Development of a High-Throughput Assay for Screening of γ-Secretase Inhibitor with Endogenous Human, Mouse or Drosophila γ-Secretase

Abstract
:1. Introduction
2. Results and Discussion
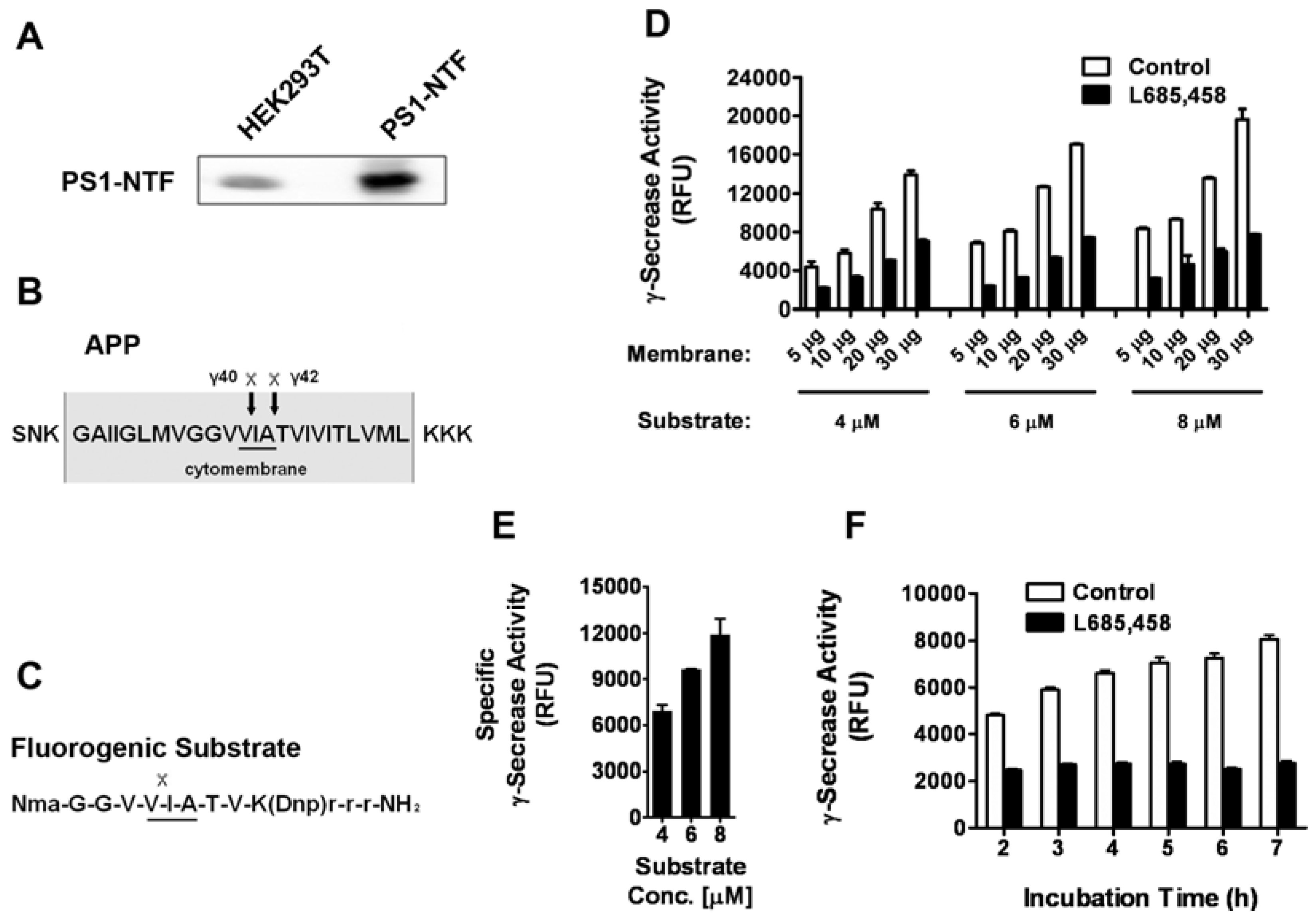
2.1. Assay optimization
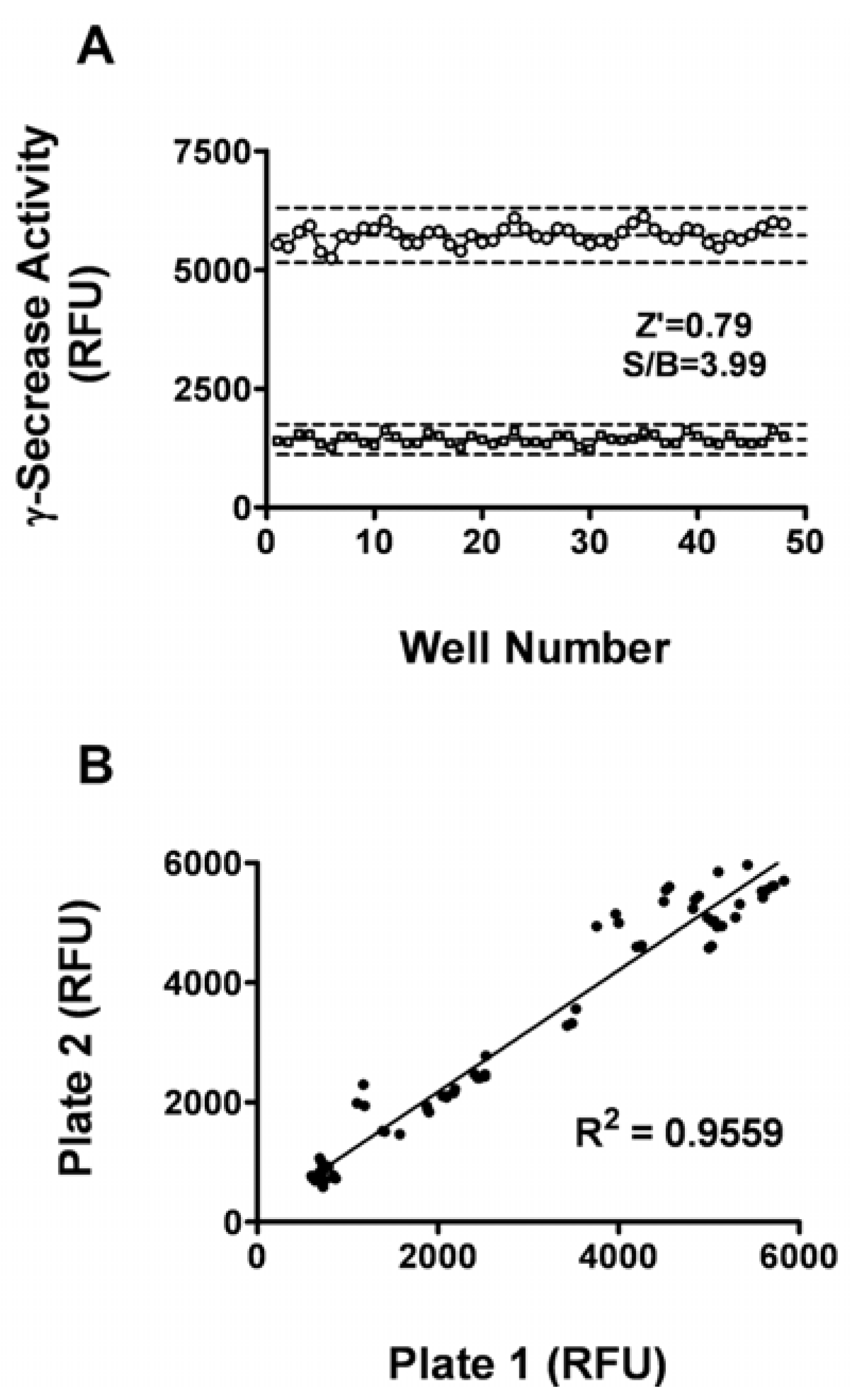
2.2. Assay performance
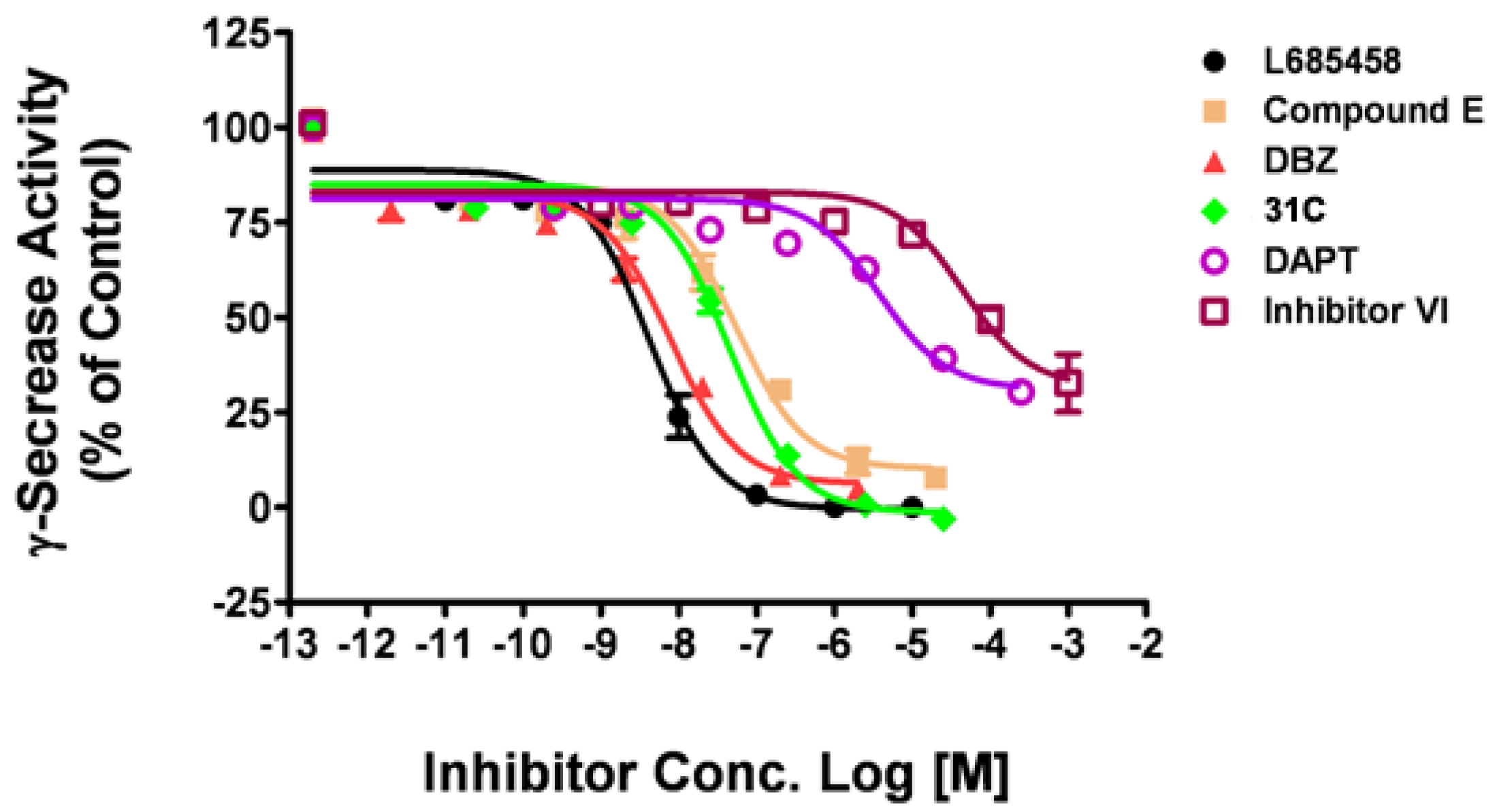
2.3. The γ-secretase inhibitor activity measurement with human γ-secretase
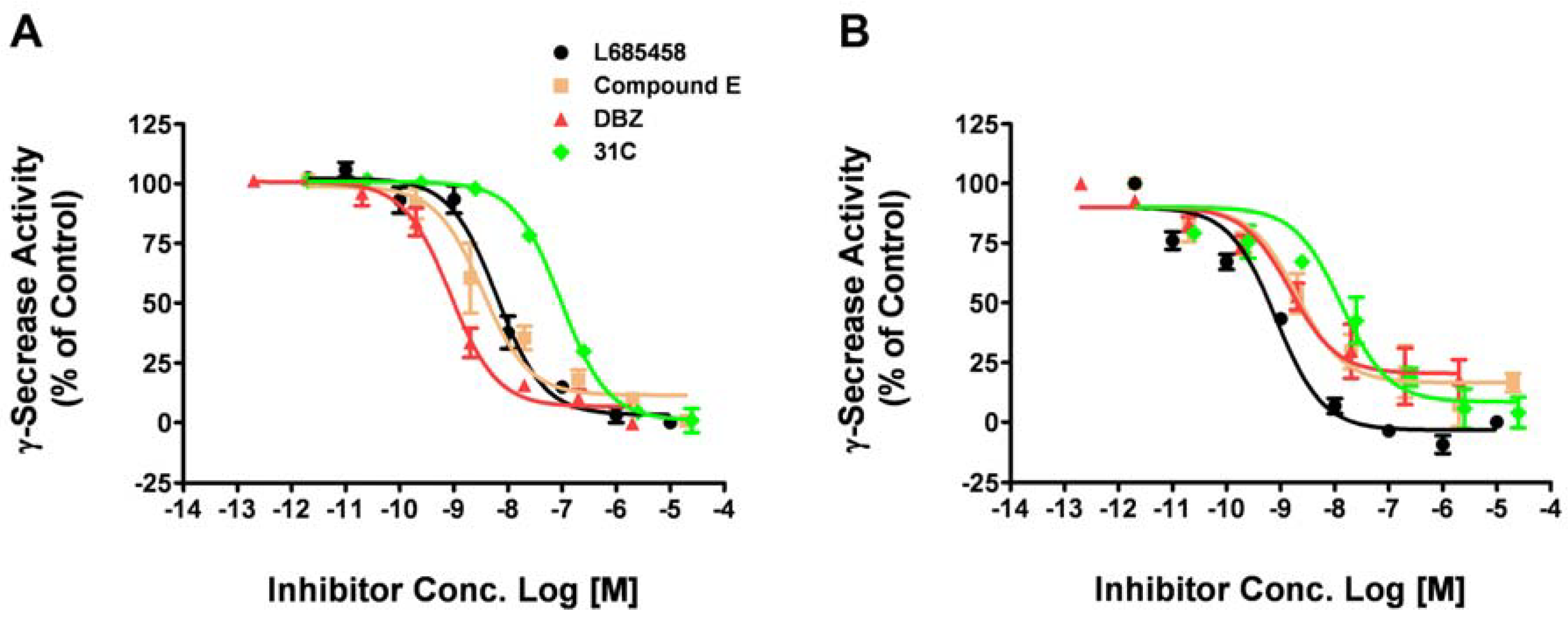
2.4. The γ-secretase inhibitor activity measurement with drosophila and mouse γ-secretase
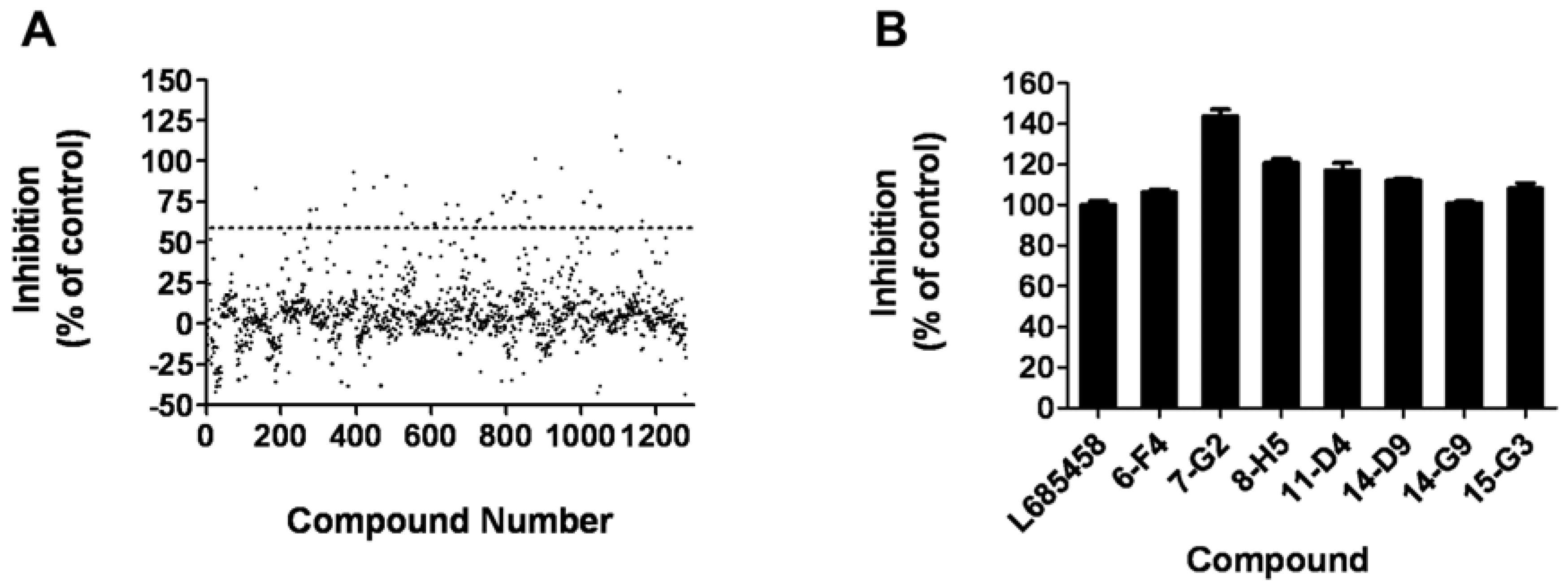
2.5. HTS campaign
3. Experimental
3.1. Reagents
3.2. Cell culture
3.3. Animals
3.4. Membrane protein preparation from cells and mouse brain
3.5. Fluorogenic substrate assay
3.6. Western blot
3.7. Data analysis
4. Conclusions
Acknowledgements
References
- Tanzi, R.E. The synaptic Aβ hypothesis of Alzheimer disease. Nat. Neurosci. 2005, 8, 977–979. [Google Scholar] [CrossRef] [PubMed]
- Iwatsubo, T.; Odaka, A.; Suzuki, N.; Mizusawa, H.; Ihara, Y. Visualization of Aβ42(43) and Aβ40 in senile plaques with end-specific Aβ monoclonals: Evidence that an initially deposited species is Aβ42(43). Neuron 1994, 13, 45–53. [Google Scholar] [CrossRef]
