Selenium Utilization Strategy by Microalgae

Abstract
:1. Se Requirement for Growth of Microalgae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Species | Reference |
|---|---|---|
| Diatoms | Amphiprora hyalina | [7] |
| Chaetoceros debilis | [7] | |
| Chaetoceros pelagicus | [7] | |
| Chaetoceros vixvisibilis | [7] | |
| Coscinodiscus asteromphalus | [7] | |
| Corethron criophilum | [7] | |
| Ditylum brightwellii | [7] | |
| Skeletonema costatum (strain 18c NEPCC) | [7] | |
| Skeletonema costatum (strain 611 NEPCC) | [7] | |
| Skeletonema costatum (strain 616 NEPCC) | [7] | |
| Stephanopyxis palmeriana | [7] | |
| Thalassiosira pseudonana | [4,7] | |
| Thalassiosira oceanica | [7] | |
| Thalassiosira rotula | [7] | |
| Thalassiosira aestivalis | [7] | |
| Dinoflagellates | Alexandrium minnutumb | [6] |
| Gymnodinium catenatumb | [7] | |
| Gymnodinium nagasakienseb | [9] | |
| Peridinium cinctum fa. Westii | [10] | |
| Pyrodinium bahamenseb | [11] | |
| Prymnesiophytes | Chrysochromulina breviturrita | [5] |
| Chrysochromulina kappa | [12] | |
| Chrysochromulina brevefilum | [12] | |
| Chrysochromulina strobilus | [12] | |
| Chrysochromulina polylepisb | [14,15] | |
| Helladosphaera sp. | [1] | |
| Emiliania huxleyi | [1] | |
| Gephyrocapsa oceanica | [1] | |
| Raphidophytes | Chattonella verruculosab | [8] |
| Chlorophytes | Platymonas subcordiformis | [13] |
| Chrysophytes | Aureococcus anophagefferensb | [16] |
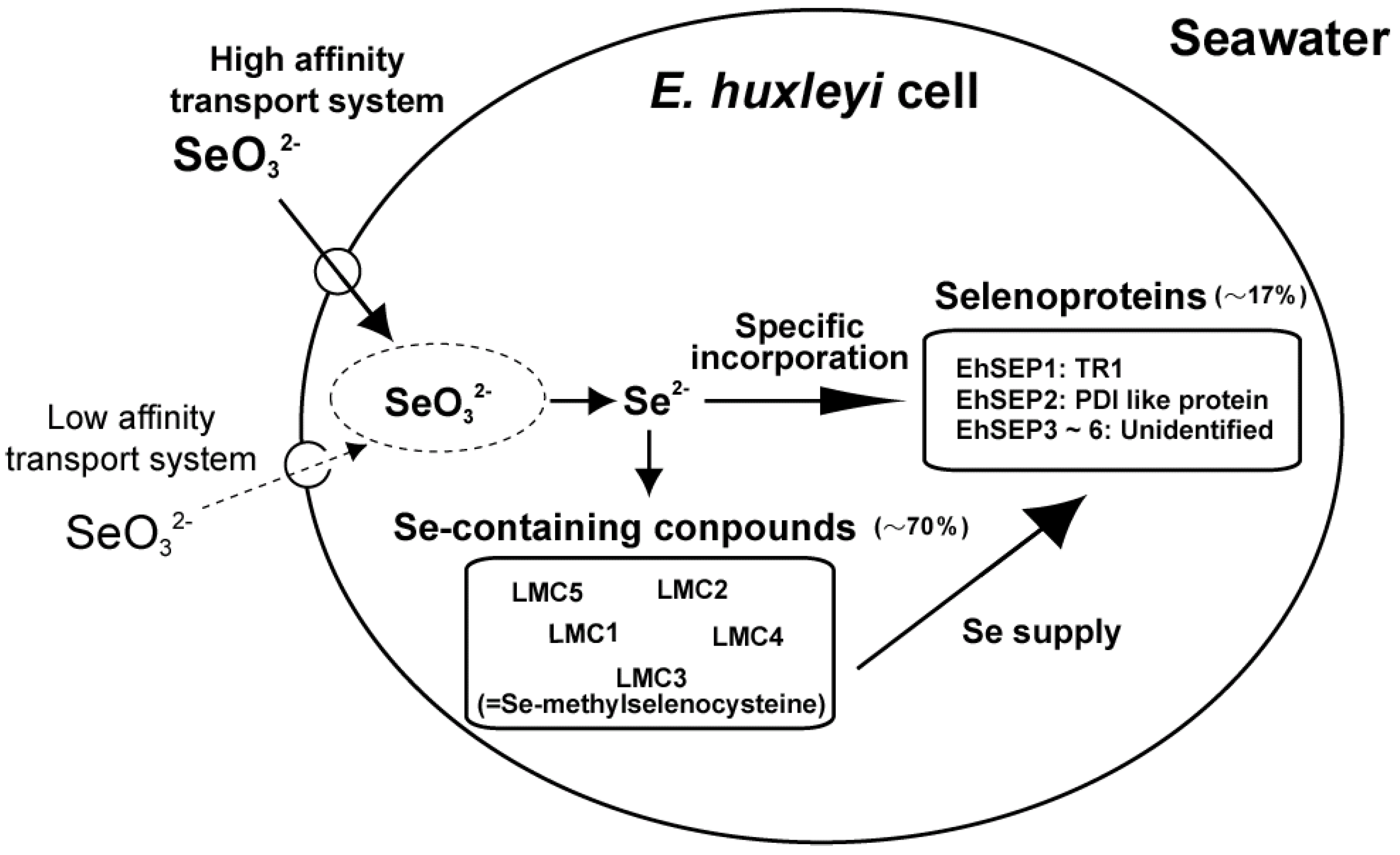
2. Se Uptake Mechanism

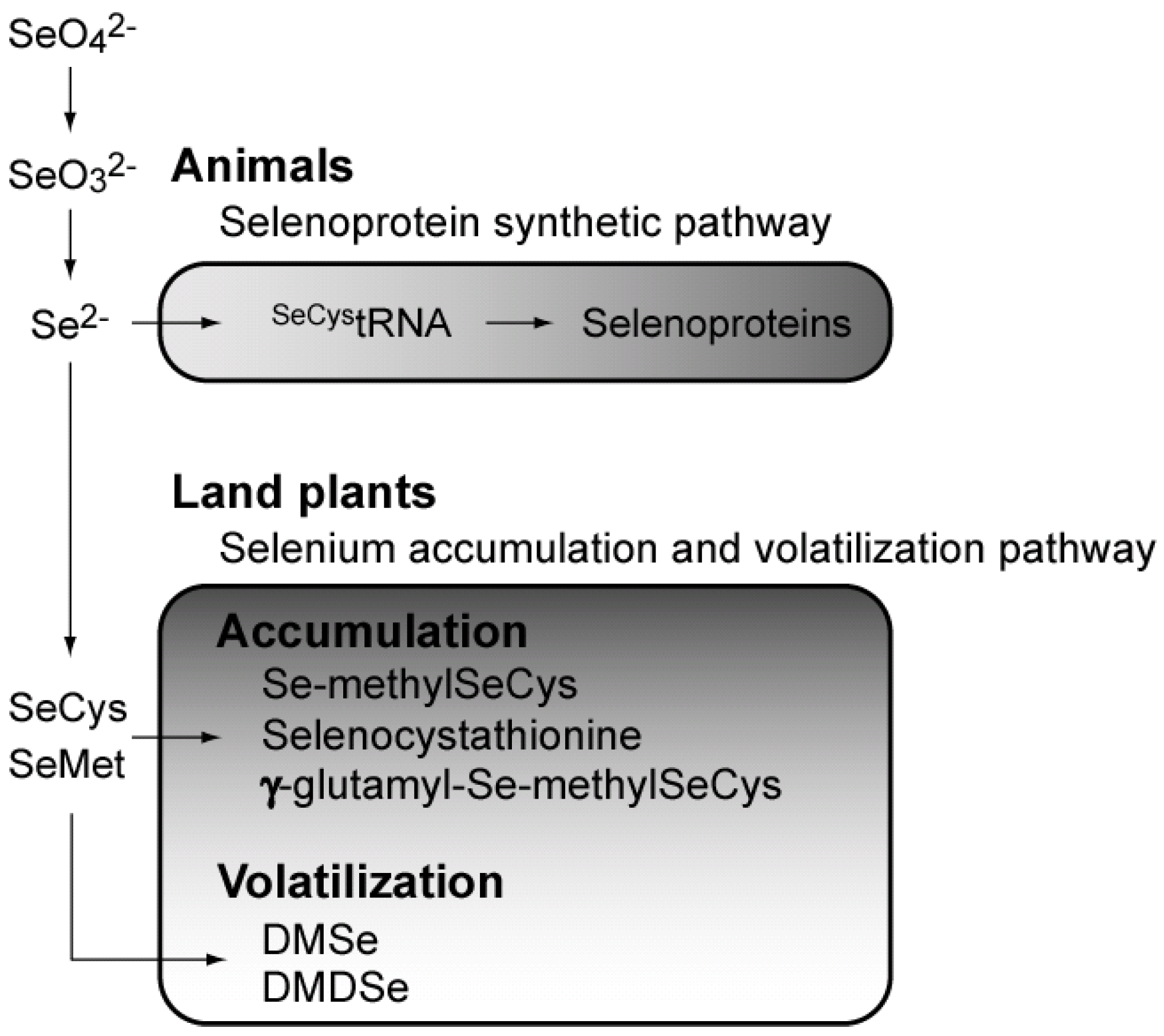
3. Metabolism of Se
| Phylum, Division | Species | Number of selenoproteins |
|---|---|---|
| Nematoda | Caenorhabditis elegans | 1 |
