Anti-Inflammatory and Free Radial Scavenging Activities of the Constituents Isolated from Machilus zuihoensis

Abstract
:1. Introduction
2. Results and Discussion

2.1. Effects of the Constituents of Leaves of M. zuihoensis on Superoxide Anion Radical (O2•−) Scavenging Activities

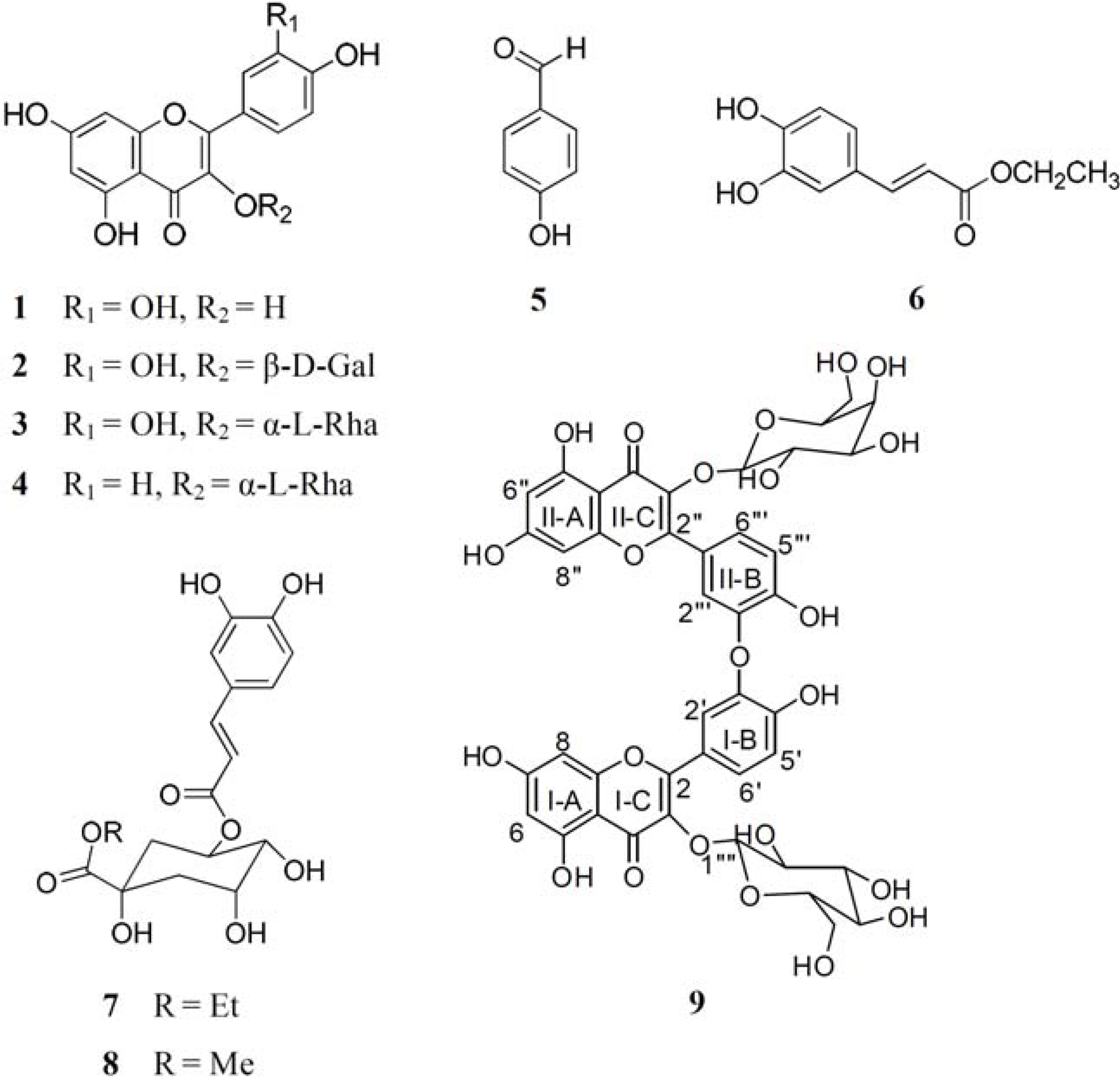{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | IC50 (μM) |
|---|---|
| (+) Catechin (positive control) | 41.6 |
| Quercetin (1) | 42.6 |
| Hyperoside (2) | >100 |
| Quercitrin (3) | 75.1 |
| Afzelin (4) | >100 |
| 4-Hydroxybenzaldehyde (5) | >100 |
| Ethyl caffeate (6) | 66.7 |
| Chlorogenic acid ethyl ester (7) | >100 |
| Clorogenic acid methyl ester (8) | 99.0 |
| Quercetin-3-O-β-D-glucopyranoside-(3′→O-3‴)-quercetin-3-O-β-D-galactopyranoside (9) | 30.4 |
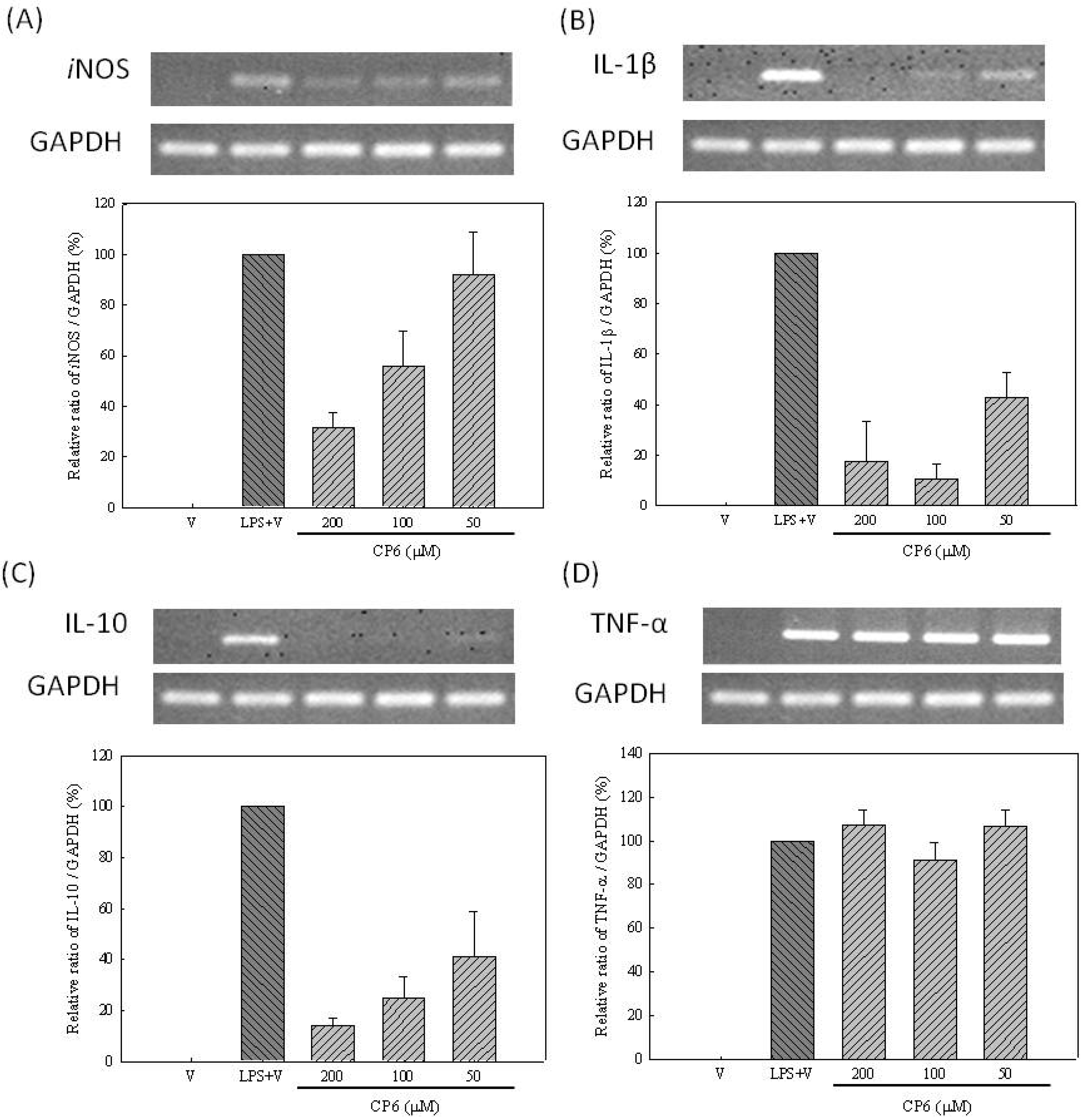
2.2. Effects of Constituents of Leaves of M. zuihoensis on Anti-Inflammatory Activities



| Samples | iNOS | IL-1β | TNF-α | IL-10 |
|---|---|---|---|---|
| Ethyl caffeate (6) | ↓ | ↓ | ─ a | ↓ |
| SB203580 (p38 inhibitor, 10 μM) | ─ a | ↓ | ─ a | ↓ |
| Wortmannin (PI3K/Akt inhibitor, 1 μM) | ─ a | ─ a | ─ a | ─ a |
| SP600125 (JNK inhibitor, 10 μM) | ↓ | ↓ | ↓ | ↓ |
| U0126 (ERK inhibitor, 1 μM) | ─ a | ↓ | ↓ | ↓ |
| PDTC (NF-κB inhibitor, 100 μM) | ↓ | ↓ | ─ a | ↓ |
3. Experimental
3.1. General
3.2. Plant Materials
3.3. Extraction and Isolation
3.4. Spectral Data of the New Compound Quercetin-3-O-β-D-glucopyranoside-(3′→O-3‴)-quercetin-3-O-β-D-galactopyranoside (9)
3.4.1. Acid Hydrolysis of Compound 9
3.4.2. Structural Determination of Compound 9
3.5. Reagents
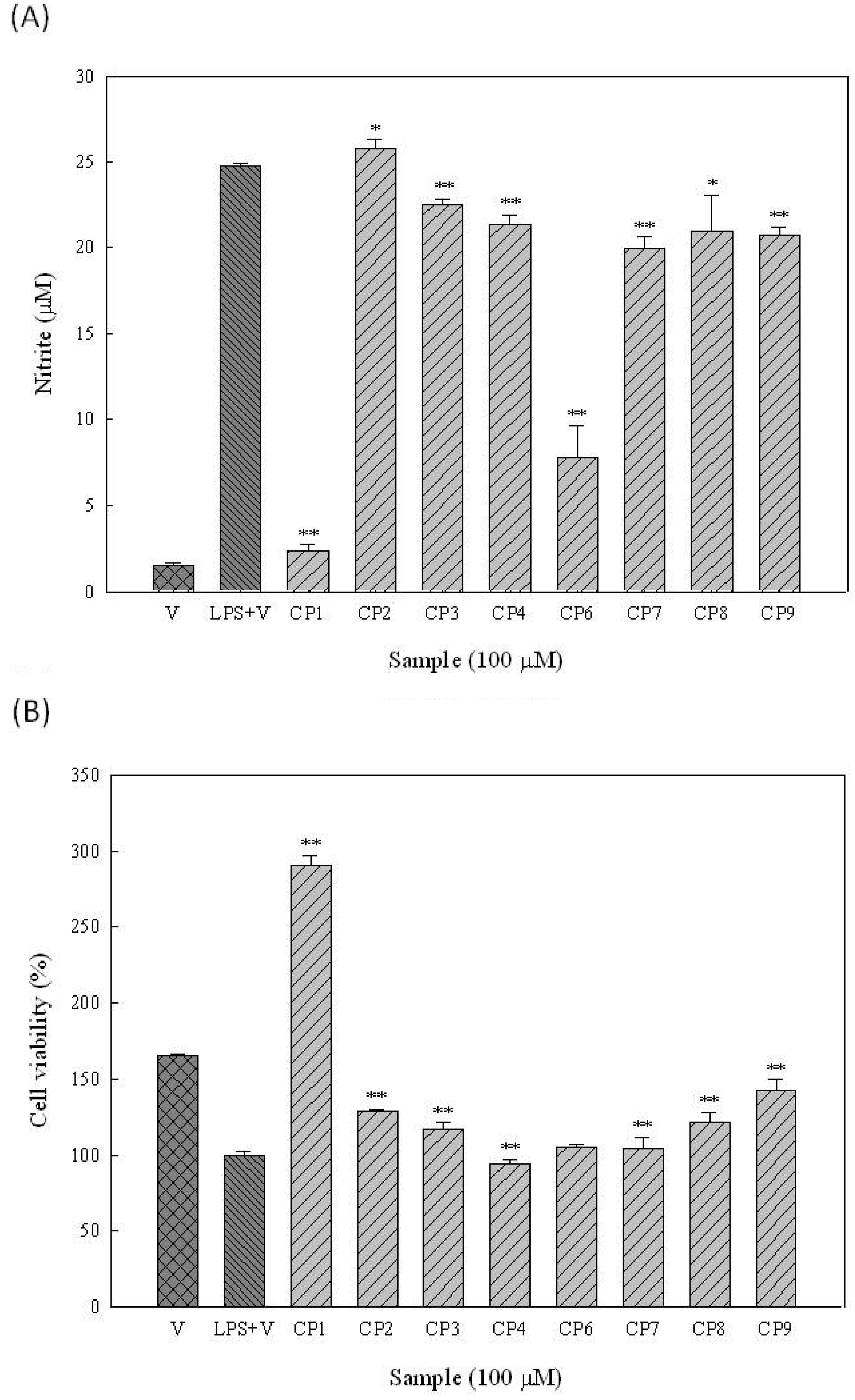
3.6. NBT (Superoxide Scavenging) Assay
3.7. Cell Culture