- Marlow, L.; Canet, R.M.; Haugabook, S.J.; Hardy, J.A.; Lahiri, D.K.; Sambamurti, K. APH1, PEN2, and Nicastrin increase Abeta levels and gamma-secretase activity. Biochem. Biophys. Res. Commun. 2003, 305, 502–509. [Google Scholar] [CrossRef]
- Li, Y.M.; Lai, M.T.; Xu, M.; Huang, Q.; DiMuzio-Mower, J.; Sardana, M.K.; Shi, X.P.; Yin, K.C.; Shafer, J.A.; Gardell, S.J. Presenilin 1 is linked with gamma-secretase activity in the detergent solubilized state. Proc. Natl. Acad. Sci. USA 2000, 97, 6138–6143. [Google Scholar] [CrossRef] [PubMed]
- Dovey, H.F.; John, V.; Anderson, J.P.; Chen, L.Z.; De, S.A.P.; Fang, L.Y.; Freedman, S.B.; Folmer, B.; Goldbach, E.; Holsztynska, E.J.; Hu, K.L.; Johnson-Wood, K.L.; Kennedy, S.L.; Kholodenko, D.; Knops, J.E.; Latimer, L.H.; Lee, M.; Liao, Z.; Lieberburg, I.M.; Motter, R.N.; Mutter, L.C.; Nietz, J.; Quinn, K.P.; Sacchi, K.L.; Seubert, P.A.; Shopp, G.M.; Thorsett, E.D.; Tung, J.S.; Wu, J.; Yang, S.; Yin, C.T.; Schenk, D.B.; May, P.C.; Altstiel, L.D.; Bender, M.H.; Boggs, L.N.; Britton, T.C.; Clemens, J.C.; Czilli, D.L.; Dieckman-McGinty, D.K.; Droste, J.J.; Fuson, K.S.; Gitter, B.D.; Hyslop, P.A.; Johnstone, E.M.; Li, W.Y.; Little, S.P.; Mabry, T.E.; Miller, F.D.; Audia, J.E. Functional gamma-secretase inhibitors reduce beta-amyloid peptide levels in brain. J. Neurochem. 2001, 173–181. [Google Scholar]