| Caenorhabditis briggsae | 1 | |
| Arthropoda | Apis mellifera | 1 |
| Drosophila melanogaster | 3 | |
| Drosophila pseudoobscura | 3 | |
| Anopheles gambiae | 3 | |
| Chordata | Mus musclulus | 24 |
| Homo sapiens | 25 | |
| Gallus gallus | 24 | |
| Xenopus tropicalis | 24 | |
| Ascomycota | Schizosaccharomyces pombe | 0 |
| Yarrowia lipolytica | 0 | |
| Saccharomyces cerevisiae | 0 | |
| Dictyosteliomycota | Dictyosteliumd iscoideum | 5 |
| Anthophyta | Oryza sativa | 0 |
| Medicago truncatula | 0 | |
| Populus trichocarpa | 0 | |
| Arabidopsis thaliana | 0 | |
| Chlorophyta | Chlamydomonas reinhardtii | 12 |
| Ostreococcus tauri | 26 | |
| Ostreococcus lucimarinus | 29 | |
| Rhodophyta | Cyanidioschyzon merolae | 0 |
| Heterokontophyta | Thalassiosira pseudonana | 16 |
| Apicomplexa | Cryptosporidium parvum | 0 |
| Plasmodium falciparum | 4 | |
| Plasmodium chabaudi | 4 | |
| Plasmodium yoelii | 4 |

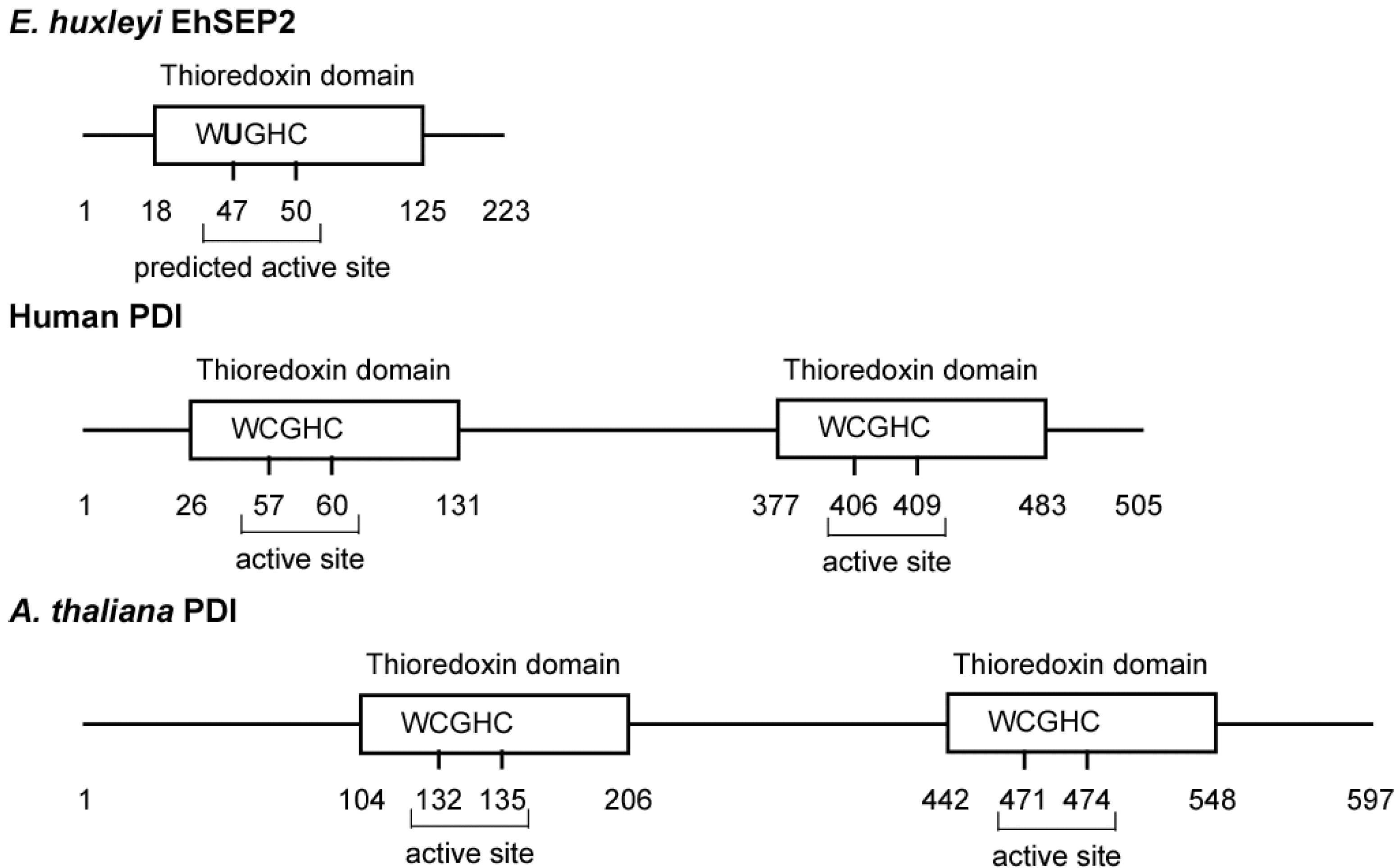
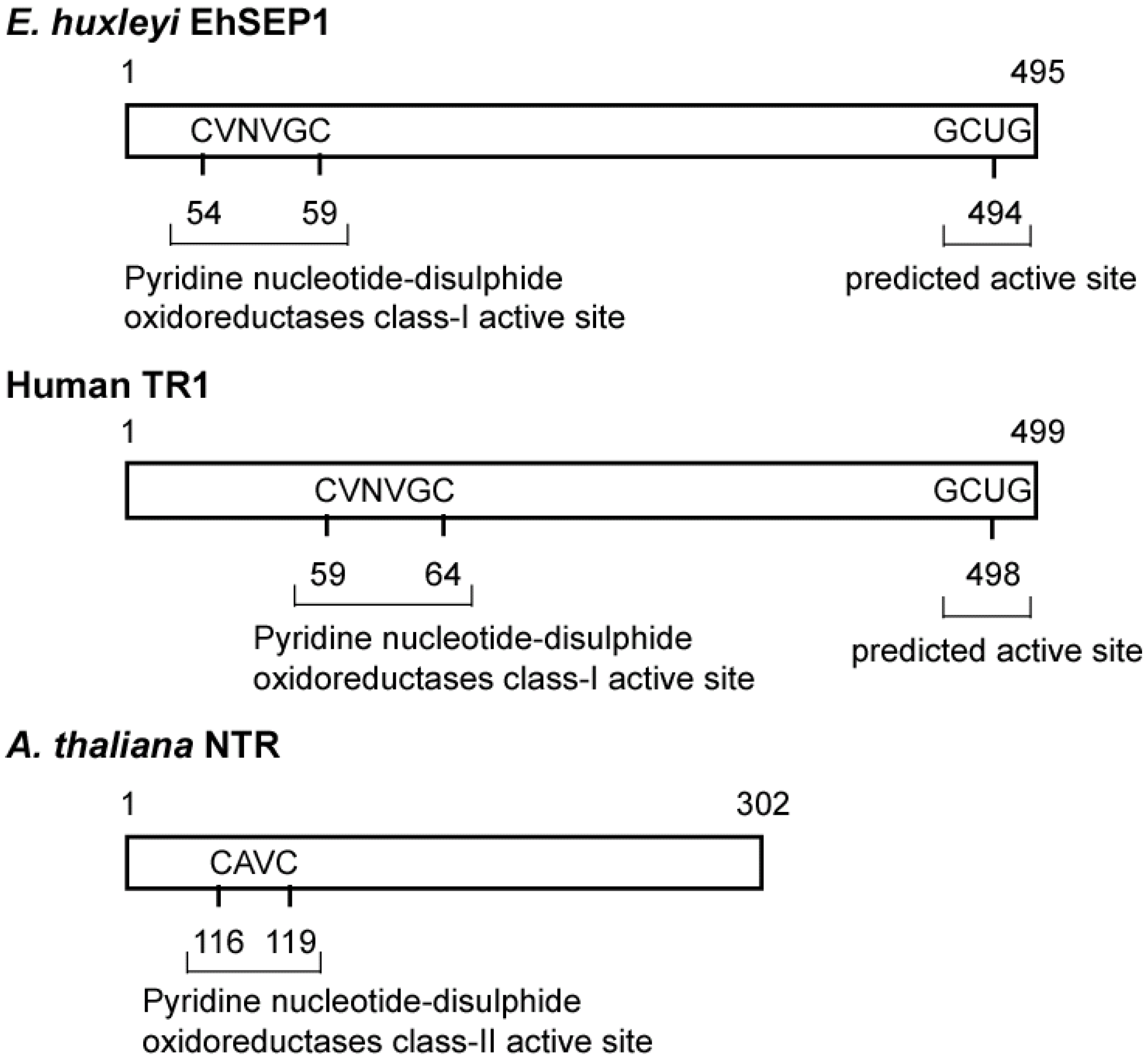
4. Selenoproteins


5. Conclusions
Acknowledgements
References and Notes
- Danbara, A.; Shiraiwa, Y. The requirement of selenium for the growth of marine coccolithophorids, Emiliania huxleyi, Gephyrocapsa oceanica and Helladosphaera sp. (Prymnesiophyceae). Plant Cell Physiol. 1999, 40, 762–766. [Google Scholar] [CrossRef]
- Cutter, G.A.; Bruland, K.W. The marine biogeochemistry of selenium: A re-evaluation. Limnol. Oceanogr. 1984, 29, 1179–1192. [Google Scholar] [CrossRef]
- Price, N.M.; Thompson, R.A.; Harrison, P.J. Selenium: An essential element for growth of the coastal marine diatom Thalassiosira pseudonana. J. Phycol. 1987, 23, 1–9. [Google Scholar] [CrossRef]