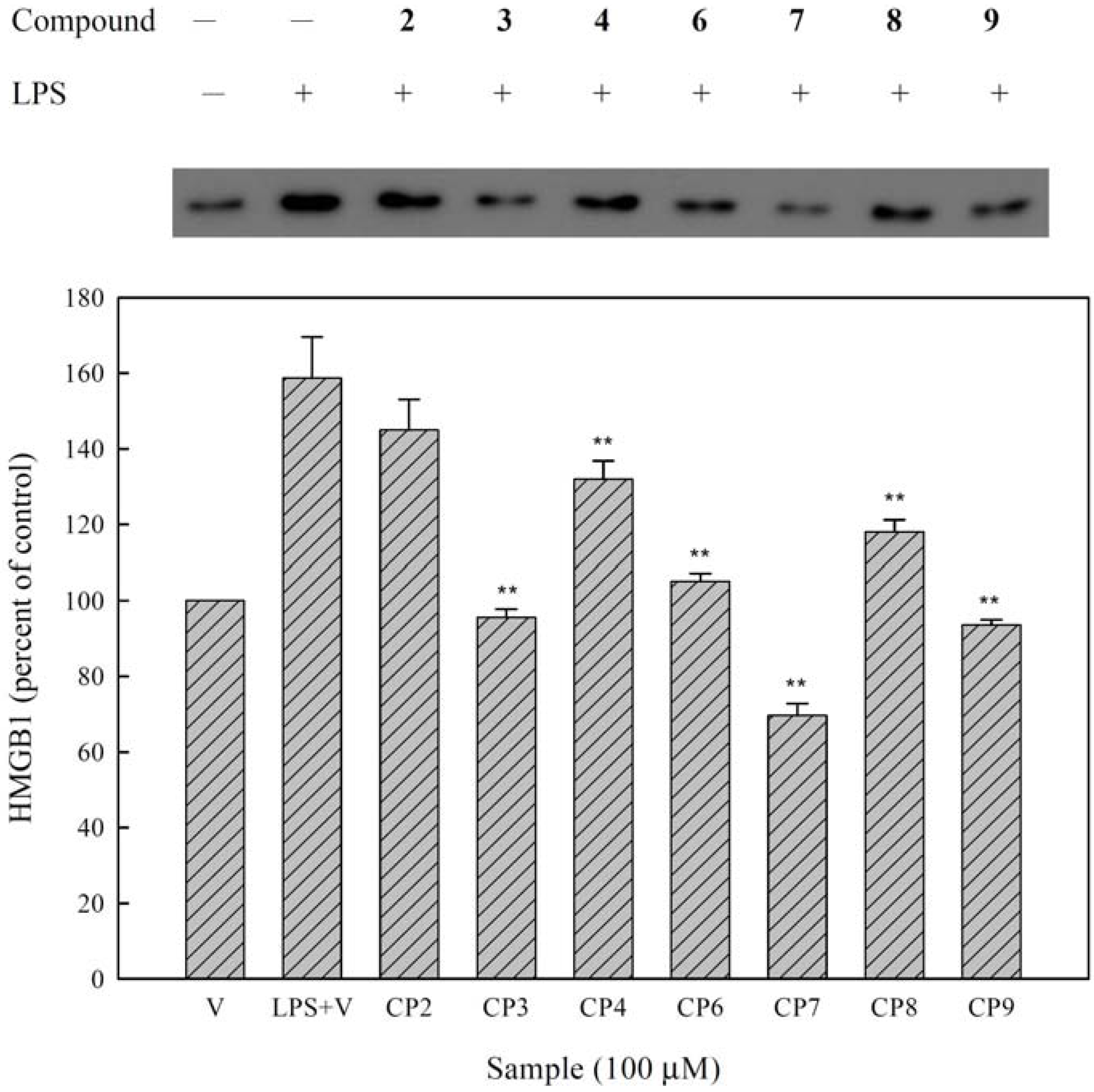
3.8. Western Blot Assay for HMGB-1
3.9. AlamarBlue Assay and Measurement of NO
3.10. RNA Extraction and RT-PCR
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Farooqui, T.; Farooqui, A.A. Aging: An important factor for the pathogenesis of neurodegenerative diseases. Mech. Ageing Dev. 2009, 130, 203–215. [Google Scholar] [CrossRef]
- Balsano, C.; Alisi, A. Antioxidant effects of natural bioactive compounds. Curr. Pharm. Des. 2009, 15, 3063–3073. [Google Scholar] [CrossRef]
- Roome, T.; Dar, A.; Ali, S.; Naqvi, S.; Choudhary, M.I. A study on antioxidant, free radical scavenging, anti-inflammatory and hepatoprotective actions of Aegiceras corniculatum (stem) extracts. J. Ethnopharmacol. 2008, 118, 514–521. [Google Scholar] [CrossRef]
- Salvemini, D.; Mazzon, E.; Dugo, L.; Riley, D.P.; Serraino, I.; Caputi, A.P.; Cuzzocrea, S. Pharmacological manipulation of the inflammatory cascade by the superoxide dismutase mimetic, M40403. Br. J. Pharmacol. 2001, 132, 815–827. [Google Scholar] [CrossRef]
- Lundberg, I.E. The role of cytokines, chemokines, and adhesion molecules in the pathogenesis of idiopathic inflammatory myopathies. Curr. Rheumatol. Rep. 2000, 2, 216–224. [Google Scholar] [CrossRef]
- Tao, J.Y.; Zhao, L.; Huang, Z.J.; Zhang, X.Y.; Zhang, S.L.; Zhang, Q.G.; Fei, X.; Zhang, B.H.; Feng, Q.L.; Zheng, G.H. Anti-inflammatory effects of ethanol extract from Kummerowia striata (Thunb.) Schindl on LPS-stimulated RAW264.7 cell. Inflammation 2008, 31, 154–166. [Google Scholar] [CrossRef]
- Annane, D.; Bellissant, E.; Cavaillon, J.M. Septic shock. Lancet 2005, 365, 63–78. [Google Scholar] [CrossRef]
