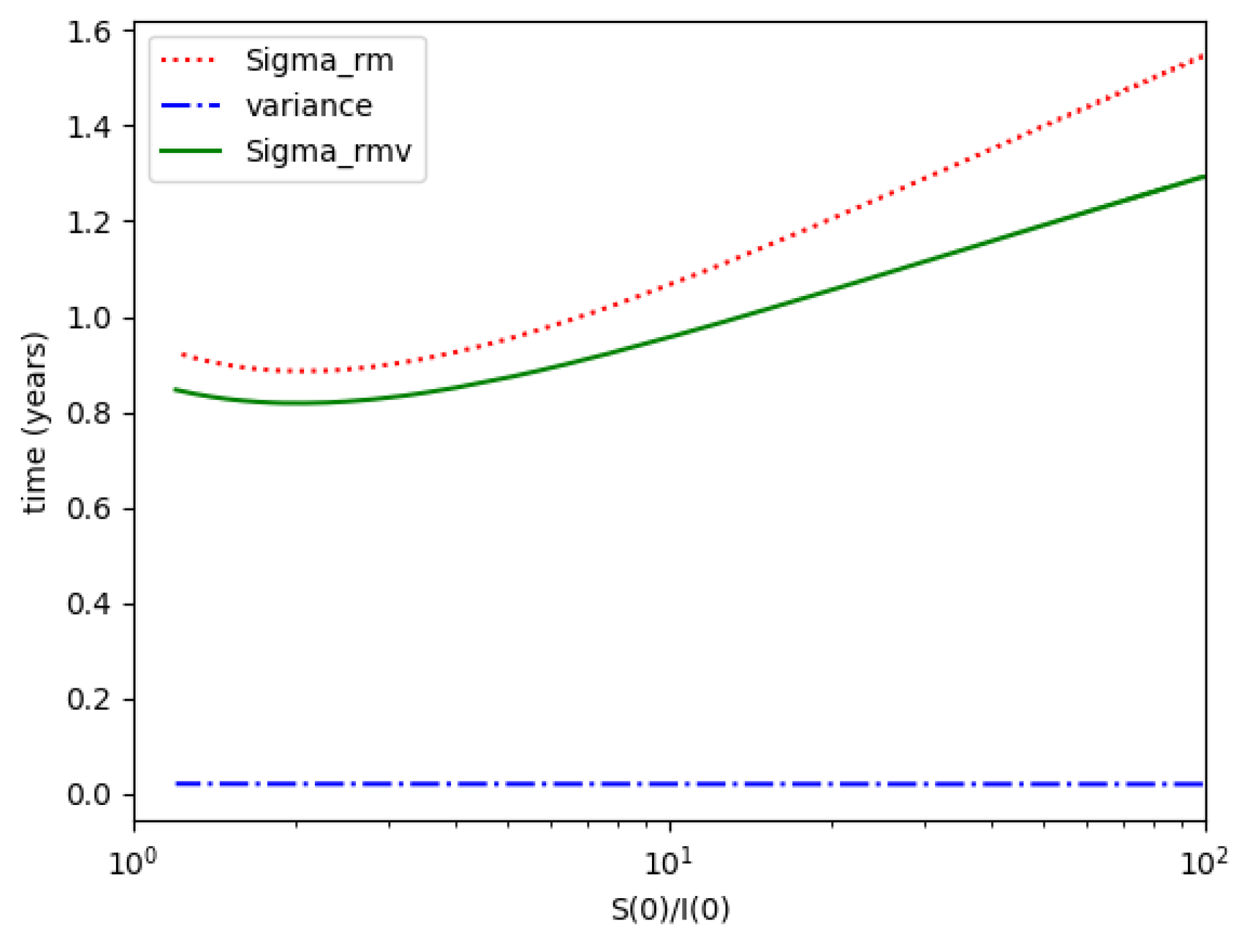
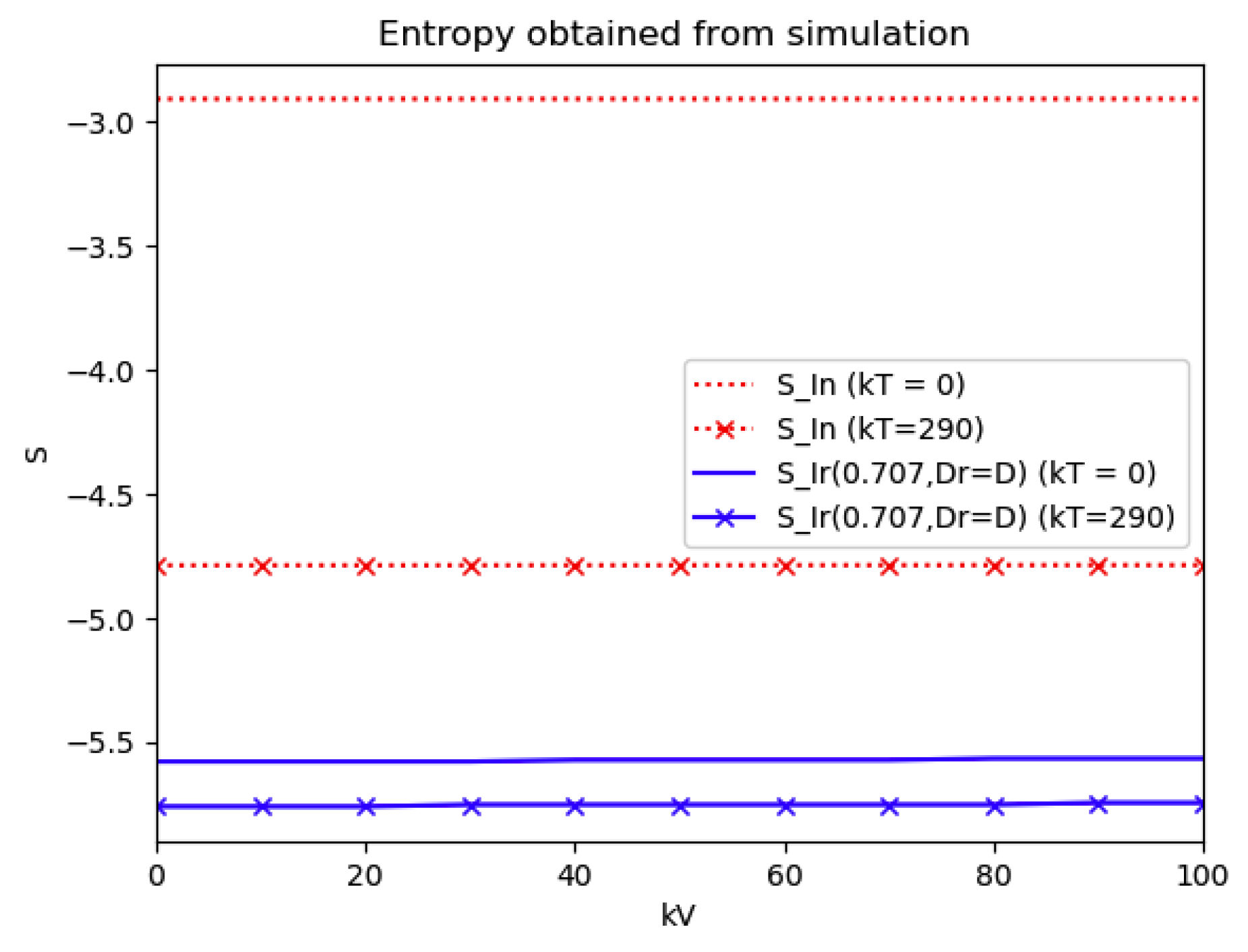
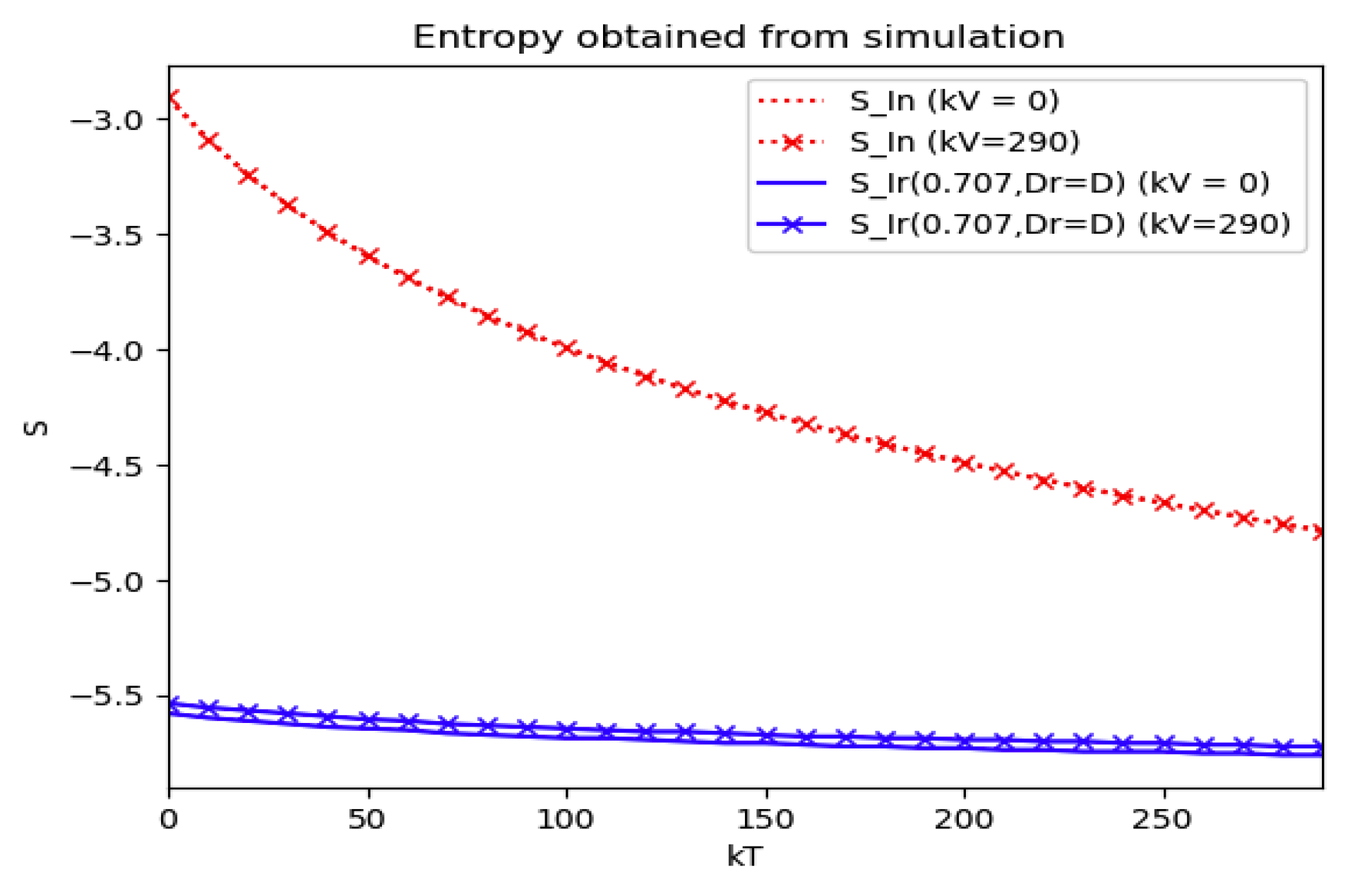
Section 4 investigates the entropy associated with the infection accordingly to the generalizations of
Section 2 concerning the specific structure of the time-varying coefficient describing the infection dynamics and its links with the theoretical examples discussed in
Section 3. The error of the entropy related to the reference one associated with the log-normal distribution is estimated. In practice, that property can be interpreted in terms of public medical and social interventions which control the disease propagation when introducing the controls of the last example discussed in
Section 3. The second part of
Section 4 is devoted to linking the entropy and inflection and maximum infection time instants and their reached values of the discussed multi-population structures to their counterparts of the maximum dissipation rate being associated to the formulation of a simpler model based on the log-normal distribution and one-dimensional infection dynamics. Some numerical tests are performed for comparisons of the entropies and its width of the basic model with two of the discussed examples in the previous sections which involve the presence of more than one integrated subpopulations. Finally, conclusions end the paper.
2. The Basic Model Description and Some Related Technical Results
Since disease propagation can be interpreted as a thermodynamic system, it can be assumed that the rate of increase or decrease is proportional to the infection at the previous day following the approach of modelling the rate of chemical reactions, [
11]. Thus, assume that the infection evolution obeys the following time-varying differential equation:
where
is continuous and time differentiable on
. The particular structure of the varying coefficient
depends on the balances between the spreading mechanism and the exerted controls during the public intervention. Such a coefficient contains the available information related to the incorporation of all the control mechanisms and the coupling dynamics between the infectious populations and the remaining interacting ones such as the susceptible, immune or vaccinated ones. By taking time-derivatives with respect to time in (1), one gets:
It is proposed in [
11] to consider two relevant time instants in the disease evolution, namely:
- (1)
The inflection time instant of I(t) which is the date in the infection evolution at which the controlling actions take effect on the evolution. Typically, this time instant is the undulation point date in the evolution of , that is the zero of , provided that the first non-zero derivative of , occurs for some even since this last condition ensures that the undulation time instant is the inflection time instant.
- (2)
The critical time instant at which the spread rate turns from initial growing to decrease which can be empirically attributed to the global influence of the control interventions. This time instant is a relative maximum of I(t) and it satisfies the constraints and under the reasonable assumption that .