- Shearman, M.S.; Beher, D.; Clarke, E.E.; Lewis, H.D.; Harrison, T.; Hunt, P.; Nadin, A.; Smith, A.L.; Stevenson, G.; Castro, J.L. L-685,458, an aspartyl protease transition state mimic, is a potent inhibitor of amyloid beta-protein precursor gamma-secretase activity. Biochemistry 2000, 39, 8698–8704. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, M.S. Inhibition and modulation of gamma-secretase for Alzheimer's disease. Neurotherapeutics 2008, 5, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Rafii, M.S.; Aisen, P.S. Recent developments in Alzheimer's disease therapeutics. BMC Med. 2009, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Beher, D.; Wrigley, J.D.; Nadin, A.; Evin, G.; Masters, C.L.; Harrison, T.; Castro, J.L.; Shearman, M.S. Pharmacological knock-down of the presenilin 1 heterodimer by a novel gamma -secretase inhibitor: Implications for presenilin biology. J. Biol. Chem. 2001, 276, 45394–45402. [Google Scholar] [CrossRef] [PubMed]
- Vandermeeren, M.; Geraerts, M.; Pype, S.; Dillen, L.; Van Hove, C.; Mercken, M. The functional gamma-secretase inhibitor prevents production of amyloid beta 1-34 in human and murine cell lines. Neurosci. Lett. 2001, 315, 145–148. [Google Scholar] [CrossRef]
- Berezovska, O.; Jack, C.; McLean, P.; Aster, J.C.; Hicks, C.; Xia, W.; Wolfe, M.S.; Kimberly, W.T.; Weinmaster, G.; Selkoe, D.J.; Hyman, B.T. Aspartate mutations in presenilin and gamma-secretase inhibitors both impair notch1 proteolysis and nuclear translocation with relative preservation of notch1 signaling. J. Neurochem. 