- Lotze, M.T.; Tracey, K.J. High-mobility group box 1 protein (HMGB1): Nuclear weapon in the immune arsenal. Nat. Rev. Immunol. 2005, 5, 331–342. [Google Scholar] [CrossRef]
- Gardella, S.; Andrei, C.; Ferrera, D.; Lotti, L.V.; Torrisi, M.R.; Bianchi, M.E.; Rubartelli, A. The nuclear protein HMGB1 is secreted by monocytes via a non-classical, vesicle-mediated secretory pathway. EMBO Rep. 2002, 3, 995–1001. [Google Scholar] [CrossRef]
- Li, J.; Wang, H.; Mason, J.M.; Levine, J.; Yu, M.; Ulloa, L.; Czura, C.J.; Tracey, K.J.; Yang, H. Recombinant HMGB1 with cytokine-stimulating activity. J. Immunol. Methods 2004, 289, 211–223. [Google Scholar] [CrossRef]
- Ulloa, L. The vagus nerve and the nicotinic anti-inflammatory pathway. Nat. Rev. Drug Discov. 2005, 4, 673–684. [Google Scholar] [CrossRef]
- Ulloa, L.; Tracey, K.J. The "cytokine profile": A code for sepsis. Trends Mol. Med. 2005, 11, 56–63. [Google Scholar] [CrossRef]
- Han, S.Y.; Li, P.P. Progress of research in antitumor mechanisms with chinese medicine. Chin. J. Integr. Med. 2009, 15, 316–320. [Google Scholar] [CrossRef]
- Benavente Garcia, O.; Castillo, J. Update on uses and properties of citrus flavonoids: New findings in anticancer, cardiovascular, and anti-inflammatory activity. J. Agric. Food Chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef]
- Araldi, E.M.; Dell'aica, I.; Sogno, I.; Lorusso, G.; Garbisa, S.; Albini, A. Natural and synthetic agents targeting inflammation and angiogenesis for chemoprevention of prostate cancer. Curr. Cancer Drug Targets 2008, 8, 146–155. [Google Scholar] [CrossRef]
- Chu, Y.F.; Sun, J.; Wu, X.; Liu, R.H. Antioxidant and antiproliferative activities of common vegetables. J. Agric. Food Chem. 2002, 50, 6910–6916. [Google Scholar] [CrossRef]