It turns out that, along the whole disease evolution, several successive inflection points and relative maxima can happen. The subsequent result which is concerned with the non-negativity, boundedness and asymptotic vanishing property of the infection as time tends to infinity and its two first- time derivatives is immediate from the above expressions (1) and (2):
Theorem 1. The following properties hold:
- (i)
The infection population and its two first-time derivatives obey the following time evolution equations: - (ii)
;if and only if; and;if and only if.
- (iii)
Ifthen;for someif and only ifis such that;.
- (iv)
asfor any given finiteif and only if.
- (v)
Ifand;for somethen;if and only if, for some,;. Ifthen;if and only if;, for someprovided that.
- (vi)
asfor any given finiteif and only if.
Ifis bounded andasthenas.
- (vii)
Ifthen;if and only if;, for some.asfor any given finiteif and only if.
Ifis bounded andasthenas.
Note that
(respectively,
) is infinity at
while it is bounded for
, as it happens for instance with the
—function proposed in [
11], then
(respectively,
) is still bounded under the conditions of Theorem 1 (v) (respectively, Theorem 1 (vii)) on
.
Note also that the vanishing infection condition of Theorem 1 typically occurs under convergence of the solution to the disease-free equilibrium point if the disease reproduction number is less than one [
19,
22,
23,
24,
27,
29,
30,
36]. However, it can happen that the infection oscillates around some stable equilibrium or that it converges to a nonzero positive constant defining the corresponding component of the endemic equilibrium steady-state as it is discussed in the next result.
Corollary 1. The following properties hold:
- (i)
Assume that there exists somesuch thatasand thatas. Then,,andas.
- (ii)
Assume thatasand thatis uniformly continuous. Then,,andas. Assume, in addition, thatis uniformly continuous. Thenas.
Proof of Property (i). Follows directly from (1)–(3). On the other hand, since is uniformly continuous and the limit exists and it is finite then as (Barbalat´s Lemma) and as from (3), is bounded, since being continuous, it cannot diverge in finite time, and as from (1). If, furthermore, is uniformly continuous and, since then as (again from Barbalat´s Lemma). Since as then as from (2). □
Let us introduce the following definitions and lemma of usefulness for the proof of the subsequent theorem [
36]:
Definition 1. Letbe everywhere continuous and twice differentiable at. Then,is an undulation point (or pre-inflection point) ofif.
Inflection points of the continuous and twice-differentiableare the undulation points of the function where the curvature changes its sign, that is, points of change of local convexity to local concavity or vice-versa. They are also the isolated extrema of. A well-known technical definition and a related result on inflection points follow:
Definition 2. Letbe everywhere continuous and twice differentiable atwhich is an isolated extremum of(that is, a local maximum or minimum, and also an undulation point of,as a result).