- Wehr, J.D.; Brown, L.M. Selenium requirement of a bloom-forming planktonic alga from softwater and acidified lakes. Can. J. Fish. Aquat. Sci. 1985, 42, 1783–1788. [Google Scholar] [CrossRef]
- Doblin, M.A.; Blackburn, S.I.; Hallegraeff, G.M. Comparative study of selenium requirements of three phytoplankton species: Gymnodinium catenatum, Alexandrium minutum (Dinophyta) and Chaetoceros cf. tenuissimus. J. Plankton Res. 1999, 21, 1153–1169. [Google Scholar] [CrossRef]
- Doblin, M.A.; Blackburn, S.I.; Hallegraeff, G.M. Intraspecific variation in the selenium requirement of different geographic strains of the toxic dinoflagellate Gymnodinium catenatum. J. Plankton Res. 2000, 22, 421–432. [Google Scholar] [CrossRef]
- Harrison, P.J.; Yu, P.W.; Thompson, P.A.; Price, N.M.; Phillips, D.J. Survey of selenium requirements in marine phytoplankton. Mar. Ecol. Prog. Ser. 1988, 47, 89–96. [Google Scholar] [CrossRef]
- Imai, I.; Itakura, S.; Matsuyama, Y.; Yamaguchi, M. Selenium requirement for growth of a novel red tide flagellate Chattonella verruculosa (Raphidophyceae) in culture. Fish. Sci. 1996, 62, 834–835. [Google Scholar]
- Ishimaru, T.; Takeuchi, T.; Fukuyo, Y.; Kodama, M. The selenium requirement of Gymnodinium nagasakiense. In Red Tides: Biology, Environmental Science and Toxicology; Okaichi, T., Anderson, D.M., Nemoto, T., Eds.; Elsevier: New York, NY, USA, 1989; pp. 357–360. [Google Scholar]
- Lindström, K.; Rodhe, W. Selenium as a micronutrient for the dinoflagellate Peridinium cinctum fa. westii. Mitt. Int. Ver. Theor. Angew. Limnol. 1978, 21, 168–173. [Google Scholar]
