Alteration of the Intra- and Inter-Lobe Connectivity of the Brain Structural Network in Normal Aging


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Image Acquisition and Cortical Feature–Based Structural Network
2.3. FD Analysis and Brain Structural Network
2.4. Modular Analysis
2.4.1. Intra-Modular Connectivity Analysis
2.4.2. Inter-Modular Connectivity Analysis
2.5. Statistical Analysis
3. Results
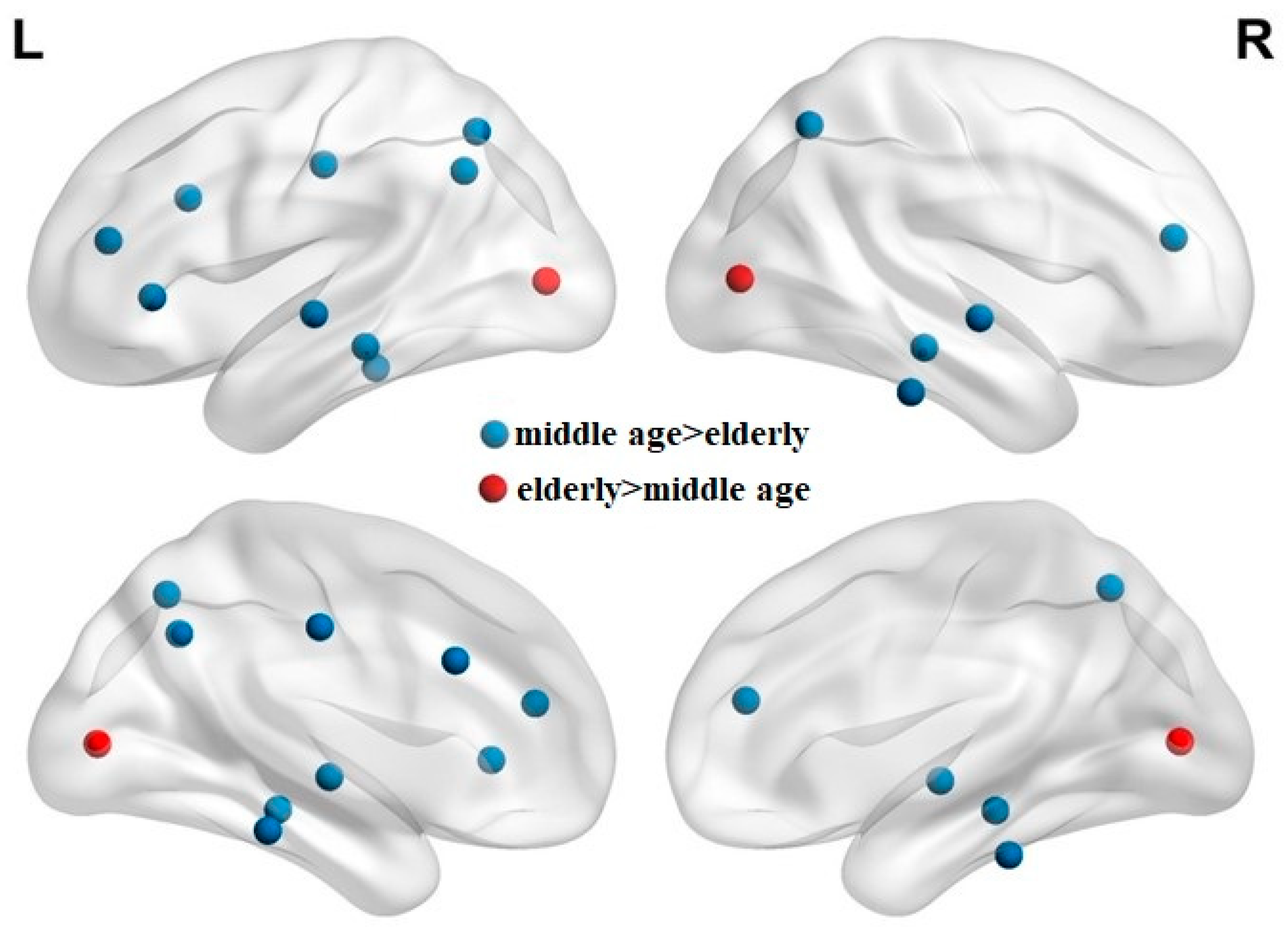
3.1. Healthy Elderly Group Exhibited More Significant Atrophy in the Left Brain Hemisphere
3.2. Elderly Group Exhibited Lower Modularity Values, Less Dense Modules, and More Separated Networks
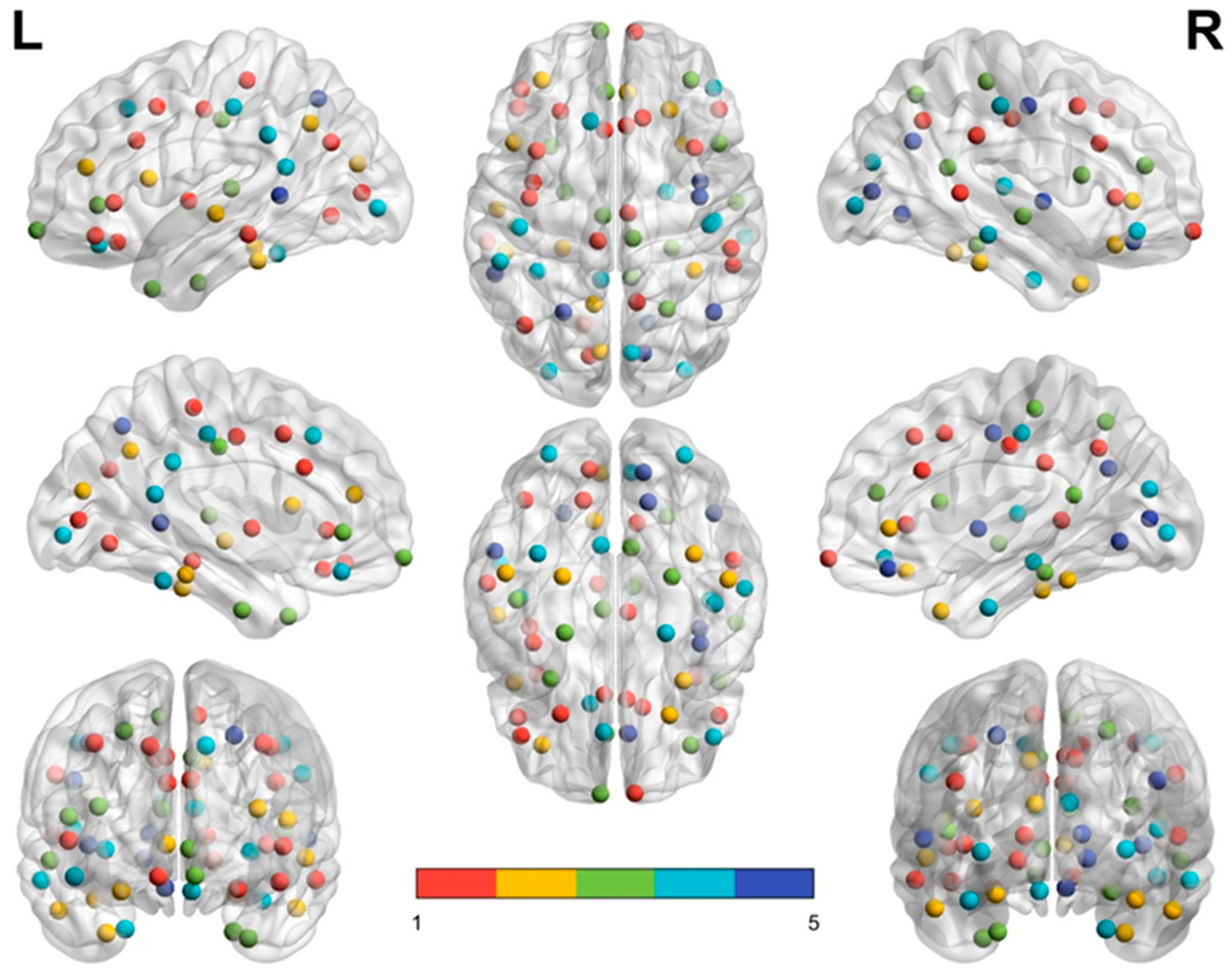
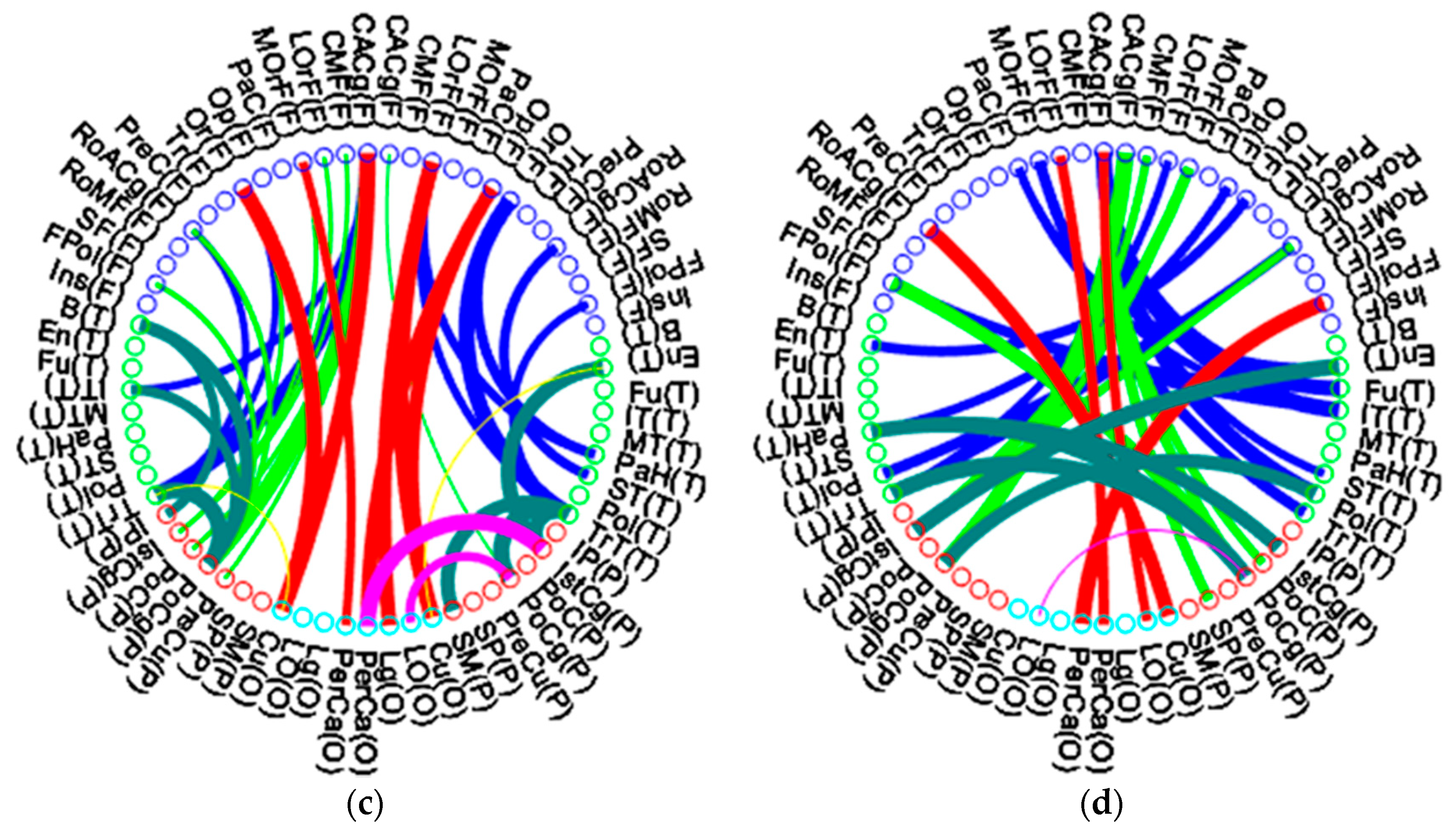
3.3. Modular Structures of the Middle-Aged Adult Brain Network
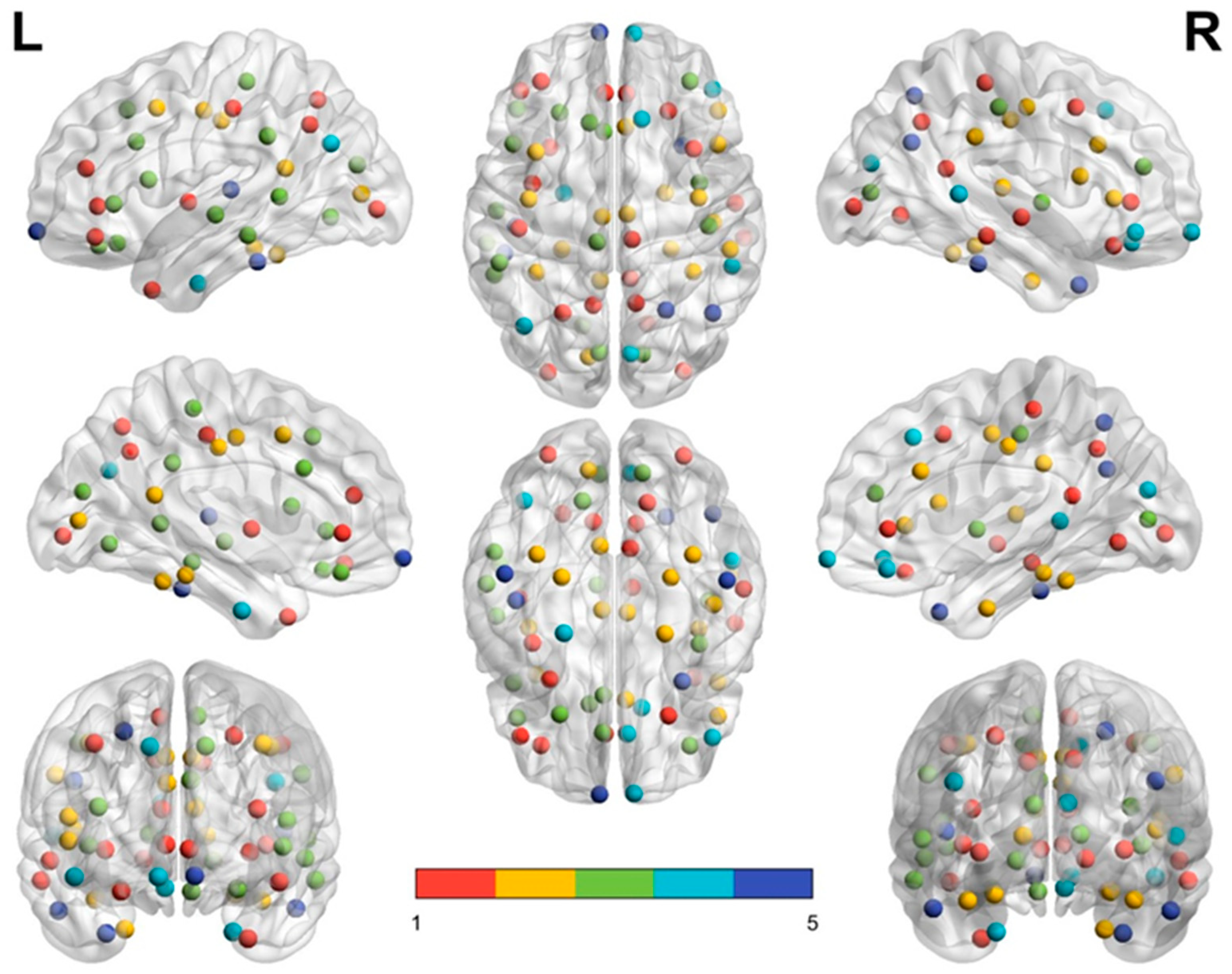
3.4. Elderly Group Revealed Significantly Decreased Intra-Modular Connectivity in Frontal and Parietal Lobes and Increased Intra-Modular Connectivity in Temporal and Occipital lobes
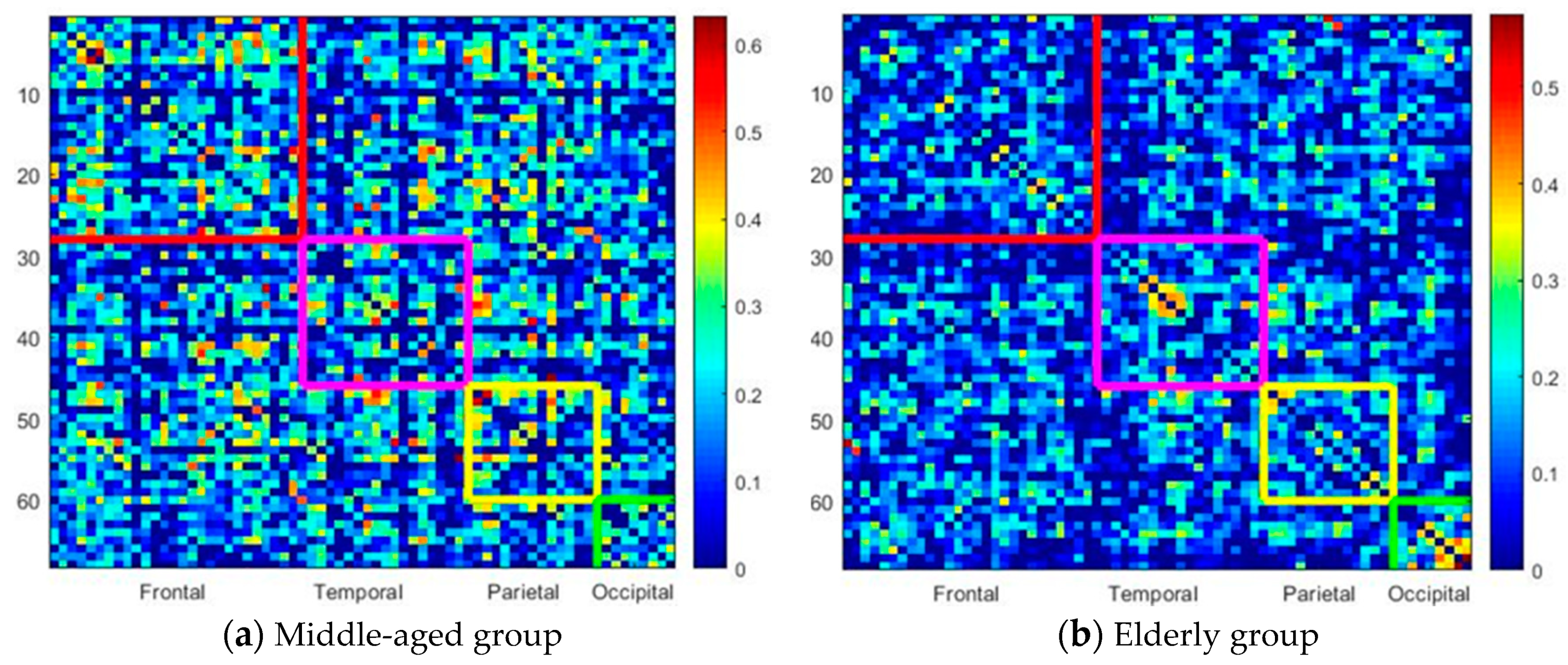
3.5. Elderly Group Revealed Significantly Decreased Inter-Lobe Connectivity in All Lobes
4. Discussion
4.1. Fractal Dimension-Based Brain Networks Can Manifest Morphological and Functional Modifications in Normal Aging
4.2. Compensation Mechanism of Intra-Lobe Connectivity of Temporal and Occipital Lobes in Normal Aging
4.3. New Wiring and Reconfiguring Modules Occurred within the Brain Structural Network during Normal Aging
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Morrison, J.H.; Hof, P.R. Life and death of neurons in the aging brain. Science 1997, 278, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Y.; Grossman, R.I.; Babb, J.S.; Rabin, M.L.; Mannon, L.J.; Kolson, D.L. Age-related total gray matter and white matter changes in normal adult brain, part I: Volumetric MR imaging analysis. Am. J. Neuroradiol. 2002, 23, 1327–1333. [Google Scholar] [PubMed]
- Winkler, A.M.; Kochunov, P.; Blangero, J.; Almasy, L.; Zilles, K.; Fox, P.T. Cortical thickness or greymatter volume? The importance of selecting the phenotype for imaging genetics studies. NeuroImage 2010, 53, 1135–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaitre, H.; Goldman, A.L.; Sambataro, F.; Verchinski, B.A.; Meyer-Lindenberg, A.; Weinberger, D.R.; Mattay, V.S. Normal age-related brain morphometric changes: Nonuniformity across cortical thickness, surface area and grey matter volume? Neurobiol. Aging 2012, 33, 617.e1–617.e9. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, H. Voxel-based Morphometry of Brain MRI in Normal Aging and Alzheimer’s Disease. Aging Dis. 2013, 4, 29–37. [Google Scholar]