2000, 75, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Farmery, M.R.; Tjernberg, L.O.; Pursglove, S.E.; Bergman, A.; Winblad, B.; Naslund, J. Partial purification and characterization of gamma-secretase from post-mortem human brain. J. Biol. Chem. 2003, 278, 24277–24284. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Lai, M.T.; Huang, Q.; DiMuzio-Mower, J.; Castro, J.L.; Harrison, T.; Nadin, A.; Neduvelil, J.G.; Shearman, M.S.; Shafer, J.A.; Gardell, S.J.; Li, Y.M. gamma-Secretase: Characterization and implication for Alzheimer disease therapy. Neurobiol. Aging 2002, 23, 1023–1030. [Google Scholar] [CrossRef]
- Zhang, J.H.; Chung, T.D.Y.; Oldenburg, K.R. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xie, X. Development of a quantitative, cell-based, high-content screening assay for epidermal growth factor receptor modulators. Acta Pharmacol. Sin. 2007, 28, 1698–1704. [Google Scholar] [CrossRef] [PubMed]
- Xu, X. gamma-Secretase catalyzes sequential cleavages of the AbetaPP transmembrane domain. J. Alzheimers Dis. 2009, 16, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Morohashi, Y.; Kan, T.; Tominari, Y.; Fuwa, H.; Okamura, Y.; Watanabe, N.; Sato, C.; Natsugari, H.; Fukuyama, T.; Iwatsubo, T.; Tomita, T. C-terminal