Lemma 1. The following properties hold:
- (i)
Letbe everywhere continuous and twice differentiable at. Then,is an inflection point ofiffor some sufficiently small.
- (ii)
Letbe everywhere continuous and an odd number-times differentiable, within a neighborhood ofwhich is an undulation point ofsatisfyingforand. Then,is an inflection point of.
The subsequent result has a very technical proof leading to the basic result that the zeros at finite time instants of and alternate if I(t) is sufficiently smooth and is sufficiently smooth. In order to simplify the result proof, it is assumed, with no loss in generality, that the disease dynamics (1)–(2) has no equilibrium points such that the zeros under study are isolated.
Theorem 2. Assume that the functiondefined by, whereandare everywhere continuous and time-differentiable such thatwithfor some, and furthermore,fulfills the constraints:for any given positive real number, withand, whereandare assumed to be nonempty and of zero Lebesgue measure. Then, the following properties hold:
- (i)
, equivalently,.
- (ii)
(a)with, and (a) if(withdenoting the infinite cardinality of denumerable sets) then;for any pairsandfulfillingand,
(b) ifthenfor.is subject to the constraint,;and.
- (iii)
is subject to;,andfor any.
Proof. First, note that ; , since even if . On the other hand, is the set of undulation points of and it is clear that is contained in the set of relative maximum and minimum points of . The properties (i)–(iii) are now proved:
Proof of Property (i). It is now proved that
is the set of extreme points of
which is disjoint to its set of undulation points
. Assume, on the contrary, that there is some
such that
. Then,
since
, and then the disease-free equilibrium point is reached in finite time contradicting the fact that
is only zero at finite time for a discrete set of time instants satisfying
so that
if and only if
. Then,
is a disease-free equilibrium point which is reached in finite time which contradicts the given hypothesis. So, it is easy to see that
and
are discrete sets of non-negative real time instants which can be strictly ordered. Note also from (1)–(2) that:
If
then
since
. Also,
and, if
and since
, one has:
Now, if there is some , equivalently, , then from (9) since and and, furthermore, one gets from (8) that since . But one also has that , since ; from the first identity of (8). Then, is a contradiction so that . Equivalently, . Property (i) has been proved. □
Proof of Property (ii). Since
then
so that:
Since the zeros of and those of its first time- derivative do not coincide since (from Property (i)), it turns out that the two sets of respective zeros alternate if there are not two zeros of within any open time interval of two consecutive zeros of or vice-versa. One proceeds by contradiction arguments by assuming two cases which are both rebutted.
Case 1: Assume that there are two consecutive zeros of between two consecutive zeros of , then satisfying the constraint for some two consecutive time instants in and two consecutive time instants in so that . Assume that for some then so that and then are not consecutive time instants in and this case has to be excluded from further reasoning. Now, assume that for all and , otherwise, if then and are not consecutive time instants in . Thus, for all . Since for all , it has no sign change in so that and since is continuous then which contradicts that . It has been proved that Case 1 is impossible cannot happen.
Case 2: Assume now that there are two consecutive zeros of between two consecutive zeros of , that is for some consecutive time instants in and some two consecutive time instants in . Then, for all since, otherwise, there exists some such that , and then the previously claimed constraint does not hold, and also for all since, otherwise, there exists some such that and then and are not two consecutive time instants in as claimed. Also, note that.
with and since . But then, by continuity arguments on , there is a change of sign point which zeroes this function which contradicts for all . Then, Case 2 is impossible so that cannot happen and Property (ii) has been proved. □
Proof of Property (iii). Assume that, contrarily to the statement, . If then and the equilibrium point is reached in finite time what is impossible, since , for a non-trivial solution of a continuous-time first-order differential equation with continuous-time parameterization. Then, is impossible. Now, assume that and with and then it exists some such that and . As a result, there is and then there are two consecutive undulation time instants what contradicts Property (ii). As a result, as claimed. □
Remark 1. In Theorem 2, note that the setsandhave the following properties:
They are nonempty so that there is at least onesuch thatimplying thatand at least onesuch thatimplying that. Otherwise, the infection could converge asymptotically to zero as time goes to infinity but it would not have finite zeros,
They are sets of zero Lebesgue measure so that they are denumerable discrete sets of strictly ordered isolated real points, for any real numbers,
They fulfill thatwithso that they are of either identical finite or infinite cardinal or the cardinal ofis finite and exceeds that ofby one,
Ifthen, that is, if both sets have infinity cardinal or identical finite one then any ordered points ofandalternate.
On the other hand, note that:
Equation (4) establishes that is the set of zeros of . At those zeros, the first-time derivative of the infection function is zeroed from (1) without such a function being necessarily zero while on the other hand, Equation (5) is a nonzero real constant for any finite undulation time instant of zeroing the second derivative of the infection function according to (2) which holds if from (5). The fact that (5) is constant follows easily under periodicity conditions of the same or integer multiple/submultiple periods of and .
Since has no finite zero coincident with a zero of its first time-derivative, by hypothesis, then since from inspection of (8)–(9). This is equivalent to , that is, the finite zeros which make zero and which do not make zero do not make zero either . However, if from (2), provided that is twice everywhere continuously differentiable in but this can only happen as time tends to infinity for certain structures of and . Note that the constraint (5) also implies that the auxiliary functions used to define the function in (1) fulfill the constraint ; .
By examining Definitions 1 and 3 and Lemma 1, it turns out that the set of undulation points of includes but, maybe non-properly, the set of its inflection points. However, it suffices to give some further weak conditions on , that is, on to guarantee that every undulation point of is also an inflection point. Some such conditions are discussed in the next corollary.
Corollary 2. The following properties hold:
- (i)
Then, the setof undulation points ofis the set of its inflection points.
- (ii)
Assume thatare twice continuously differentiable at each undulation point. Then, the sets of undulation points and that of the inflection points ofcoincide if
Proof. Note that , so that . Since , if and , since is continuous, one gets that if and only if . Property (i) has been proved.
On the other hand, if
are twice continuously differentiable at each undulation point
of
, then
exist in
. Then, defining
;
yields:
Since ; then ; if and only if ; , equivalently, if and only if ; which is fully equivalent to the condition of Property (ii). The proof is complete. □
Remark 2. Note that Theorem 2 applies, in particular, to the case when there are equilibrium points with the initial conditions being distinct from such points. It can be also extended by including the above case by redefining finite discrete sets of the zeros ofand¸for any givenin the sense that the eventual zeros at finite time ofandalternate although an equilibrium points has not still been reached provided that it exists.
Inspired in Theorem 2, some conditions are discussed in the next result which imply that the first undulation point of the infection evolution function (i.e., the first zero of its second-time derivative) precedes the first zero of its first time-derivative. It is not required that the infection has necessarily a disease-free equilibrium point or that it might be oscillatory leading to successive zeros of its time- derivative along time.
Theorem 3. Assume that the function, whereandare everywhere continuous and time-differentiable and satisfy the constraints:
- (1)
;,
- (2)
- (3)
andif
- (4)
Assume also that. Then,;;and there is somesuch that;and.
Proof. Note from the definition of
, (1), (2) and the given constraints 1 and 2 that
, since
,
, since
),
, from the condition 2 since
and since
is continuous and time-differentiable since
are everywhere continuous and time-differentiable. Note also that, from the given assumptions and constraints,
since
by hypothesis,
and
. Furthermore,
. From the constraint 3 and the continuity of
, one has that
are continuous and bounded on
,
;
and
;
and some
. Furthermore since
and
, from the constraint 4,
;
, from the constraint 1, and
and
if
, from the constraint 3. Then
;
. Since
are continuous and positive on any bounded interval
then
is positive and finite on
. It is now proved that
is the first zero of
. Assume that this is not the case so that there is some
such that
, with
, and
;
. Then
from (2) and the infection extinguishes in a finite time
. This leads to a contradiction since
since
and
;
. Therefore, if
such that
then
. But then
from (1) which contradicts that
;
. As a result,
is the first zero of
and there is no
such that
. Since
are continuous with
;
and
and
;
and some
then there is some
such that
. Assume that this is not the case. Then,
. Hence, a contradiction arises. Thus, there is some
such that
. □
Remark 3. Note that, under all the conditions of Theorem 3,;and. Furthermore, the first zero ofoccurs at, there is nosuch thatand there is somesuch that.
