The Effect of Curcumin on Human Bronchial Epithelial Cells Exposed to Fine Particulate Matter: A Predictive Analysis


Abstract
:1. Introduction
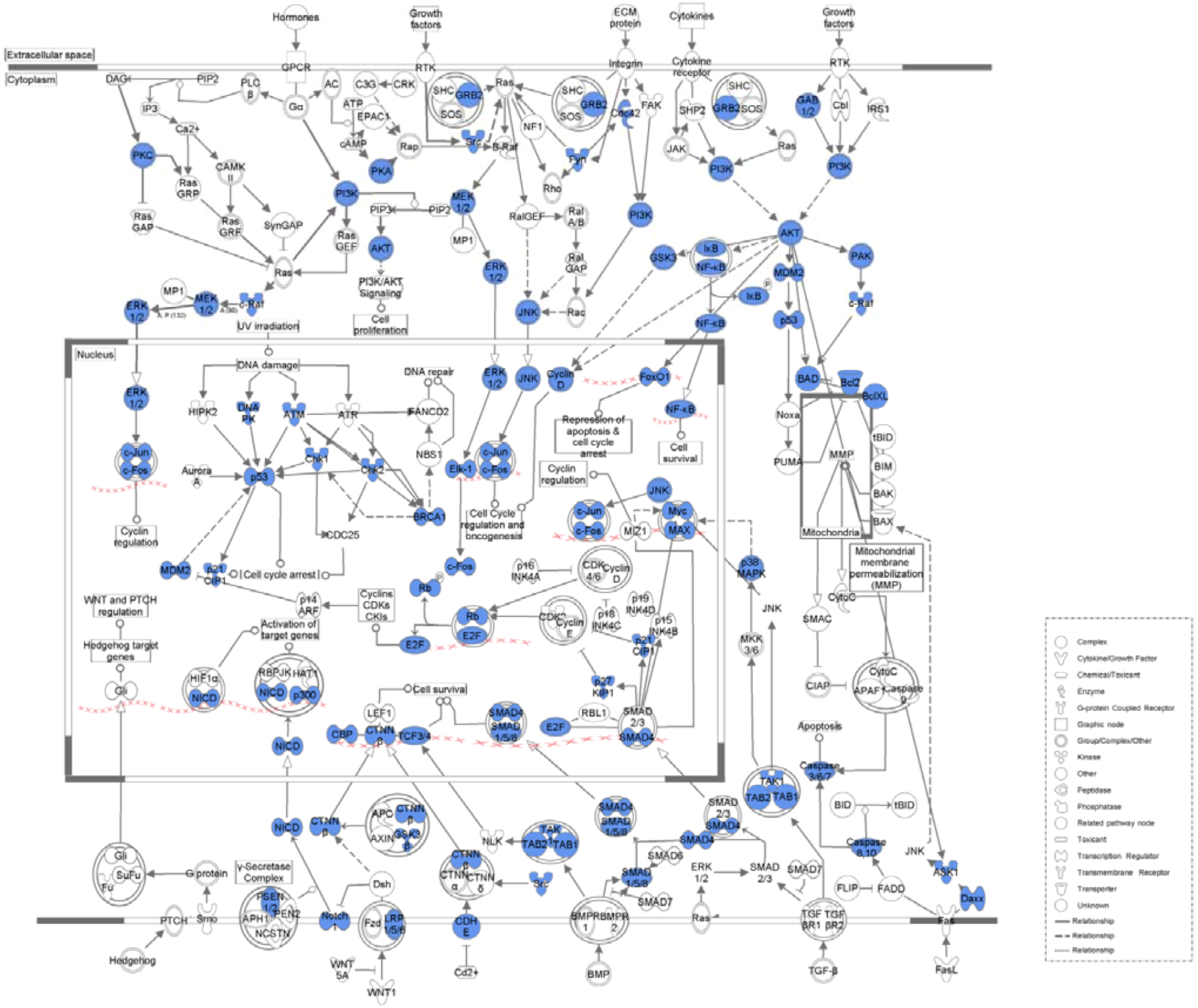