fragment of presenilin is the molecular target of a dipeptidic gamma-secretase-specific inhibitor DAPT (N-[N-(3,5-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester). J. Biol. Chem. 2006, 281, 14670–14676. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, H.; Sai, X.; Wang, H.Q.; Maeda, Y.; Kurono, Y.; Nishimura, M.; Yanagisawa, K.; Komano, H. PEN-2 enhances gamma-cleavage after presenilin heterodimer formation. J. Neurochem. 2004, 90, 1402–1413. [Google Scholar] [CrossRef] [PubMed]
- Milano, J.; McKay, J.; Dagenais, C.; Foster-Brown, L.; Pognan, F.; Gadient, R.; Jacobs, R.T.; Zacco, A.; Greenberg, B.; Ciaccio, P.J. Modulation of notch processing by gamma-secretase inhibitors causes intestinal goblet cell metaplasia and induction of genes known to specify gut secretory lineage differentiation. Toxicol. Sci. 2004, 82, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Esler, W.P.; Kimberly, W.T.; Ostaszewski, B.L.; Ye, W.; Diehl, T.S.; Selkoe, D.J.; Wolfe, M.S. Activity-dependent isolation of the presenilin- gamma -secretase complex reveals nicastrin and a gamma substrate. Proc. Natl. Acad. Sci. USA 2002, 99, 2720–2725. [Google Scholar] [CrossRef] [PubMed]