- Wu, Y.T.; Shyu, K.K.; Jao, C.W.; Wang, Z.Y.; Soong, B.W.; Wu, H.M.; Wang, P.S. Fractal dimension analysis for quantifying cerebellar morphological change of multiple system atrophy of the cerebellar type (MSA-C). NeuroImage 2010, 49, 539–551. [Google Scholar] [CrossRef]
- Madan, C.R.; Kensinger, E.A. Cortical complexity as a measure of age-related brain atrophy. Neuroimage 2016, 134, 617–629. [Google Scholar] [CrossRef] [Green Version]
- Im, K.; Lee, J.M.; Lee, J.; Shin, Y.W.; Kim, I.Y.; Kwon, J.S.; Kim, S.I. Gender difference analysis of cortical thickness in healthy young adults with surface-based methods. Neuroimage 2006, 31, 31–38. [Google Scholar] [CrossRef]
- McKay, D.R.; Knowles, E.E.; Winkler, A.A.; Sprooten, E.; Kochunov, P.; Olvera, R.L.; Curran, J.E.; Kent, J.W.; Carless, M.A.; Harald, H.H.; et al. Influence of age, sex and genetic factors on the human brain. Brain Imaging Behav. 2014, 8, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Storsve, A.B.; Fjell, A.M.; Tamnes, C.K.; Westlye, L.T.; Overbye, K.; Aasland, H.W.; Walhovd, K.B. Differential longitudinal changes in cortical thickness, surface area and volume across the adult life span: Regions of accelerating and decelerating change. J. Neurosci. 2014, 34, 8488–8498. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, S.J.; Cox, S.R.; Shen, X.; Lombardo, M.V.; Reus, L.M.; Alloza, C.; Harris, M.A.; Alderson, H.L.; Hunter, S.; Neilson, E.; et al. Sex Differences in the Adult Human Brain: Evidence from 5216 UK Biobank Participants. Cereb. Cortex. 2018, 28, 2959–2975. [Google Scholar] [CrossRef]
- Mandelbrot, B. How long is the coast of Britain? Statistical self-similarity and fractional dimension. Science 1967, 156, 636–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteban, F.J.; Sepulcre, J.; de Mendizábal, N.V.; Goñi, J.; Navas, J.; de Miras, J.R.; Villoslada, P. Fractal dimension and white matter changes in multiple sclerosis. Neuroimage 2007, 36, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Esteban, F.J.; Sepulcre, J.; de Miras, J.R.; Navas, J.; de Mendizábal, N.V.; Goñi, J.; Villoslada, P. Fractal dimension analysis of grey matter in multiple sclerosis. J. Neurol. Sci. 2009, 282, 67–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.F.; Zhang, X.H.; Zou, T.X.; Huang, N.X.; Chen, H.J. Reduced Cortical Complexity in Cirrhotic Patients with Minimal Hepatic Encephalopathy. Neural Plast. 2020, 2020, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, R.D.; George, A.T.; Jeon, T.; Hynan, L.S.; Youn, T.S.; Kennedy, D.N. Alzheimer’s Disease Neuroimaging Initiative. Characterization of atrophic changes in the cerebral cortex using fractal dimensional analysis. Brain Imaging Behav. 2009, 3, 154–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collantoni, E.; Madan, C.R.; Meneguzzo, P.; Chiappini, I.; Tenconi, E.; Manara, R.; Favaro, A. Cortical Complexity in Anorexia Nervosa: A Fractal Dimension Analysis. J. Clin. Med. 2020, 9, 833. [Google Scholar] [CrossRef] [Green Version]
