Molecular Docking Studies and Anti-Tyrosinase Activity of Thai Mango Seed Kernel Extract

Abstract
:Introduction
Results and Discussion
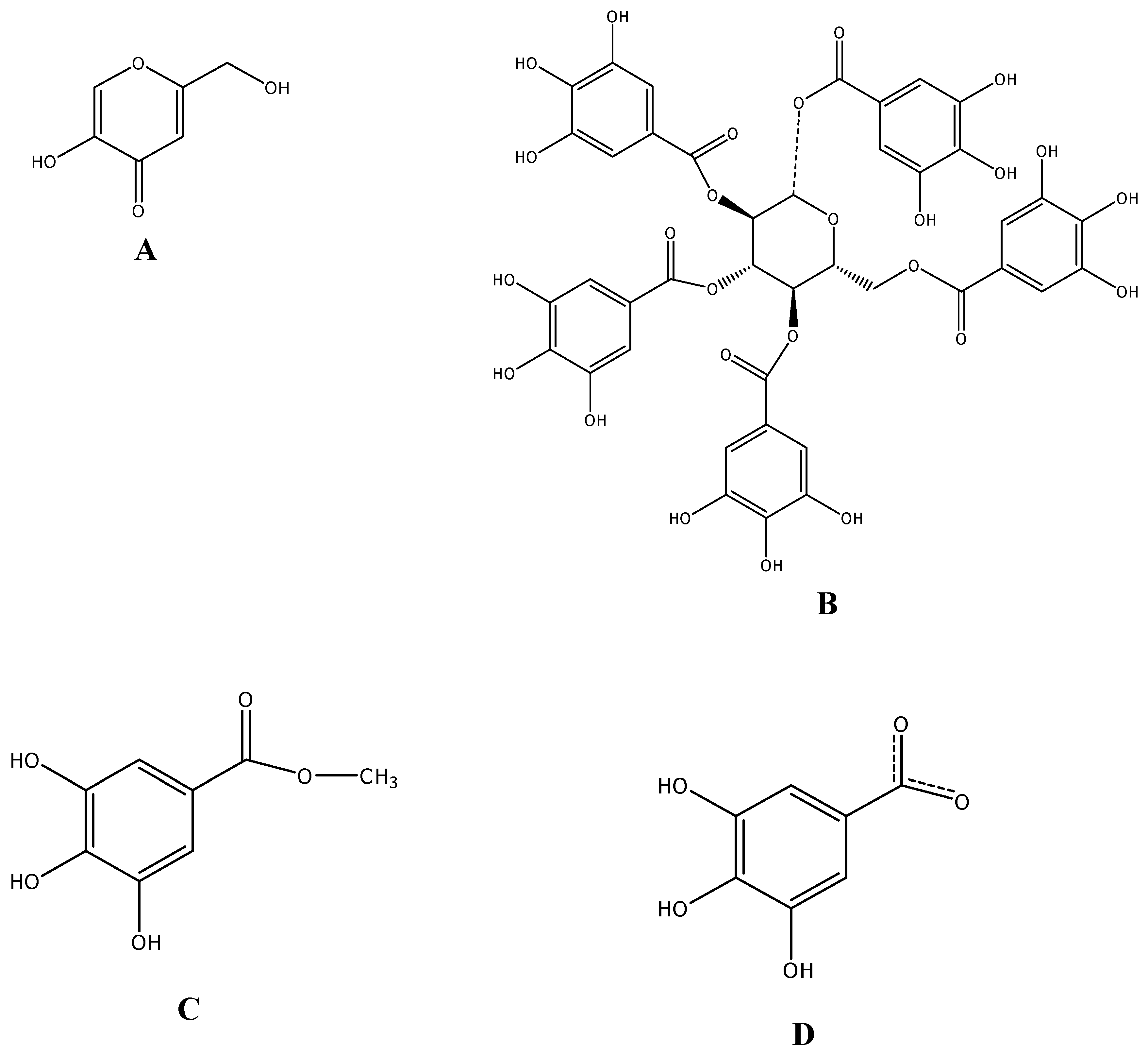
Effect of MSKE and its phenolic principles on the activity of mushroom tyrosinase


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test compounds | Anti-tyrosinase |
|---|---|
| IC50 ± SEM (µg/mL) | |
| MSKE | 98.63 ± 1.62 |
| Phenolic principles of MSKE [13]: | |
| GA | 644.00± 14.00 |
| MG | 62.50 ± 0.50 |
| PGG | 42.65 ± 1.85 |
| Positive reference: KA | 2.21 ± 0.05 |
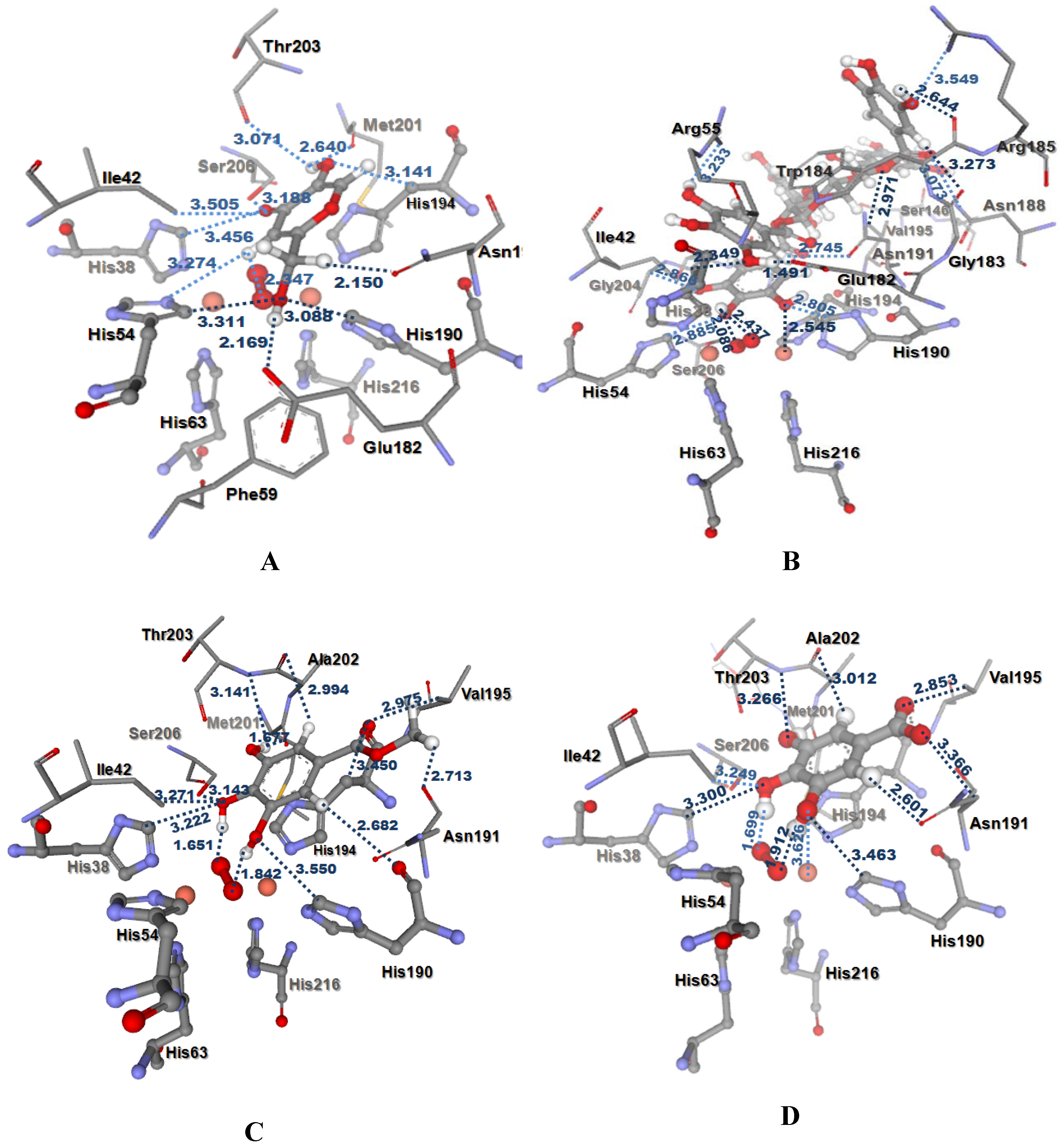
Molecular docking study


Conclusions
Experimental
Chemicals
Plant materials
Standardization
Determination of mushroom tyrosinase inhibition in vitro
Molecular modeling

Statistical analysis
Acknowledgements
References
- Kim, Y.-J.; Uyama, H. Tyrosinase inhibitors from natural and synthetic sources: structure, inhibition mechanism and perspective for the future. Cell. Mol. Life Sci. 2005, 62, 1707–1723. [Google Scholar] [CrossRef]
- Nerya, O.; Ben-Arie, R.; Luzzatto, T.; Musa, R.; Khatib, S.; Vaya, J. Prevention of Agaricus bisporus postharvest browning with tyrosinase inhibitors. Postharv. Biol. Technol. 2006, 39, 272–277. [Google Scholar] [CrossRef]
- Seo, S.Y.; Sharma, V.K.; Sharma, N. Mushroom tyrosinase: recent prospects. J. Agric. Food Chem. 2003, 51, 2837–2853. [Google Scholar] [CrossRef]
- Asanuma, M.; Miyazaki, I.; Ogawa, N. Dopamine- or L-DOPA induced neurotoxicity: the role of dopamine quinone formation and tyrosinase in a model of Parkinson’s disease. Neurotox. Res. 2003, 5, 165–176. [Google Scholar] [CrossRef]
- Xu, Y.; Stokes, A.H.; Roskoski, R., Jr.; Vrana, K.E. Dopamine, in the presence of tyrosinase, covalently modifies and inactivates tyrosine hydroxylase. J. Neurosci. Res. 1998, 54, 691–697. [Google Scholar] [CrossRef]