The following example describes the basic model proposed in [
11] under a first-order differential equation for the infection evolution without any entropy considerations at this stage:
Example 1. The function, for some, proposed in [11] satisfies all the conditions of Theorem 3 withand. It satisfies, in addition, that. This function satisfies also the given further conditions of Theorem 2with. Note that the conditionof Theorem 3 avoids thatifso thatis a zero of.
It can be argued that the proposed basic model (1) is a very simple time-varying differential equation of first-order which describes the infective population time-evolution. Note that the use of appropriate particular structures in the definition of the time-varying coefficient can take care of the eventual incorporation of the necessary supplementary environment information to make such an equation well-posed to practically describe a concrete disease evolution through time. The incorporation which can be incorporated is the eventual couplings of the infectious subpopulation with another ones (such as the susceptible, recovered or vaccinated subpopulations and their associated dynamics) or the information about the feedback information controls in more elaborated models. The next section develops some work in this direction.
3. Further Examples of Linking the Basic Model to Some Existing Epidemic Models Incorporating Other Subpopulations
The infection description via (1) assumes implicitly that it has a first-order dynamics. It has been argued that in (1) contains the information about the controls and other coupled subpopulations influencing the disease evolution through time. It can be of interest to discuss its application to infection descriptions described by differential equations of orders higher than one which is a very common situation in disease transmission mathematical models.
It is now seen how a well-known epidemic model can be also discussed under the point of view of Theorem 3. In the subsequent example, the above characterization, based on the first zero of infection evolution time-derivative and on the undulation point of the infection evolution, is used for a model with three subpopulations via an appropriate choice of and in the definition of .
Example 2. Consider the following SIR model without demography [30]:where,andare, respectively, the susceptible, infectious and recovered (or immune) subpopulations, under nonzero initial conditions being subject to, whereis the coefficient transmission rate andis the removal or recovery rate (its inversebeing the average infectious period). The mathematical study of this model and their variants is not easy as seen in [30,40]. First, note that the total population;is constant for all time. The basic reproductive ratio (or reproduction number) isand, if, thenwhile if, it becomes endemic for all time since. The solution of (10) becomes in closed form: Note that by combining the above equations that:
Note from (11) that is non-increasing so that there exists a susceptible equilibrium subpopulation for any given non-negative initial conditions. Note also from (10) that and then ; Note that If then , and ; . We examine three cases for :
Case (a) if then and ; , then and as . Since is non-increasing, . This implies that and , at exponential rate as for some from (10) and (11) since so that . Then, is integrable on . Thus, so that (then there is a nonzero susceptible equilibrium level) and .
Case (b) if then as since is non-increasing and then it converges to satisfying . By inspection of the second equation of (11), it also follows that and as satisfying and . Assume that then from the first equation of (11). But if then since then is strictly decreasing on for some finite from the second equation of (11). Hence, a contradiction to follows implying that if . Now, assume that . Then, from the second equation of (11), as . But then , from the first equation of (12), since if and then . From the second equation of (12) and, under a similar reasoning as that of Case a, is integrable on and . In summary, if and then and as in the same way as in Case a if .
Case (c) if then from (10) and is increasing on some interval . The fact that is strictly increasing on some initial time interval is of interest from the point of view of hospital management of availability of beds and other sanitary specific means in the event that the disease might have a relevant number of seriously infected individuals. Since is non-increasing then either , and as or as from (11) since is non-increasing. The firs possibility is unfeasible since from the first equation of (11) as . Then, as . Now, first, assume that . Then, from the first equation of (12), as . Then, which contradicts that , As a result, . Now, assume that . Then, from (11), and being square-integrable, and following a similar argument as that of Cases a–b, one again concludes that so that and , as a result. But, since then from (11) since is strictly decreasing after some finite time instant and integrable on and a following again the reasoning of Cases a–b, one concludes that . As a result, if and , then , and . Thus, the relevant conclusions on the disease- free equilibrium point which is a disease- free one are similar for the three above cases.
On the other hand, since
it exists a finite
such that
and
,
, if
and, furthermore,
and also:
and
under the reasonable assumption that
is sufficiently small (the initial numbers of infectious is usually very small in practice) satisfying
. As a result, there is some time instant
such that
so that it is an undulation point of
. As a result, we find that if the basic reproduction number exceeds unity then the infection curve corresponding to the endemic solution has a minimum at a larger time instant that the one defining its undulation point. That situation corresponds to the situation of small initial infection force with reproduction number greater than one. On the other hand, if
, then
does not hold.
Comparing the infectious subpopulation evolution to (1) and the structure of the function in Theorem 3 yields:
. If one defines
;
and
;
, then
;
. It is easy to verify that these functions satisfy the conditions of Theorem 3.
In the case when the reproduction number is less than unity and it is an upper-bound of the normalized susceptible population, each primary infection generates, in average, less than one secondary one so that the infection extinguishes asymptotically. According to this particular model, also the susceptible subpopulation extinguishes asymptotically. See Case a referred to (11). Thus, the disease-free equilibrium point is . In this case, as but there are no finite time instants of minimum and undulation of the infectious curve to the light of Theorem 3.
However, we can have a practical visualization of the disease removal by defining a design quadruple
and the following cut associate time instants:
Note that and generalize the roles of the time instants and , that is, the finite minimum infection and undulation time instants, respectively, within prescribed margins when those time instants do not exist.
Example 3. Consider Case a of Example 2 so thatleading toandasand,andare strictly decreasing on. Take prescribed constantsfor. The solution trajectory converges to the disease-free equilibrium point at exponential rate. Then, one gets by combining (10)–(12) and (18) that:implying that:which leads to:
and:
what implies that ; such that: Example 4. Consider the following SIS model with vaccination and antiviral or antibiotic controls:subject to,withwhere the vaccination and treatment feedback controls on the susceptible and infectious are, respectively,andwith. If it is assumed that the total population;is constant through time then there is a complementary recovered (or immune) subpopulation present which obeys the differential equationwith. The solution is: The following result links the above SIS model with a complementary recovered subpopulation to the generic one (1) under a minimum number of initial susceptible and sufficiently large number of initial infectious with initial growing rate.
Theorem 4. Assume that,and.
Then, the following properties hold:
- (i)
and,
- (ii)
is strictly decreasing onwith,
- (iii)
is strictly increasing on, and
with,
- (iv)
There iswhich is an undulation and, furthermore, strict inflection time instant of,
- (v)
Assume, in addition, thatis large enough to satisfy. Then, the epidemic model (26) can be written in the form (1) onwith the following function:
which is of the formwith;and any givenand;. - (vi)
The equilibrium points are,ifand, and,andwhich is only reachable ifsince, otherwise,.
Proof. Since
and
then
and
. Also,
if
. Property (i) has been proved. Furthermore,
implies from (27) that
is strictly decreasing on
where
what proves Property (ii) with
. On the other hand and since
is continuous, there exists some
such that
with
if and only if
. From (26),
and
for
since
. On the other hand, one has from (26) and (28) that:
and
has a relative maximum
at
which is also the absolute maximum on
. Property (iii) has been proved. Note also that since
is continuous and
, there exists some
such that
is an undulation point of
. Note furthermore that
From Lemma 1(i),
;
and some
implies that
is also an inflection time instant of
. The equivalent logic contrapositive proposition establishes that:
Then, if ; and some then is in fact an inflection time instant of . Assume that there is some arbitrarily small such that
Then:
; .
Since
is continuous on
and one gets that
It is known that so that, for some arbitrarily small such that , there are and with such that the following joint constraints hold:
- (1)
; with being strictly increasing on .
- (2)
Then, one gets from Condition 2 that:
so that
is not strictly increasing on
, hence a contradiction. As a result, the undulation time instant
of
is also a strict inflection time instant of
since
since Lemma 1 (ii) holds and the first zero of
occurs at
. Property (iv) has been proved. To prove Property (v), note that Equation (30) follows from (26)–(27). Now, we equalize (30) to (1) to get admissible functions
leading to:
and note that
. Note also that
from the use of (31) in (30) implies that
irrespective of
while
is chosen arbitrary and continuous time-differentiable subject to
and
,
(so that
) with
for
.