- Liao, J.C. Lauraceae in Flora of Taiwan; Editorial Committee of the Flora of Taiwan: Taipei, Taiwan, 1996; Volume 2, p. 483. [Google Scholar]
- Cheng, M.J.; Tsai, I.L.; Lee, S.J.; Jayaprakasam, B.; Chen, I.S. Steryl epoxide, secobutanolide and butanolides from the stem wood of Machilus zuihoensis. Phytochemistry 2005, 66, 1180–1185. [Google Scholar] [CrossRef]
- Lee, M.H.; Jiang, C.B.; Juan, S.H.; Lin, R.D.; Hou, W.C. Antioxidant and heme oxygenase-1 (HO-1)-induced effects of selected Taiwanese plants. Fitoterapia 2006, 77, 109–115. [Google Scholar] [CrossRef]
- Hsieh, T.J.; Wu, Y.C.; Chen, C.Y. Chemical constituents from the leaves of Epimedium sagittatum. Chin. Pharm. J. 2003, 55, 121–128. [Google Scholar]
- Lu, Y.; Foo, L.Y. Identification and quantification of major polyphenols in apple pomace. Food Chem. 1997, 59, 187–194. [Google Scholar] [CrossRef]
- Rao, L.J.M.; Yada, H.; Ono, H.; Yoshida, M. Acylated and non-acylated flavonol monoglycosides from the indian minor spice Nagkesar (Mammea longifolia). J. Agric. Food Chem. 2002, 50, 3143–3146. [Google Scholar] [CrossRef]
- Kim, H.; Ralph, J.; Lu, F.; Ralph, S.A.; Boudet, A.M.; MacKay, J.J.; Sederoff, R.R.; Ito, T.; Kawai, S.; Ohashi, H.; Higuchi, T. NMR analysis of lignins in CAD-deficient plants. Part 1. Incorporation of hydroxycinnamaldehydes and hydroxybenzaldehydes into lignins. Org. Biomol. Chem. 2003, 1, 268–281. [Google Scholar] [CrossRef]
- Etzenhouser, B.; Hansch, C.; Kapur, S.; Selassie, C.D. Mechanism of toxicity of esters of caffeic and dihydrocaffeic acids. Bioorg. Med. Chem. 2001, 9, 199–209. [Google Scholar]