- Usup, G.; Azanza, R.V. Physiology and bloom dynamics of the tropical dinoflagellate Pyrodinium bahamense (Dinophyceae). In Physiological Ecology of Harmful Algal Blooms; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer-Verlag: Heidelberg, Germany, 1998; pp. 81–94. [Google Scholar]
- Pintner, I.; Provasoli, L. Heterotrophy in subdued light of three Chrysochromulina species. Bull. Misaki Mar. Biol. Inst. Kyoto Univ. 1968, 12, 25–31. [Google Scholar]
- Wheeler, A.E.; Zingaro, R.A.; Irgolic, K.; Bottino, N.R. The effect of selenate, selenite and sulphate on the growth of six unicellular green algae. J. Exp. Mar. Biol. Ecol. 1982, 57, 181–194. [Google Scholar] [CrossRef]
- Dahl, E.; Lindahl, O.; Paasche, E.; Throndsen, J. The Chrysochromulina polylepsis bloom in Scandinavian waters during spring 1988. In Novel Phytoplankton Blooms; Cosper, E.M., Bricelj, V.M., Carpenter, E.J., Eds.; Springer-Verlag: Berlin, Germany, 1989; pp. 383–403. [Google Scholar]
- Edvardsen, E.; Moy, F.; Paasche, E. Hemolytic activity in extracts of Chrysochromulina polylepsis grown at different levels of selenite and phosphate. In Toxic Marine Phytoplankton; Granéli, E., Sundström, B., Edler, L., Anderson, D.M., Eds.; Elsevier: New York, NY, USA, 1990; pp. 284–289. [Google Scholar]
- Cosper, E.M.; Garry, R.T.; Milligan, A.J.; Doall, M.H. Iron, selenium and citric acid are critical to the growth of the brown tide microalga, Aureococcus Anophagefferens. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimazu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 667–674. [Google Scholar]
- Hu, M.; Yang, Y.; Martin, J.-M.; Yin, K.; Harrison, P.J. Preferential uptake of Se(IV) over Se(VI) and the production of dissolved organic Se by marine phytoplankton. Mar. Environ. Res. 1996, 44, 225–231. [Google Scholar]