- Sporns, O.; Chialvo, D.R.; Kaiser, M.; Hilgetag, C.C. Organization, development and function of complex brain networks. Trends Cogn. Sci. 2004, 8, 418–425. [Google Scholar] [CrossRef] [Green Version]
- Fortunato, S.; Barthelemy, M. Resolution limit in community detection. Proc. Natl. Acad. Sci. USA 2007, 104, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.J.; He, Y.; Rosa-Neto, P.; Germann, J.; Evans, A.C. Revealing modular architecture of human brain structural networks by using cortical thickness from MRI. Cereb. Cortex 2008, 18, 2374–2381. [Google Scholar] [CrossRef]
- Luo, Y.G.; Wang, D.; Liu, K.; Weng, J.; Guan, Y.; Chan, K.C.C.; Chu, W.C.W.; Shi, L. Brain Structure Network Analysis in Patients with Obstructive Sleep Apnea. PLoS ONE 2015, 10, e0139055. [Google Scholar] [CrossRef] [Green Version]
- Sanabria-Diaz, G.; Melie-García, L.; Iturria-Medina, Y.; Alemán-Gómez, Y.; Hernández-González, G.; Valdés-Urrutia, L.; Valdés-Sosa, P. Surface area and cortical thickness descriptors reveal different attributes of the structural human brain networks. Neuroimage 2010, 50, 1497–1510. [Google Scholar] [CrossRef]
- Seeley, W.W. Selective functional, regional, and neuronal vulnerability in frontotemporal dementia. Curr. Opin. Neurol. 2008, 21, 701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Dagher, A.; Chen, Z.; Charil, A.; Zijdenbos, A.; Worsley, K.; Evans, A. Impaired small-world efficiency in structural cortical networks in multiple sclerosis associated with white matter lesion load. Brain 2009, 132, 3366–3379. [Google Scholar] [CrossRef] [Green Version]
- Supekar, K.; Menon, V.; Rubin, D.; Musen, M.; Greicius, M.D. Network analysis of intrinsic functional brain connectivity in Alzheimer’s disease. PLoS Comput. Biol. 2008, 4, e1000100. [Google Scholar] [CrossRef] [PubMed]
- Jao, C.W.; Soong, B.W.; Wang, T.Y.; Wu, H.M.; Lu, C.F.; Wang, P.S.; Wu, Y.T. Intra-and Inter-Modular Connectivity Alterations in the Brain Structural Network of Spinocerebellar Ataxia Type 3. Entropy 2019, 21, 317. [Google Scholar] [CrossRef] [Green Version]
- Desikan, R.S.; Ségonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Albert, M.S. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage 2006, 31, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.H.; Yoon, U.; Lee, K.J.; Shin, Y.W.; Lee, J.M.; Kim, I.Y.; Ha, K.S.; Kim, S.I.; Kwon, J.S. Fractal dimension of cerebral cortical surface in schizophrenia and obsessive–compulsive disorder. Neurosci. Lett. 2005, 384, 172–176. [Google Scholar] [CrossRef]