- Wang, Y.; Hamburger, M.; Gueho, J.; Hostettmann, K. Cyclohexanecarboxylic-acid derivatives from Psiadia trinervia. Helv. Chim. Acta 1992, 75, 269–275. [Google Scholar] [CrossRef]
- Zhu, X.; Dong, X.; Wang, Y.; Peng, J.; Luo, S. Phenolic compounds from Viburnum cylindricum. Helv. Chim. Acta 2005, 88, 339–342. [Google Scholar] [CrossRef]
- Amic, D.; Davidovic Amic, D.; Beslo, D.; Rastija, V.; Lucic, B.; Trinajstic, N. SAR and QSAR of the antioxidant activity of flavonoids. Curr. Med. Chem. 2007, 14, 827–845. [Google Scholar]
- Cos, P.; Ying, L.; Calomme, M.; Hu, J.P.; Cimanga, K.; van Poel, B.; Pieters, L.; Vlietinck, A.J.; Vanden Berghe, D. Structure-activity relationship and classification of flavonoids as inhibitors of xanthine oxidase and superoxide scavengers. J. Nat. Prod. 1998, 61, 71–76. [Google Scholar] [CrossRef]
- Lowe, J.B. Glycan-dependent leukocyte adhesion and recruitment in inflammation. Curr. Opin. Cell Biol. 2003, 15, 531–538. [Google Scholar] [CrossRef]
- Kim, S.S.; Oh, O.J.; Min, H.-Y.; Park, E.J.; Kim, Y.; Park, H.J.; Han, Y.N.; Lee, S.K. Eugenol suppresses cyclooxygenase-2 expression in lipopolysaccharide-stimulated mouse macrophage RAW264. 7 cells. Life Sci. 2003, 73, 337–348. [Google Scholar]
- Qin, Y.H.; Dai, S.M.; Tang, G.S.; Zhang, J.; Ren, D.; Wang, Z.W.; Shen, Q. HMGB1 enhances the proinflammatory activity of lipopolysaccharide by promoting the phosphorylation of MAPK p38 through receptor for advanced glycation end products. J. Immunol. 2009, 183, 6244–6250. [Google Scholar] [CrossRef]
- Ulloa, L.; Ochani, M.; Yang, H.; Tanovic, M.; Halperin, D.; Yang, R.; Czura, C.J.; Fink, M.P.; Tracey, K.J. Ethyl pyruvate prevents lethality in mice with established lethal sepsis and systemic inflammation. Proc. Natl. Acad. Sci. USA 2002, 99, 12351–12356. [Google Scholar]