- Matoba, Y.; Kumagai, T.; Yamamoto, A.; Yoshitsu, H.; Sugiyama, M. Crystallographic evidence that the dinuclear copper center of tyrosinase is flexible during catalysis. J. Biol. Chem. 2006, 281, 8981–8990. [Google Scholar] [CrossRef]
- Coe, F.G.; Anderson, G.J. Screening of medicinal plants used by the Garífuna of Eastern Nicaragua for bioactive compounds. J. Ethnopharmacol. 1996, 53, 29–50. [Google Scholar] [CrossRef]
- Singh, Y.N. Traditional medicine in Fiji. Some herbal folk cures used by Fiji Indians. J. Ethnopharmacol. 1986, 15, 57–88. [Google Scholar] [CrossRef]
- Sharma, L.D.; Bahga, H.S.; Srivastava, P.S. In vitro anthelmintic screening if indigenous medicinal plants against Haemonchus contortus (Rudolphi, 1803) cobbold, 1898 of sheep and goats. Indian J. Anim. Res. 1971, 5, 33–38. [Google Scholar]
- Soong, Y.Y.; Barlow, P.J. Antioxidant activity and phenolic content of selected fruit seeds. Food Chem. 2004, 88, 411–417. [Google Scholar] [CrossRef]
- Soong, Y.Y.; Barlow, P.J. Quantification of gallic acid and ellagic acid from longa (Dimocarpus longan Lour.) seed and mango (Mangifera indica L.) kernel and their effects on antioxidant activity. Food Chem. 2001, 97, 524–530. [Google Scholar] [CrossRef]
- Abdalla, A.E.M.; Darwish, S.M.; Ayad, E.H.E.; El-Hamahm, R.M. Egyptian mango by-product 1. Compositional quality of mango seed kernel. Food Chem. 2007, 103, 1134–1140. [Google Scholar] [CrossRef]
- Nithitanakool, S.; Pithayanukul, P.; Bavovada, R. Antioxidant and hepatoprotective activities of Thai mango seed kernel extract. Planta Med. (Submitted).
- Kim, J.H.; Sapers, G.M.; Choi, S.W. Isolation and identification of tyrosinase inhibitor from Galla rhois. Food Sci. Biotechnol. 1998, 7, 56–59. [Google Scholar]
- Duckworth, H.W.; Coleman, J.E. Physicochemical and kinetic properties of mushroom tyrosinase. J. Biol. Chem. 1970, 245, 1613–1625. [Google Scholar]