- Arvy, M. P. Selenate and selenite uptake and translocation in bean plants (Phaseolus vulgaris). J. Exp. Bot. 1993, 44, 1083–1087. [Google Scholar] [CrossRef]
- Morlon, H.; Fortin, C.; Adam, C.; Garnier-Laplace, J. Selenite transport and its inhibition in the unicellular green alga Chlamydomonas reinhardtii. Environ. Toxicol. Chem. 2006, 25, 1408–1417. [Google Scholar] [CrossRef]
- Li, H.F.; McGrath, S.P.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytologist 2008, 178, 92–102. [Google Scholar]
- Obata, T.; Araie, H.; Shiraiwa, Y. Bioconcentration mechanism of selenium by a coccolithophorid. Emiliania huxleyi. Plant Cell Physiol. 2004, 45, 1434–1441. [Google Scholar] [CrossRef]
- Pickering, I.J.; Wright, C.; Bubner, B.; Ellis, D.; Persans, M.W.; Yu, E.Y.; George, G.N.; Prince, R.C.; Salt, D.E. Chemical form and distribution of selenium and sulfur in the selenium hyperaccumulator Astragalus bisulcatus. Plant Physiol. 2003, 131, 1460–1467. [Google Scholar]
- Sors, T.G.; Ellis, D.R.; Salt, D.E. Selenium uptake, translocation, assimilation and metabolic fate in plants. Photosynth. Res. 2005, 86, 373–389. [Google Scholar] [CrossRef]
- Obata, T.; Sera, K.; Futatsugawa, S.; Shiraiwa, Y. Multi-element analysis of marine microalgae using particle-induced X-ray emission (PIXE). Mar. Biotechnol. 2004, 6, S66–S70. [Google Scholar]
- Brown, T.A.; Shrift, A. Exclusion of selenium from proteins in selenium-tolerant Astragalus species. Plant Physiol. 1981, 67, 1951–1953. [Google Scholar]
- Brown, K.M.; Arthur, J.R. Selenium, selenoproteins and human health: A review. Public HealthNutr. 2001, 4, 593–599. [Google Scholar]