- Wang, H.; Liao, H.; Ochani, M.; Justiniani, M.; Lin, X.; Yang, L.; Al-Abed, Y.; Wang, H.; Metz, C.; Miller, E.J.; et al. Cholinergic agonists inhibit HMGB1 release and improve survival in experimental sepsis. Nat. Med. 2004, 10, 1216–1221. [Google Scholar] [CrossRef]
- Manjeet, K.R.; Ghosh, B. Quercetin inhibits LPS-induced nitric oxide and tumor necrosis factor-alpha production in murine macrophages. Int. J. Immunopharmacol. 1999, 21, 435–443. [Google Scholar] [CrossRef]
- Comalada, M.; Camuesco, D.; Sierra, S.; Ballester, I.; Xaus, J.; Galvez, J.; Zarzuelo, A. In vivo quercitrin anti-inflammatory effect involves release of quercetin, which inhibits inflammation through down-regulation of the NF-κB pathway. Eur. J. Immunol. 2005, 35, 584–592. [Google Scholar] [CrossRef]
- Liu, J.; Li, X.; Yue, Y.; Li, J.; He, T.; He, Y. The inhibitory effect of quercetin on IL-6 production by LPS-stimulated neutrophils. Cell. Mol. Immunol. 2005, 2, 455–460. [Google Scholar]
- Tang, D.; Kang, R.; Xiao, W.; Zhang, H.; Lotze, M.T.; Wang, H.; Xiao, X. Quercetin prevents LPS-induced high-mobility group box 1 release and proinflammatory function. Am. J. Respir. Cell Mol. Biol. 2009, 41, 651–660. [Google Scholar] [CrossRef]
- Chiang, Y.M.; Lo, C.P.; Chen, Y.P.; Wang, S.Y.; Yang, N.S.; Kuo, Y.H.; Shyur, L.F. Ethyl caffeate suppresses NF-κB activation and its downstream inflammatory mediators, iNOS, COX-2, and PGE2 in vitro or in mouse skin. Br. J. Pharmacol. 2005, 146, 352–363. [Google Scholar] [CrossRef]
- Feghali, C.A.; Wright, T.M. Cytokines in acute and chronic inflammation. Front. Biosci. 1997, 2, d12–d26. [Google Scholar]