Now, note that
is a primary
—type indetermination which is resolved through L´H
pital rule leading to:
Since
then for sufficiently large
such that
then:
fulfilling, in particular:
Property (v) has been proved. Property (vi) is obvious by zeroing (26). □
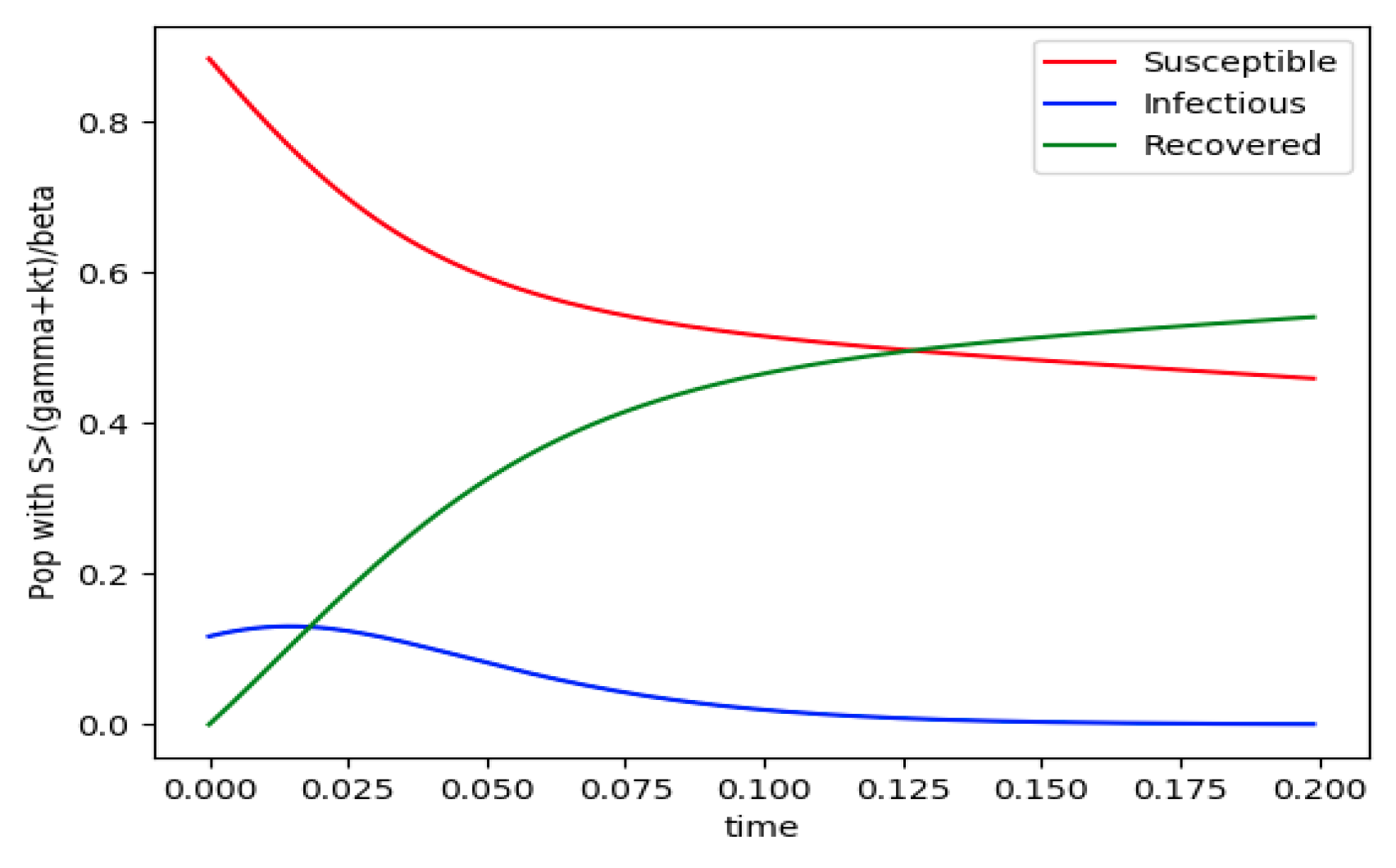
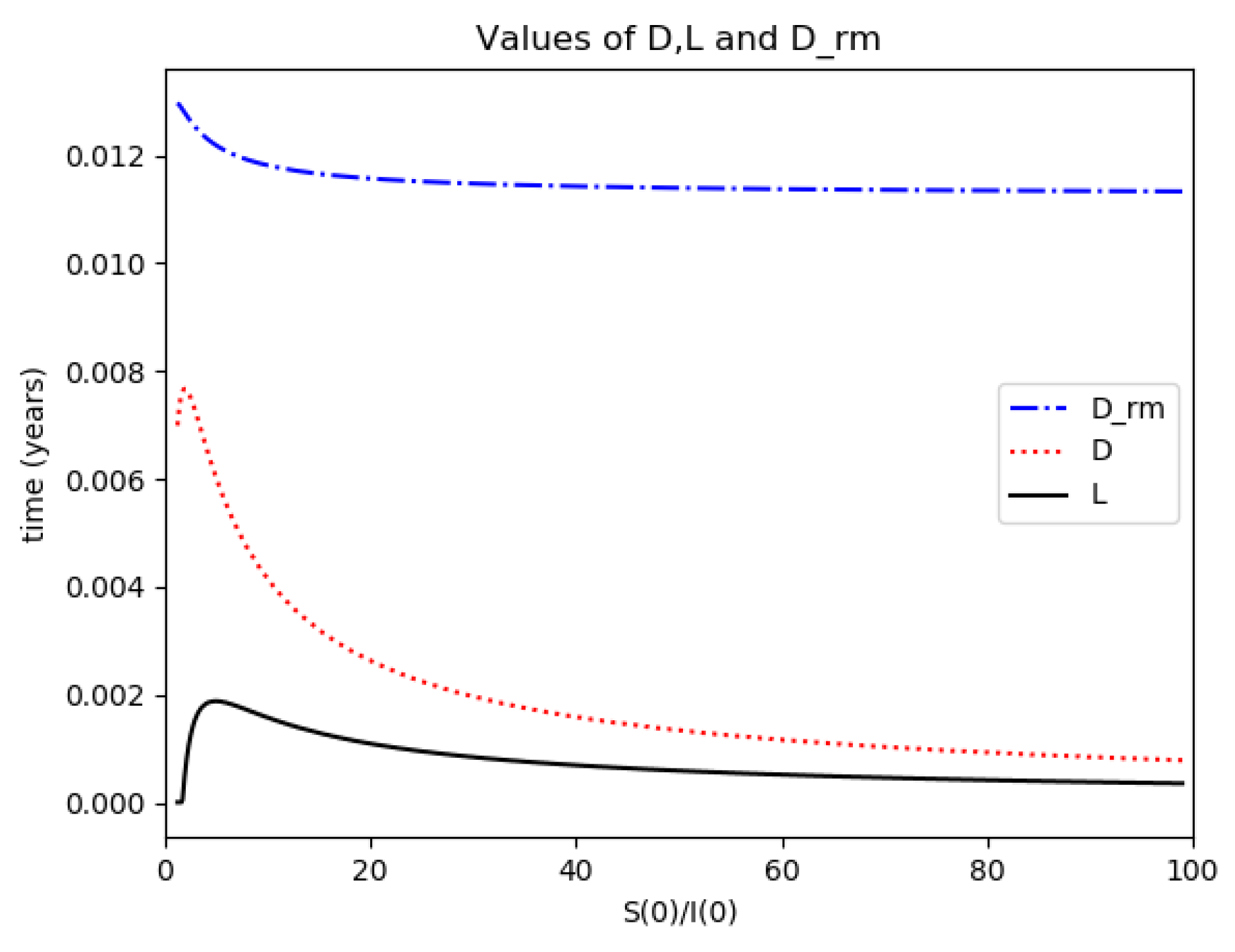
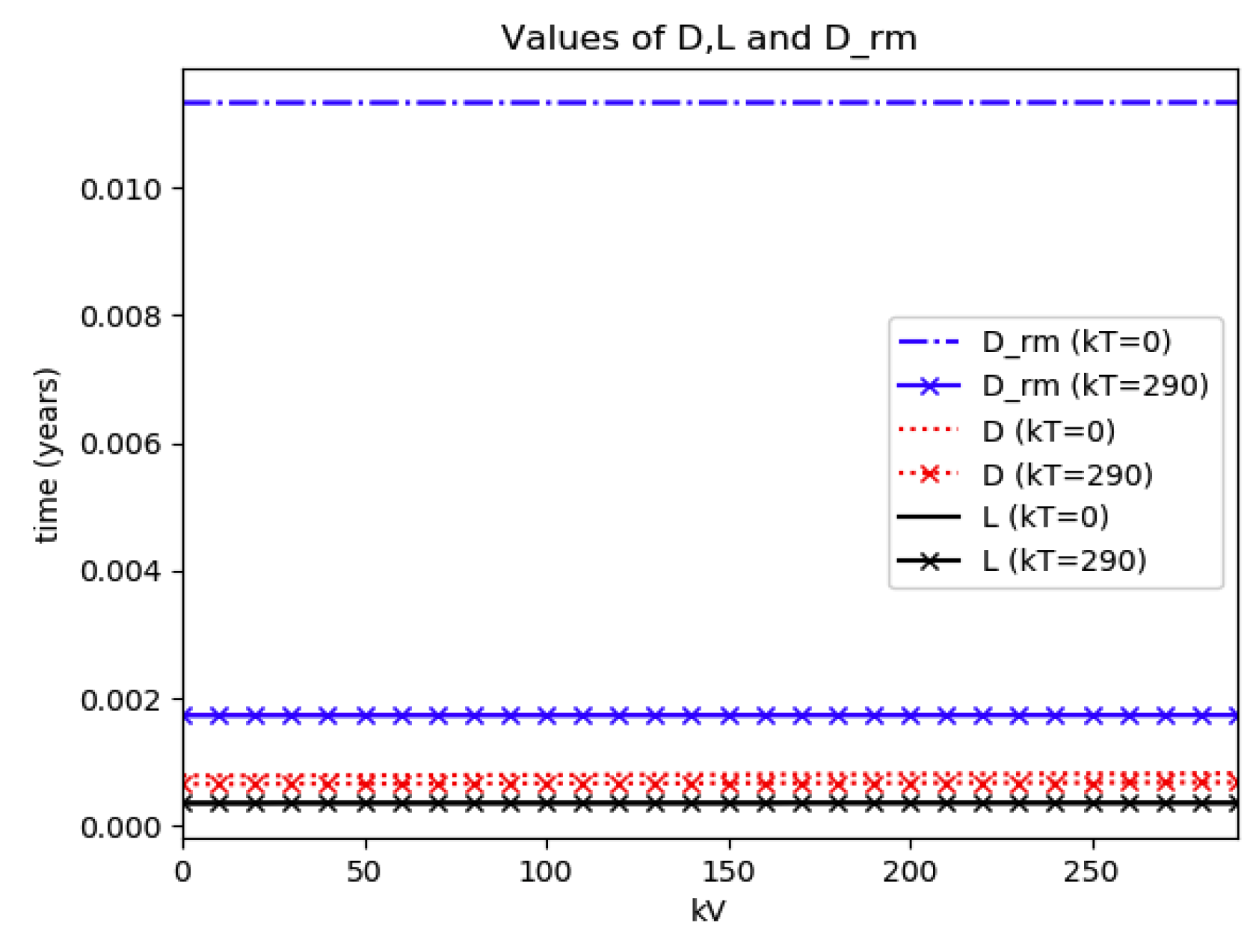
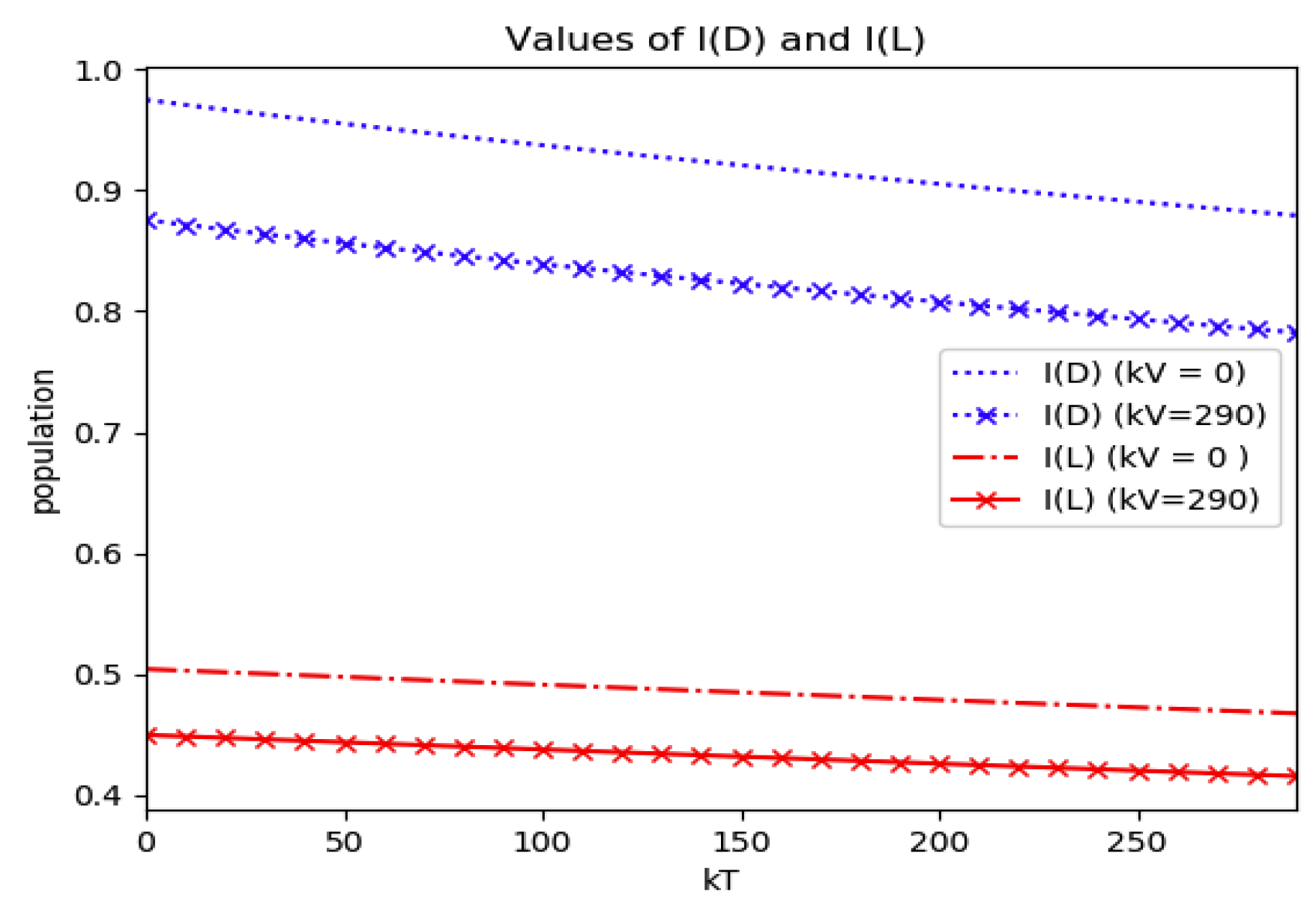
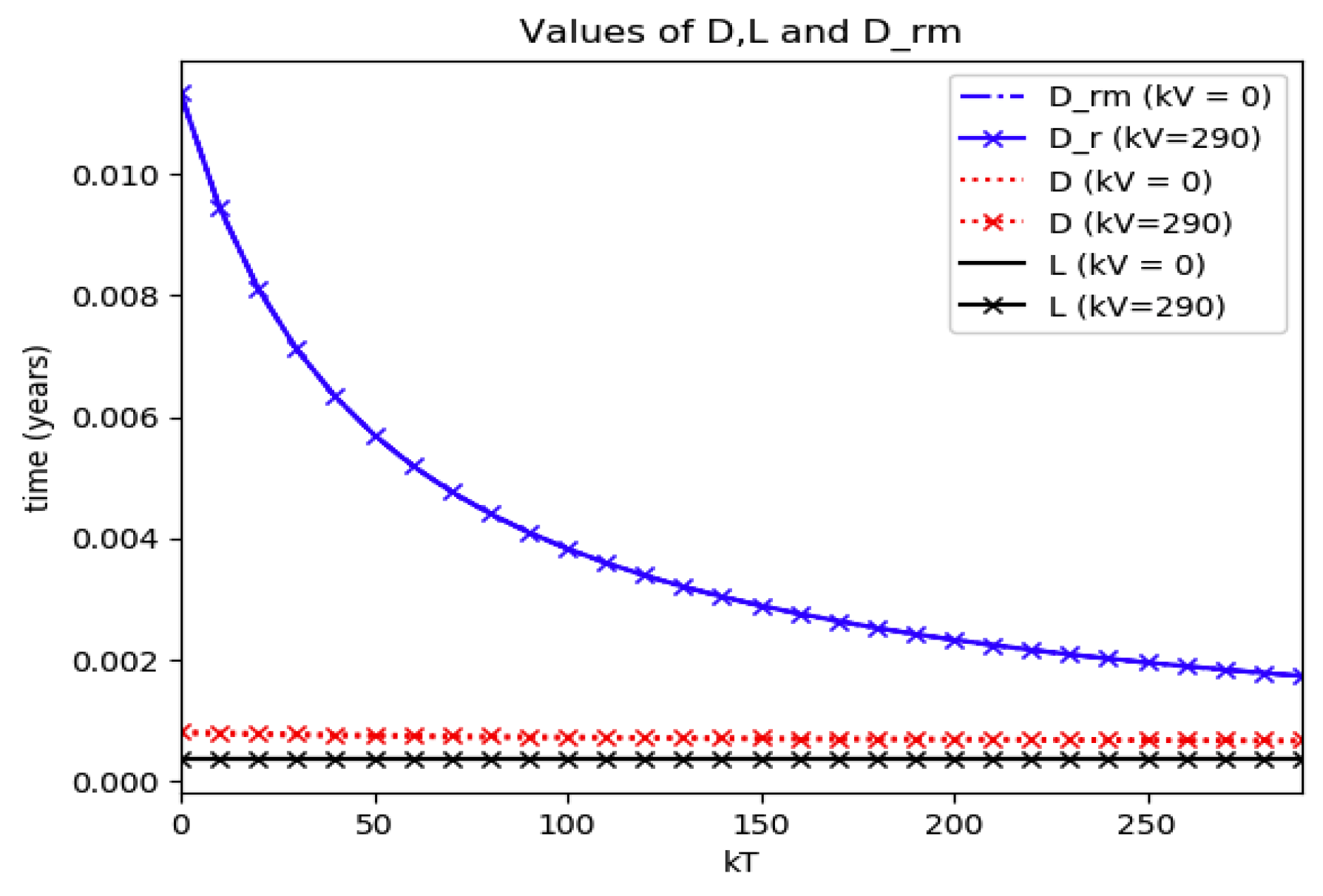
Example 4 is tested numerically in the sequel with the following data
β = 30,
γ = 50 years
−1, implying that the average infectious period is