- Rishton, G.M.; Retz, D.M.; Tempest, P.A.; Novotny, J.; Kahn, S.; Treanor, J.J.; Lile, J.D.; Citron, M. Fenchylamine sulfonamide inhibitors of amyloid beta peptide production by the gamma-secretase proteolytic pathway: Potential small-molecule therapeutic agents for the treatment of Alzheimer's disease. J. Med. Chem. 2000, 43, 2297–2299. [Google Scholar] [CrossRef] [PubMed]
- Takasugi, N.; Takahashi, Y.; Morohashi, Y.; Tomita, T.; Iwatsubo, T. The mechanism of gamma-secretase activities through high molecular weight complex formation of presenilins is conserved in Drosophila melanogaster and mammals. J. Biol. Chem. 2002, 277, 50198–50205. [Google Scholar] [CrossRef] [PubMed]
- Micchelli, C.A.; Esler, W.P.; Kimberly, W.T.; Jack, C.; Berezovska, O.; Kornilova, A.; Hyman, B.T.; Perrimon, N.; Wolfe, M.S. Gamma-secretase/presenilin inhibitors for Alzheimer's disease phenocopy Notch mutations in Drosophila. FASEB J. 2003, 17, 79–81. [Google Scholar] [CrossRef] [PubMed]
- Best, J.D.; Smith, D.W.; Reilly, M.A.; O'Donnell, R.; Lewis, H.D.; Ellis, S.; Wilkie, N.; Rosahl, T.W.; Laroque, P.A.; Boussiquet-Leroux, C.; Churcher, I.; Atack, J.R.; Harrison, T.; Shearman, M.S. The novel gamma secretase inhibitor N-[cis-4-[(4-chlorophenyl)sulfonyl]-4-(2,5-difluorophenyl)cyclohexyl]-1,1, 1-trifluoromethanesulfonamide (MRK-560) reduces amyloid plaque deposition without evidence of notch-related pathology in the Tg2576 mouse. J. Pharmacol. Exp. Ther. 2007, 320, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Hyde, L.A.; McHugh, N.A.; Chen, J.; Zhang, Q.; Manfra, D.; Nomeir, A.A.; Josien, H.; Bara, T.; Clader, J.W.; Zhang, L.; Parker, E.M.; Higgins, G.A. Studies to investigate the in vivo therapeutic window of the gamma-secretase inhibitor N2-[(2S)-2-(3,5-difluorophenyl)-2-hydroxyethanoyl]-N1-[(7S)-5-methyl-6-oxo-6,7-dihydro-5H-dibenzo[b,d]azepin-7-yl]-L-alaninamide (LY411,575) in the CRND8 mouse. J. Pharmacol. Exp. Ther. 2006, 319, 1133–1143. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Fluorogenic substrate assay IC50 (nM) | Reported IC50(nM) / Assay used |
|---|---|---|
| L685,458 | 4.43 ± 0.9691 | IC50 (Aβ40) = 4 nM; IC50(Aβ42) = 8 nM /immunoblotting assay [9] IC50 = 17 ± 8 nM/ HTRF assay [6] IC50 = 3 nM / in vitro γ-secretase assay [17] |
| Compound E | 53.86 ± 5.312 | IC50(Aβ40) = 0.24 nM; IC50(Aβ42) = 0.37 nM / immunoblotting assay[9] IC50(Aβ40) = 0.8 nM; IC50(Aβ42) = 7 nM / Aβ ELISA assay [18] IC50 = 7 nM / in vitro γ-secretase assay [17] |
| DBZ | 12.50 ± 5.764 | IC50 = 1.7 nM / Sup-T1 NICT assay[19] |
| DAPT | 3910 ± 550.7 | IC50(Aβ40) = 1600 nM IC50(Aβ42) = 4000 nM / Aβ ELISA assay [18] |
| 31C | 53.48 ± 13.58 | IC50((Aβ) = 300 nM / In vivo γ-secretase assay [20] |
| Inhibitor VI | 44457 ± 25568 | IC50 (Aβ1-42) = 1800 nM [21] |
| Compound | human γ-secretase IC50 (nM) | drosophila γ-secretase IC50 (nM) | mouse γ-secretase IC50 (nM) |