- Sandu, A.L.; Rasmussen, I.A.; Lundervold, A.; Frank, K.F.; Neckelmann, G.; Hugdahl, K.; Specht, K. Fractal dimension analysis of MR images reveals grey matter structure irregularities in schizophrenia. Comput. Med. Imaging Graph. 2008, 32, 150–158. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.E. Modularity and community structure in networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef] [Green Version]
- Meunier, D.; Lambiotte, R.; Fornito, A.; Ersche, K.D.; Bullmore, E.T. Hierarchical modularity in human brain functional networks. Front. Neuroinform. 2009, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimerà, R.; Amaral, L.A.N. Functional Cartography of Complex Metabolic Networks. Nature 2005, 433, 895–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullmore, E.T.; Suckling, J.; Overmeyer, S.; Rabe-Hesketh, S.; Taylor, E.; Brammer, M.J. Global, voxel, and cluster tests, by theory and permutation, for a difference between two groups of structural MR images of the brain. IEEE Trans. Med. Imaging 1999, 18, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Duncan, J.; Owen, A.M. Common regions of the human frontal lobe recruited by diverse cognitive demands. Trends Neurosci. 2000, 23, 475–483. [Google Scholar] [CrossRef]
- Xia, M.; Wang, J.; He, Y. BrainNet Viewer: A network visualization tool for human brain connectomics. PLoS ONE 2013, 8, e68910. [Google Scholar] [CrossRef] [Green Version]
- Raz, N.; Lindenberger, U.; Rodrigue, K.M.; Kennedy, K.M.; Head, D.; Williamson, A.; Acker, J.D. Regional brain changes in aging healthy adults: General trends, individual differences and modifiers. Cereb. Cortex 2005, 15, 1676–1689. [Google Scholar] [CrossRef]
- Song, J.; Birn, R.M.; Boly, M.; Meier, T.B.; Nair, V.A.; Meyerand, M.E.; Prabhakaran, V. Age-related reorganizational changes in modularity and functional connectivity of human brain networks. Brain Connect. 2014, 4, 662–676. [Google Scholar] [CrossRef]
- Meier, J.; Tewarie, P.; Van Mieghem, P. The union of shortest path trees of functional brain networks. Brain Connect. 2015, 5, 575–581. [Google Scholar] [CrossRef]
- Mišić, B.; Betzel, R.F.; De Reus, M.A.; Van Den Heuvel, M.P.; Berman, M.G.; McIntosh, A.R.; Sporns, O. Network-level structure-function relationships in human neocortex. Cereb. Cortex 2016, 26, 3285–3296. [Google Scholar] [CrossRef] [Green Version]