Tγ = 365/50 = 7.3 days,
kV = 1 and
kT = 50. The time scale of the figures is in a scale of years accordingly with the above numerical values. In
Figure 1, the solution trajectories of all the subpopulation are shown with the constraints of Theorem 4 being fulfilled by the initial conditions, in particular
,
and
so that
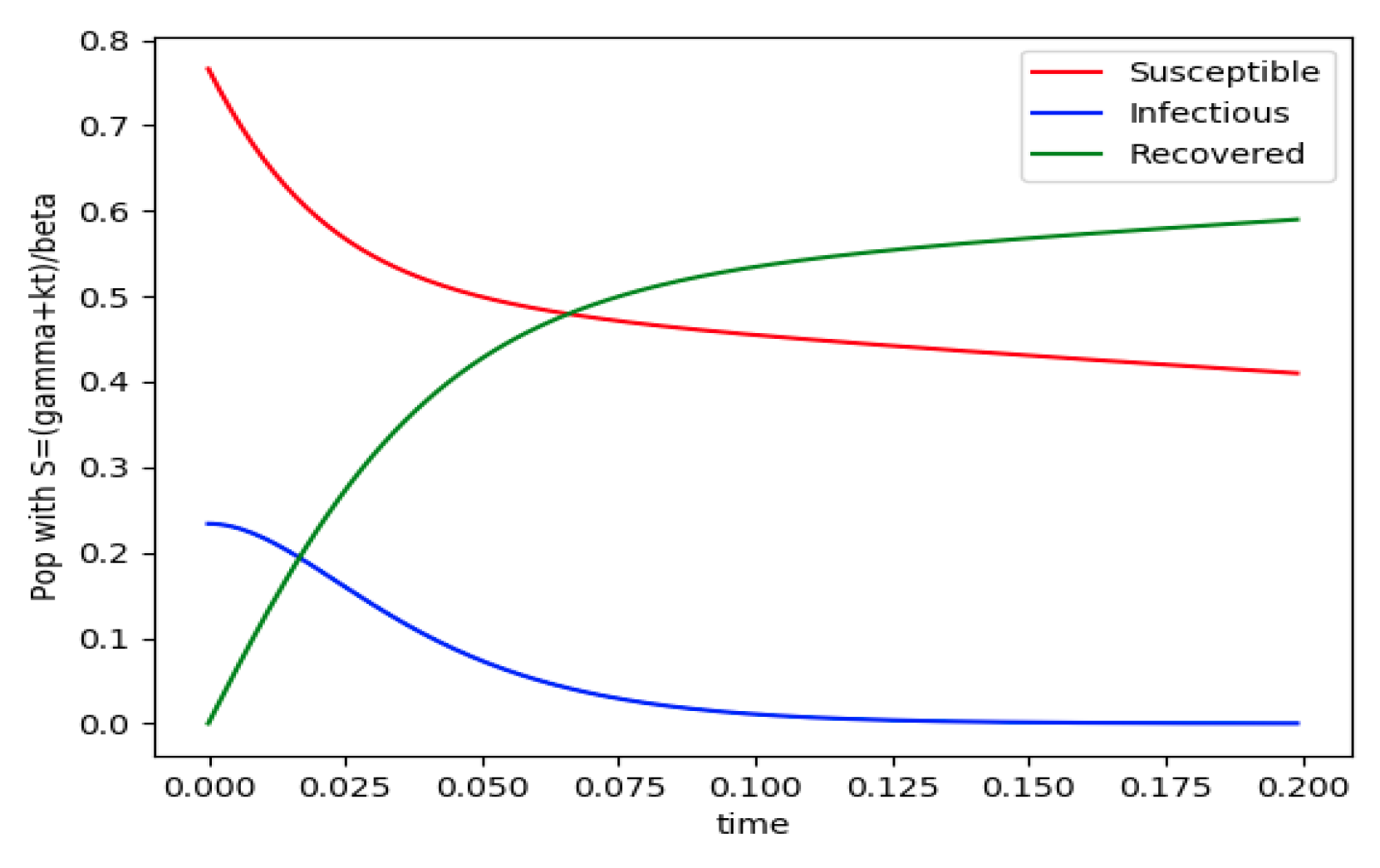
is normalized to unity. It is seen that the infectious subpopulation trajectory has a maximum at a finite time and that the state trajectory solution converges asymptotically to an endemic equilibrium point. In
Figure 2, the state trajectory solution is shown with