|---|---|---|---|
| L685,458 | 4.43 ± 0.9691 | 10.35 ± 1.657 | 0.9759 ± 0.1467 |
| Compound E | 53.86 ± 5.312 | 40.55 ± 6.997 | 2.398 ± 0.6046 |
| DBZ | 12.50 ± 5.764 | 6.970 ± 1.984 | 1.446 ± 0.4350 |
| 31C | 53.48 ± 13.58 | 0.9328 ± 0.1690 | 20.69 ± 5.242 |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, L.-F.; Zhang, R.; Xie, X. Development of a High-Throughput Assay for Screening of γ-Secretase Inhibitor with Endogenous Human, Mouse or Drosophila γ-Secretase. Molecules 2009, 14, 3589-3599. https://doi.org/10.3390/molecules14093589
Wang L-F, Zhang R, Xie X. Development of a High-Throughput Assay for Screening of γ-Secretase Inhibitor with Endogenous Human, Mouse or Drosophila γ-Secretase. Molecules. 2009; 14(9):3589-3599. https://doi.org/10.3390/molecules14093589
Chicago/Turabian StyleWang, Lie-Feng, Ru Zhang, and Xin Xie. 2009. "Development of a High-Throughput Assay for Screening of γ-Secretase Inhibitor with Endogenous Human, Mouse or Drosophila γ-Secretase" Molecules 14, no. 9: 3589-3599. https://doi.org/10.3390/molecules14093589
APA StyleWang, L. -F., Zhang, R., & Xie, X. (2009). Development of a High-Throughput Assay for Screening of γ-Secretase Inhibitor with Endogenous Human, Mouse or Drosophila γ-Secretase. Molecules, 14(9), 3589-3599. https://doi.org/10.3390/molecules14093589