- Farahibozorg, S.; Hashemi-Golpayegani, S.M.; Ashburner, J. Age-and sex-related variations in the brain white matter fractal dimension throughout adulthood: An MRI study. Clin. Neuroradiol. 2015, 25, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.; Taki, Y.; Sato, K.; Kinomura, S.; Goto, R.; Okada, K.; Kawashima, R.; He, Y.; Evans, A.C.; Fukuda, H. Age-related changes in topological organization of structural brain networks in healthy individuals. Hum. Brain Mapp. 2012, 33, 552–568. [Google Scholar] [CrossRef] [PubMed]
- Douaud, G.; Groves, A.R.; Tamnes, C.K.; Westlye, L.T.; Duff, E.P.; Engvig, A.; Matthews, P.M. A common brain network links development, aging, and vulnerability to disease. Proc. Natl. Acad. Sci. USA 2014, 111, 17648–17653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geerligs, L.; Renken, R.J.; Saliasi, E.; Maurits, N.M.; Lorist, M.M. A brain-wide study of age-related changes in functional connectivity. Cereb. Cortex 2015, 25, 1987–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.; Ratnarajah, N.; Tuan, T.A.; Chen, S.H.A.; Qiu, A. Adaptation of brain functional and structural networks in aging. PLoS ONE 2015, 10, e0123462. [Google Scholar] [CrossRef]
- Liu, K.; Yao, S.; Chen, K.; Zhang, J.; Yao, L.; Li, K.; Guo, X. Structural brain network changes across the adult lifespan. Front. Aging Neurosci. 2017, 9, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafkemeijer, A.; Altmann-Schneider, I.; de Craen, A.J.; Slagboom, P.E.; van der Grond, J.; Rombouts, S.A. Associations between age and gray matter volume in anatomical brain networks in middle-aged to older adults. Aging Cell 2014, 13, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Puxeddu, M.G.; Faskowitz, J.; Betzel, R.F.; Petti, M.; Astolfi, L.; Sporns, O. The modular organization of brain cortical connectivity across the human lifespan. NeuroImage 2020, 218, 116974. [Google Scholar] [CrossRef]
- Catani, M.; Dell’Acqua, F.; Vergani, F.; Malik, F.; Hodge, H.; Roy, P.; De Schotten, M.T. Short frontal lobe connections of the human brain. Cortex 2012, 48, 273–291. [Google Scholar] [CrossRef]
- Bender, A.R.; Völkle, M.C.; Raz, N. Differential aging of cerebral white matter in middle-aged and older adults: A seven-year follow-up. Neuroimage 2016, 125, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Pareek, V.; Rallabandi, V.S.; Roy, P.K. A Correlational Study between Microstructural White Matter Properties and Macrostructural Gray Matter Volume across Normal Ageing: Conjoint DTI and VBM Analysis. Magn. Reson. Insights 2018, 11, 1178623X18799926. [Google Scholar] [CrossRef] [Green Version]