- Kim, H.-Y.; Fomenko, D.E.; Yoon, Y.-E.; Gladyshev, V.N. Catalytic advantages provided by selenocysteine in methionine-S-sulfoxide reductases. Biochemistry 2006, 45, 13697–13704. [Google Scholar]
- Stadtman, T.C. Selenocysteine. Annu. Rev. Biochem. 1996, 65, 83–100. [Google Scholar] [CrossRef]
- Hatfield, D.L.; Gladyshev, V.N. How selenium has altered our understanding of the genetic code. Mol Cell Biol. 2002, 22, 3565–3576. [Google Scholar] [CrossRef]
- Novoselov, S.V.; Rao, M.; Onoshko, N.V.; Zhi, H.; Kryukov, G.V.; Xiang, Y.; Weeks, D.P.; Hatfield, D.L.; Gladyshev, V.N. Selenoprotein and selenocysteine insertion system in the model plant cell system, Chlamydomonas reinhardtii. EMBO J. 2002, 21, 3681–3693. [Google Scholar] [CrossRef]
- Price, N.M.; Harrison, P.J. Specific selenium-containing macromolecules in the marine diatom Thalassiosira pseudonana. Plant Physiol. 1988, 86, 192–199. [Google Scholar] [CrossRef]
- Obata, T.; Shiraiwa, Y. A novel eukaryotic selenoprotein in the haptophyte alga Emiliania huxleyi. J. Biol. Chem. 2005, 280, 18462–18468. [Google Scholar] [CrossRef]
- Araie, H.; Suzuki, I.; Shiraiwa, Y. Identification and characterization of a selenoprotein, thioredoxin reductase, in a unicellular marine haptophyte alga, Emiliania huxleyi. J. Biol. Chem. 2008, 283, 35329–35336. [Google Scholar] [CrossRef]
- Lobanov, A.V.; Fomenko, D.E.; Zhang, Y.; Sengupta, A.; Hatfield, D.L.; Gladyshev, V.N. Evolutionary dynamics of eukaryotic selenoproteomes: large selenoproteomes may associate with aquatic life and small with terrestrial life. Genome Biol. 2007, 8, R198. [Google Scholar] [CrossRef]