- Khatib, S.; Nerya, O.; Musa, R.; Tamir, S.; Peter, T.; Vaya, J. Enhanced substituted resorcinol hydrophobicity augments tyrosinase inhibition potency. J. Med. Chem. 2007, 50, 2676–2681. [Google Scholar] [CrossRef]
- Iida, K.; Hase, K.; Shimomura, K.; Sudo, S.; Kadota, S.; Namba, T. Potent inhibitors of tyrosinase activity and melanin biosynthesis from Rheum officinale. Planta Med. 1995, 161, 425–428. [Google Scholar]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; et al. Gaussian03; Gaussian, Inc: Pittsburgh, PA, USA, 2003. [Google Scholar]
- Jones, G.; Wilett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and validation of a genetic algorithm for flexible docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef]
- Sample Availability: Samples are available from the authors.
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nithitanakool, S.; Pithayanukul, P.; Bavovada, R.; Saparpakorn, P. Molecular Docking Studies and Anti-Tyrosinase Activity of Thai Mango Seed Kernel Extract. Molecules 2009, 14, 257-265. https://doi.org/10.3390/molecules14010257
Nithitanakool S, Pithayanukul P, Bavovada R, Saparpakorn P. Molecular Docking Studies and Anti-Tyrosinase Activity of Thai Mango Seed Kernel Extract. Molecules. 2009; 14(1):257-265. https://doi.org/10.3390/molecules14010257
Chicago/Turabian StyleNithitanakool, Saruth, Pimolpan Pithayanukul, Rapepol Bavovada, and Patchreenart Saparpakorn. 2009. "Molecular Docking Studies and Anti-Tyrosinase Activity of Thai Mango Seed Kernel Extract" Molecules 14, no. 1: 257-265. https://doi.org/10.3390/molecules14010257
APA StyleNithitanakool, S., Pithayanukul, P., Bavovada, R., & Saparpakorn, P. (2009). Molecular Docking Studies and Anti-Tyrosinase Activity of Thai Mango Seed Kernel Extract. Molecules, 14(1), 257-265. https://doi.org/10.3390/molecules14010257