- Kim, J.H.; Kim, S.J.; Lee, I.S.; Lee, M.S.; Uematsu, S.; Akira, S.; Oh, K.I. Bacterial endotoxin induces the release of high mobility group box 1 via the IFN-β signaling pathway. J. Immunol. 2009, 182, 2458–2466. [Google Scholar] [CrossRef]
- Hsu, F.L.; Chou, C.J.; Chang, Y.C.; Chang, T.T.; Lu, M.K. Promotion of hyphal growth and underlying chemical changes in Antrodia camphorata by host factors from Cinnamomum camphora. Int. J. Food Microbiol. 2006, 106, 32–38. [Google Scholar] [CrossRef]
- Valentao, P.; Fernandes, E.; Carvalho, F.; Andrade, P.B.; Seabra, R.M.; Bastos, M.L. Antioxidant activity of Centaurium erythraea infusion evidenced by its superoxide radical scavenging and xanthine oxidase inhibitory activity. J. Agric. Food Chem. 2001, 49, 3476–3479. [Google Scholar] [CrossRef]
- Wang, H.; Bloom, O.; Zhang, M.; Vishnubhakat, J.M.; Ombrellino, M.; Che, J.; Frazier, A.; Yang, H.; Ivanova, S.; Borovikova, L.; et al. HMG-1 as a late mediator of endotoxin lethality in mice. Science 1999, 285, 248–251. [Google Scholar] [CrossRef]
- Rendon Mitchell, B.; Ochani, M.; Li, J.; Han, J.; Wang, H.; Yang, H.; Susarla, S.; Czura, C.; Mitchell, R.A.; Chen, G.; et al. IFN-gamma induces high mobility group box 1 protein release partly through a TNF-dependent mechanism. J. Immunol. 2003, 170, 3890–3897. [Google Scholar]
- Nociari, M.M.; Shalev, A.; Benias, P.; Russo, C. A novel one-step, highly sensitive fluorometric assay to evaluate cell-mediated cytotoxicity. J. Immunol. Methods 1998, 213, 157–167. [Google Scholar] [CrossRef]
- Guevara, I.; Iwanejko, J.; Dembinska-Kiec, A.; Pankiewicz, J.; Wanat, A.; Anna, P.; Golabek, I.; Bartus, S.; Malczewska-Malec, M.; Szczudlik, A. Determination of nitrite/nitrate in human biological material by the simple Griess reaction. Clin. Chim. Acta 1998, 274, 177–188. [Google Scholar] [CrossRef]