when
which violates the conditions of Theorem 4 with
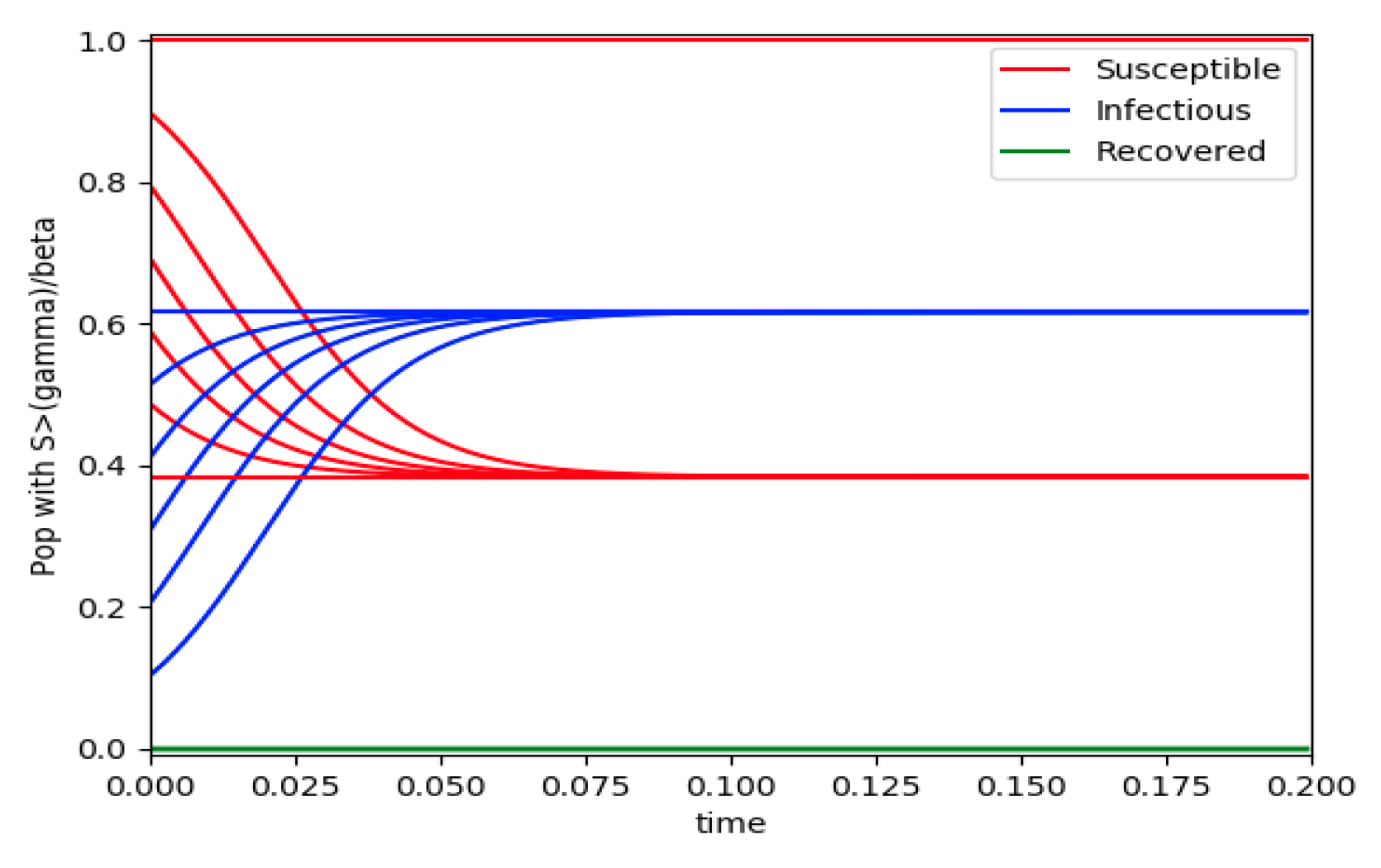
. In this case, there is no relative maximum of the infectious subpopulation at finite time. In both situations, it has been observed by extending the overall simulation time that the susceptible and the infectious subpopulations converge asymptotically to zero while the recovered subpopulation converges to unity as time tends to infinity. The controls are suppressed in
Figure 3 with
N0 = 1. In this case, the recovered subpopulation may be deleted from the model since it is unnecessary