- Grydeland, H.; Vértes, P.E.; Váša, F.; Romero-Garcia, R.; Whitaker, K.; Alexander-Bloch, A.F.; Westlye, L.T. Waves of maturation and senescence in micro-structural MRI markers of human cortical myelination over the lifespan. Cereb. Cortex 2019, 29, 1369–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, S.R.; Ritchie, S.J.; Tucker-Drob, E.M.; Liewald, D.C.; Hagenaars, S.P.; Davies, G.; Deary, I.J. Ageing and brain white matter structure in 3,513 UK Biobank participants. Nat. Commun. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, S.W.; Dennis, N.A.; Buchler, N.G.; White, L.E.; Madden, D.J.; Cabeza, R. Assessing the effects of age on long white matter tracts using diffusion tensor tractography. Neuroimage 2009, 46, 530–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freret, T.; Gaudreau, P.; Schumann-Bard, P.; Billard, J.M.; Popa-Wagner, A. Mechanisms underlying the neuroprotective effect of brain reserve against late life depression. J. Neural Transm. 2015, 122, 55–61. [Google Scholar] [CrossRef]
- Fratiglioni, L.; Wang, H.X. Brain reserve hypothesis in dementia. J. Alzheimer’s Dis. 2007, 12, 11–22. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Frontal | ROI | Abbreviation | Temporal | ROI | Abbreviation |
|---|---|---|---|---|---|
| 1, 2 | Caudal anterior cingulate | CACg | 37, 38 | Middle temporal | MT |
| 3, 4 | Caudal middle frontal | CMF | 39, 40 | Parahippocampal | PaH |
| 5, 6 | Lateral orbito frontal | LOrF | 41, 42 | Superior temporal | ST |
| 7, 8 | Medial orbito frontal | MOrF | 43, 44 | Temporal pole | TPol |
| 9, 10 | Paracentral | PaC | 45, 46 | Transverse temporal | TrT |
| 11, 12 | Parsopercularis | Op | Parietal | ||
| 13, 14 | Parsorbitalis | Or | 47, 48 | Inferior parietal | IP |
| 15, 16 | Parstriangularis | Tr | 49, 50 | Isthmus cingulate | IstCg |
| 17, 18 | Precentral | PreC | 51, 52 | Postcentral | PoC |
| 19, 20 | Rostral anterior cingulate | RoACg | 53, 54 | Posterior cingulate | PoCg |
| 21, 22 | Rostral middle frontal | RoMF | 55, 56 | Precuneus | PreCu |
| 23, 24 | Superior frontal | SF | 57, 58 | Superior parietal | SP |
| 25, 26 | Frontalpole | FPol | 59, 60 | Supra marginal | SM |
| 27, 28 | Insula | Ins | Occipital | ||
| Temporal | 61, 62 | Cuneus | Cu | ||
| 29, 30 | Bankssts | B | 63, 64 | Lateral occipital | LO |
| 31, 32 | Entorhinal | En | 65, 66 | Lingual | Lg |
| 33, 34 | Fusiform | Fu | 67, 68 | Pericalcarine | PerCa |
| 35, 36 | Inferior temporal | IT |
| FD Measure of Lobes between Middle-Aged and Elderly Groups | Significantly Different Parcellated Regions | ||||||
|---|---|---|---|---|---|---|---|
| Lobes (L/R) | Middle-Age | Elderly | p Value | Regions | Middle-Age | Elderly | p Value |
| Frontal (F) | 2.2399 ± 0.1035 | 2.2326 ± 0.1032 | 0.0221 | F_CACg(L) | 2.1081 | 2.0668 | 0.037 |