- Horton, J.W. Free radicals and lipid peroxidation mediated injury in burn trauma: The role of antioxidant therapy. Toxicology 2003, 189, 75–88. [Google Scholar] [CrossRef]
- Rahman, I. Oxidative stress and gene transcription in asthma and chronic obstructive pulmonary disease: Antioxidant therapeutic targets. Curr. Drug Targets Inflamm. Allergy 2002, 1, 291–315. [Google Scholar] [CrossRef]
- Sample Availability: Please contact the corresponding author.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mao, Y.-W.; Tseng, H.-W.; Liang, W.-L.; Chen, I.-S.; Chen, S.-T.; Lee, M.-H. Anti-Inflammatory and Free Radial Scavenging Activities of the Constituents Isolated from Machilus zuihoensis. Molecules 2011, 16, 9451-9466. https://doi.org/10.3390/molecules16119451
Mao Y-W, Tseng H-W, Liang W-L, Chen I-S, Chen S-T, Lee M-H. Anti-Inflammatory and Free Radial Scavenging Activities of the Constituents Isolated from Machilus zuihoensis. Molecules. 2011; 16(11):9451-9466. https://doi.org/10.3390/molecules16119451
Chicago/Turabian StyleMao, Yi-Wen, Hsiang-Wen Tseng, Wen-Li Liang, Ih-Sheng Chen, Shui-Tein Chen, and Mei-Hsien Lee. 2011. "Anti-Inflammatory and Free Radial Scavenging Activities of the Constituents Isolated from Machilus zuihoensis" Molecules 16, no. 11: 9451-9466. https://doi.org/10.3390/molecules16119451
APA StyleMao, Y. -W., Tseng, H. -W., Liang, W. -L., Chen, I. -S., Chen, S. -T., & Lee, M. -H. (2011). Anti-Inflammatory and Free Radial Scavenging Activities of the Constituents Isolated from Machilus zuihoensis. Molecules, 16(11), 9451-9466. https://doi.org/10.3390/molecules16119451