while being identically zero. The infectious and susceptible subpopulations are in an endemic equilibrium point for all time so that the infection results to be permanent in the sense that it cannot be asymptotically removed. See Theorem 4(vi) for the case
kV= 0.
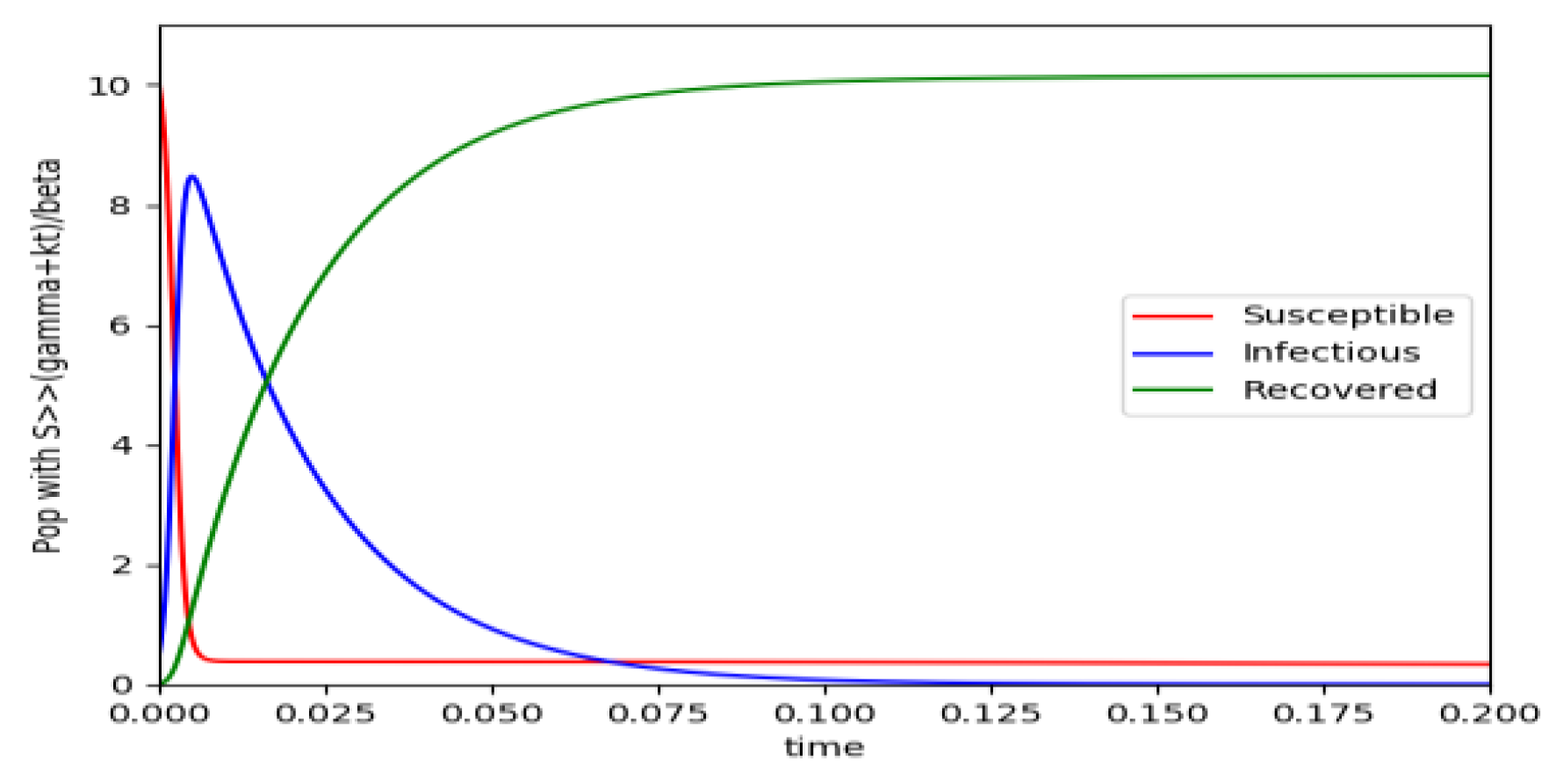
Figure 4 exhibits a trajectory solution which agrees with Theorem 4 while there is no normalization of the initial conditions to unity. In this case, the maximum of the infectious subpopulation at a finite time becomes very apparent.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}