| Frontal (L) | 2.2375 ± 0.1091 | 2.2294 ± 0.1109 | 0.0336 | F_Tr(L) | 2.2499 | 2.2334 | 0.044 |
| Frontal (R) | 2.2423 ± 0.1017 | 2.2358 ± 0.0989 | 0.0374 | F_RoMF(L) | 2.4128 | 2.3997 | 0.005 |
| Temporal (T) | 2.1991 ± 0.1232 | 2.1897 ± 0.1244 | 0.0084 | F_RoMF(R) | 2.4128 | 2.4014 | 0.048 |
| Temporal (L) | 2.2065 ± 0.1145 | 2.1968 ± 0.1171 | 0.003 | T_MT(L) | 2.3118 | 2.2989 | 0.043 |
| Temporal (R) | 2.1918 ± 0.1379 | 2.1825 ± 0.138 | 0.034 | T_MT(R) | 2.3254 | 2.3103 | 0.018 |
| Parietal (P) | 2.2972 ± 0.1047 | 2.2858 ± 0.1078 | 0.0005 | T_ST(L) | 2.3267 | 2.3148 | 0.033 |
| Parietal (L) | 2.2953 ± 0.1055 | 2.2833 ± 0.1109 | 0.0026 | T_ST(R) | 2.3392 | 2.329 | 0.037 |
| Parietal (R) | 2.2990 ± 0.1122 | 2.2882 ± 0.1135 | 0.00025 | T_PaH(L) | 2.0762 | 2.0536 | 0.035 |
| Occipital (O) | 2.2033 ± 0.1306 | 2.2134 ± 0.1139 | 0.0267 | T_IT(R) | 2.3278 | 2.3138 | 0.031 |
| Occipital (L) | 2.1908 ± 0.151 | 2.2061 ± 0.1283 | 0.007 | P_PoCg(L) | 2.1961 | 2.1684 | 0.038 |
| Occipital (R) | 2.2159 ± 0.1287 | 2.2208 ± 0.1170 | 0.348 | P_PreCu(L) | 2.3302 | 2.3171 | 0.045 |
| P_SP(L) | 2.3567 | 2.3487 | 0.031 | ||||
| P_SP(R) | 2.3567 | 2.3487 | 0.016 | ||||
| O_PerCa(L) | 2.0416 | 2.0935 | 0.0009 | ||||
| O_PerCa(R) | 2.0836 | 2.1097 | 0.04 | ||||
| Middle Age Group | Lobe (Parcellated Regions (Left, Right)) |
|---|---|
| Module 1 (22) | Frontal:CACg(L), CACg(R), CMF(L), CMF(R), LOrF(L), PaC(L), Or(L), Tr(L), Tr(R), PreC(L), PreC(R), RoACg(R), FPol(R), Ins(L) Temporal: MT(L), B(R), TPol(R) Parietal: IP(L), PoCg(R), SP(R) Occipital: Lg(L), PerCa(L) |
| Module 2 (14) | Frontal: MOrF(L), SF(L), Or(R), Temporal: Fu(L), En(R), MT(R), TrT(R) Parietal: PoC(L), SM(L), IstCg(L), PoC(R) Occipital: LO(L), LO(R), Cu(R) |
| Module 3 (13) | Frontal: RoACg(L), FPol(L), PaC(R), Op(R), SF(R), Temporal: En(L), TPol(L), TrT(L) PaH(R), ST(R) Parietal: PoCg(L), IstCg(R), SM(R) |
| Module 4 (11) | Frontal: Op(L), RoMF(L), RoMF(R), LOrF(R), Temporal: IT(L), PaH(L), ST(L), Fu(R), IT(R), Parietal: PreCu(L), Occipital: Cu(L) |
| Module 5 (8) | Frontal: MOrF(R), Ins(R), Temporal: B(L), Parietal: SP(L), IP(R), PreCu(R) Occipital: Lg(R), PerCa(R) |
| Elderly Group | Lobe (Parcellated Regions (Left, Right)) |
|---|---|
| Module 1 (19) | Frontal: Or(L), RoACg(L), RoACg(R), RoMF(L), Ins(L), CMF(R) LOrF(R) PaC(R) Temporal: TPol(L), MT(R), ST(R) Parietal: PoC(L), PreCu(L), SP(L), IstCg(R) preCu(R) Occipital: LO(L), LO(R), Lg(R) |
| Module 2 (17) | Frontal: CMF(L) PreC(L) PreC(R), CACg(R) OP(R) Tr(R) Temporal: Fu(L), PaH(L) En(R) Fu(R) PaH(R) TrT(R) Parietal: IstCg(L), PoCg(L), PoCg(R) SM(R), Occipital: PerCa(L) |