- Ferrari, D.M.; Söling, H.D. The protein disulphide-isomerase family: unravelling a string of folds. Biochem. J. 1999, 339, 1–10. [Google Scholar] [CrossRef]
- Umysová, D.; Vítová, M.; Doušková, I.; Bišová, K.; Hlavová, M.; Čížková, M.; Machát, J.; Doucha, J.; Zachleder, V. Bioaccumulation of selenium compounds and thioredoxin reductase activity in the green alga Scenedesmus quadricauda. BMC Plant Biol. 2009, 9, 58. [Google Scholar] [CrossRef]
- Jakupoglu, C.; Przemeck, G.K.; Schneider, M.; Moreno, S.G.; Mayr, N.; Hatzopoulos, A.K.; de Angelis, M.H.; Wurst, W.; Bornkamm, G.W.; Brielmeier, M.; Conrad, M. Cytoplasmic Thioredoxin reductase is essential for embryogenesis but dispensable for cardiac development. Mol. Cell. Biol. 2005, 25, 1980–1988. [Google Scholar] [CrossRef]
- Marchetti, A.; Parker, M.S.; Moccia, L.P.; Lin, E.O.; Arrieta, A.L.; Ribalet, F.; Murphy, M.E.P; Maldonado, M.T.; Armbrust, E.V. Ferritin is used for iron storage in bloom-forming marine pennate diatoms. Nature 2009, 457, 467–470. [Google Scholar]
- Sample Availability: Not available.
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Araie, H.; Shiraiwa, Y. Selenium Utilization Strategy by Microalgae. Molecules 2009, 14, 4880-4891. https://doi.org/10.3390/molecules14124880
Araie H, Shiraiwa Y. Selenium Utilization Strategy by Microalgae. Molecules. 2009; 14(12):4880-4891. https://doi.org/10.3390/molecules14124880
Chicago/Turabian StyleAraie, Hiroya, and Yoshihiro Shiraiwa. 2009. "Selenium Utilization Strategy by Microalgae" Molecules 14, no. 12: 4880-4891. https://doi.org/10.3390/molecules14124880
APA StyleAraie, H., & Shiraiwa, Y. (2009). Selenium Utilization Strategy by Microalgae. Molecules, 14(12), 4880-4891. https://doi.org/10.3390/molecules14124880