| Module 3 (17) | Frontal: CACg(L) LOrF(L), MOrF(L) PaC(L) Op(L) Tr(L) SF(L) RoMF(R) Ins(R) Temporal: B(L) MT(L) ST(L) Parietal: SM(L), PoC(R) Occipital: Cu(L), Lg(L), PerCa(R) |
| Module 4 (8) | Frontal: MOrF(R), Or(R), SF(R), FPol(R) Temporal: En(L), B(R), Parietal: IP(L), Occipital: Cu(R) |
| Module 5 (7) | Frontal: FPol(L) Temporal: IT(L) IT(R) TrT(L) TPol(R), Parietal: SP(R), IP(R) |
| Group | Frontal Lobe | Temporal Lobe | Parietal Lobe | Occipital Lobe | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | Left | Right | Total | Left | Right | Total | Left | Right | Total | Left | Right | |
| Middle age (M) | 0.3986 | 0.4552 | 0.3421 | 0.3351 | 0.3127 | 0.3575 | 0.3779 | 0.426 | 0.3298 | 0.3876 | 0.3551 | 0.4207 |
| Elderly (E) | 0.3686 | 0.4168 | 0.3211 | 0.4239 | 0.3867 | 0.4488 | 0.3378 | 0.3576 | 0.3195 | 0.4642 | 0.4309 | 0.5012 |
| Ratio (E/M) | 92.4% | 91.6% | 93.8% | 126.5% | 123.7% | 125.6% | 89.4% | 83.9% | 96.9% | 119.8% | 121.3% | 119.1% |
| Group | Frontal Lobe | Temporal Lobe | Parietal Lobe | Occipital Lobe | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | Left | Right | Total | Left | Right | Total | Left | Right | Total | Left | Right | |
| Middle age (M) | 0.7119 | 0.7131 | 0.7106 | 0.7088 | 0.7087 | 0.7089 | 0.7104 | 0.7099 | 0.7108 | 0.7134 | 0.7137 | 0.7131 |
| Elderly (E) | 0.6345 | 0.636 | 0.633 | 0.6546 | 0.658 | 0.6512 | 0.6265 | 0.6405 | 0.6125 | 0.6341 | 0.6383 | 0.630 |
| Ratio (E/M) | 89.1% | 89.2% | 89.1% | 92.3% | 92.8% | 91.9% | 88.1% | 90.2% | 86.1% | 88.8% | 89.4% | 88.4% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jao, C.-W.; Yeh, J.-H.; Wu, Y.-T.; Lien, L.-M.; Tsai, Y.-F.; Chu, K.-E.; Hsiao, C.-Y.; Wang, P.-S.; Lau, C.I. Alteration of the Intra- and Inter-Lobe Connectivity of the Brain Structural Network in Normal Aging. Entropy 2020, 22, 826. https://doi.org/10.3390/e22080826
Jao C-W, Yeh J-H, Wu Y-T, Lien L-M, Tsai Y-F, Chu K-E, Hsiao C-Y, Wang P-S, Lau CI. Alteration of the Intra- and Inter-Lobe Connectivity of the Brain Structural Network in Normal Aging. Entropy. 2020; 22(8):826. https://doi.org/10.3390/e22080826
Chicago/Turabian StyleJao, Chi-Wen, Jiann-Horng Yeh, Yu-Te Wu, Li-Ming Lien, Yuh-Feng Tsai, Kuang-En Chu, Chen-Yu Hsiao, Po-Shan Wang, and Chi Ieong Lau. 2020. "Alteration of the Intra- and Inter-Lobe Connectivity of the Brain Structural Network in Normal Aging" Entropy 22, no. 8: 826. https://doi.org/10.3390/e22080826
APA StyleJao, C. -W., Yeh, J. -H., Wu, Y. -T., Lien, L. -M., Tsai, Y. -F., Chu, K. -E., Hsiao, C. -Y., Wang, P. -S., & Lau, C. I. (2020). Alteration of the Intra- and Inter-Lobe Connectivity of the Brain Structural Network in Normal Aging. Entropy, 22(8), 826. https://doi.org/10.3390/e22080826