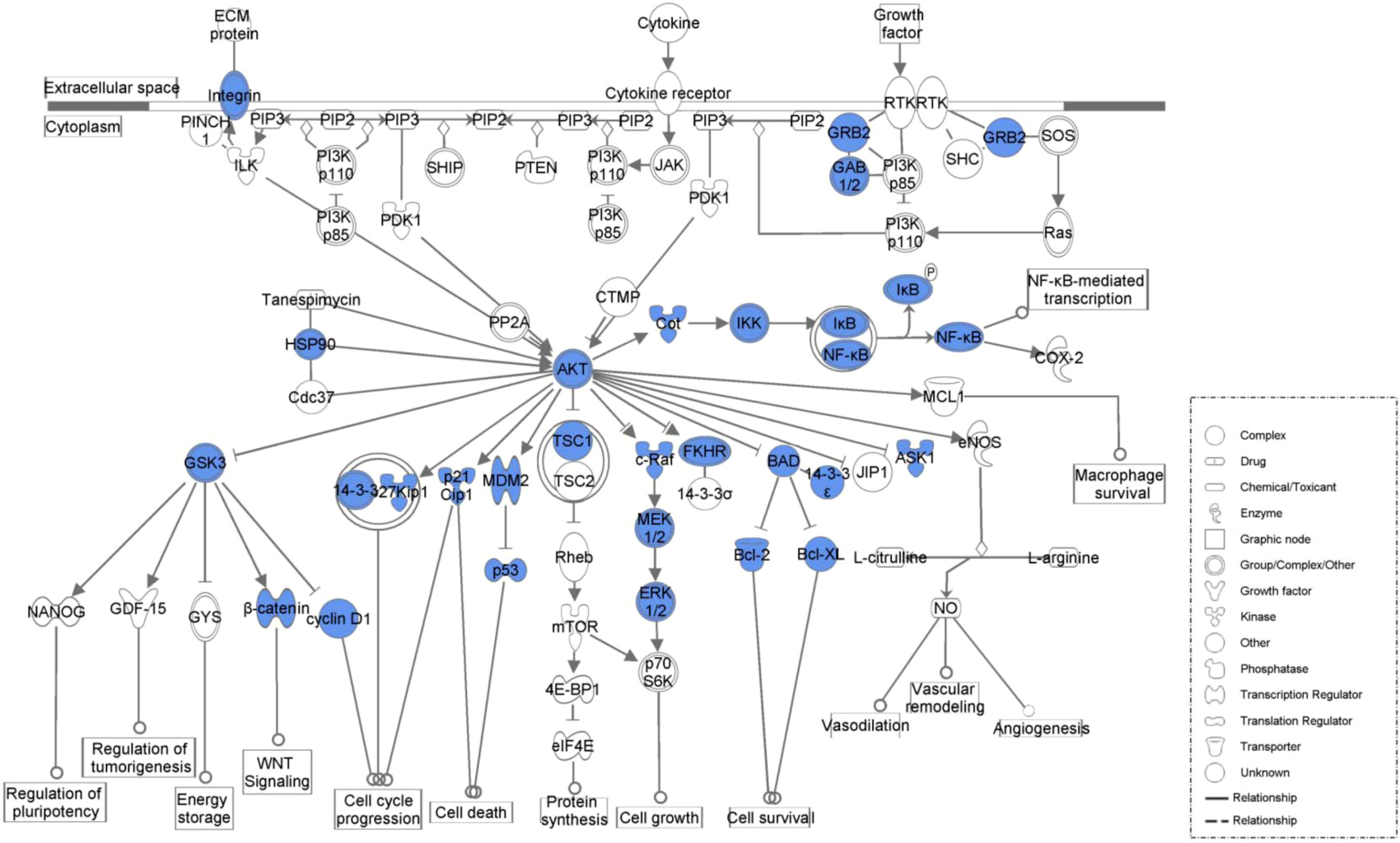
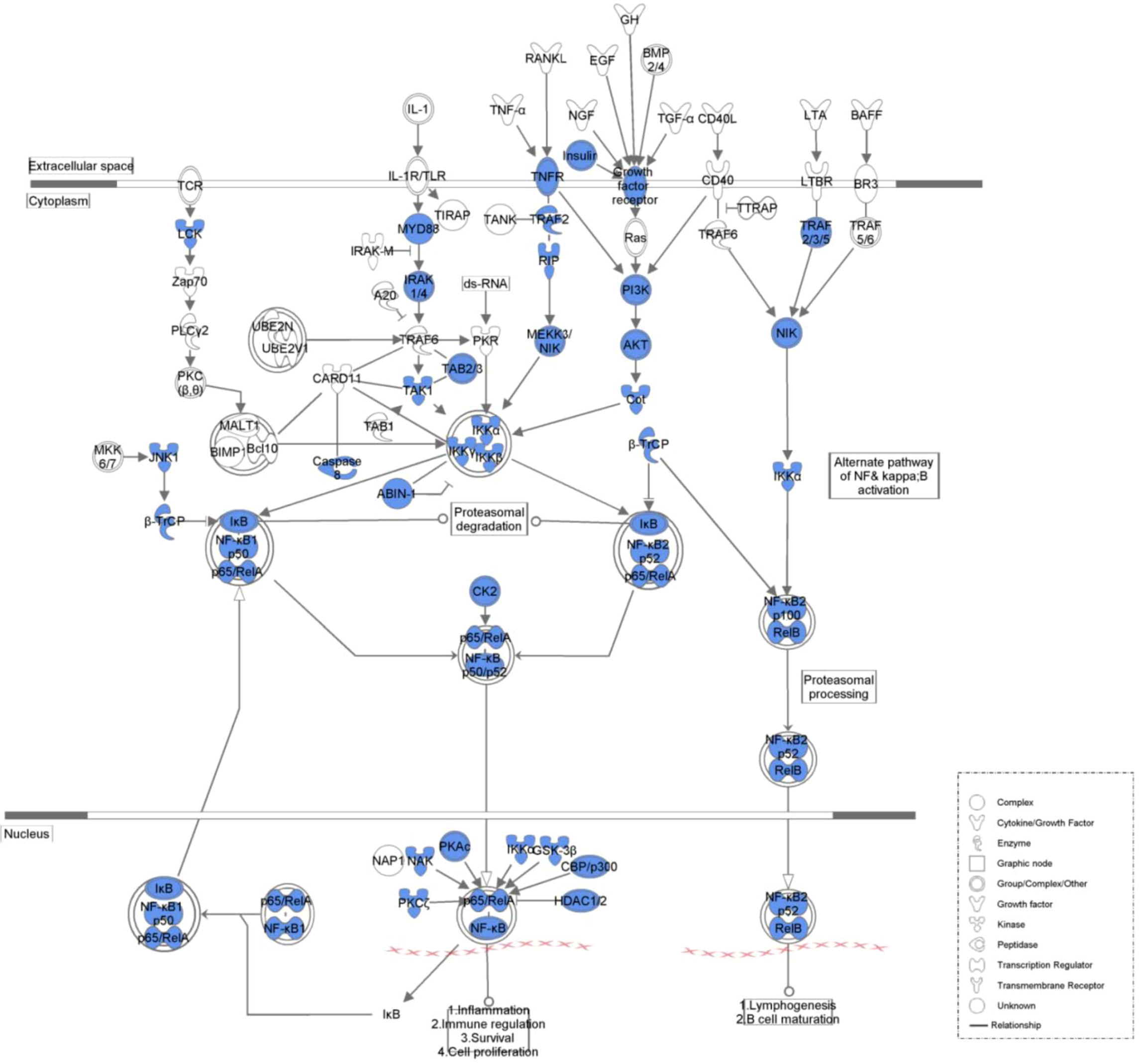
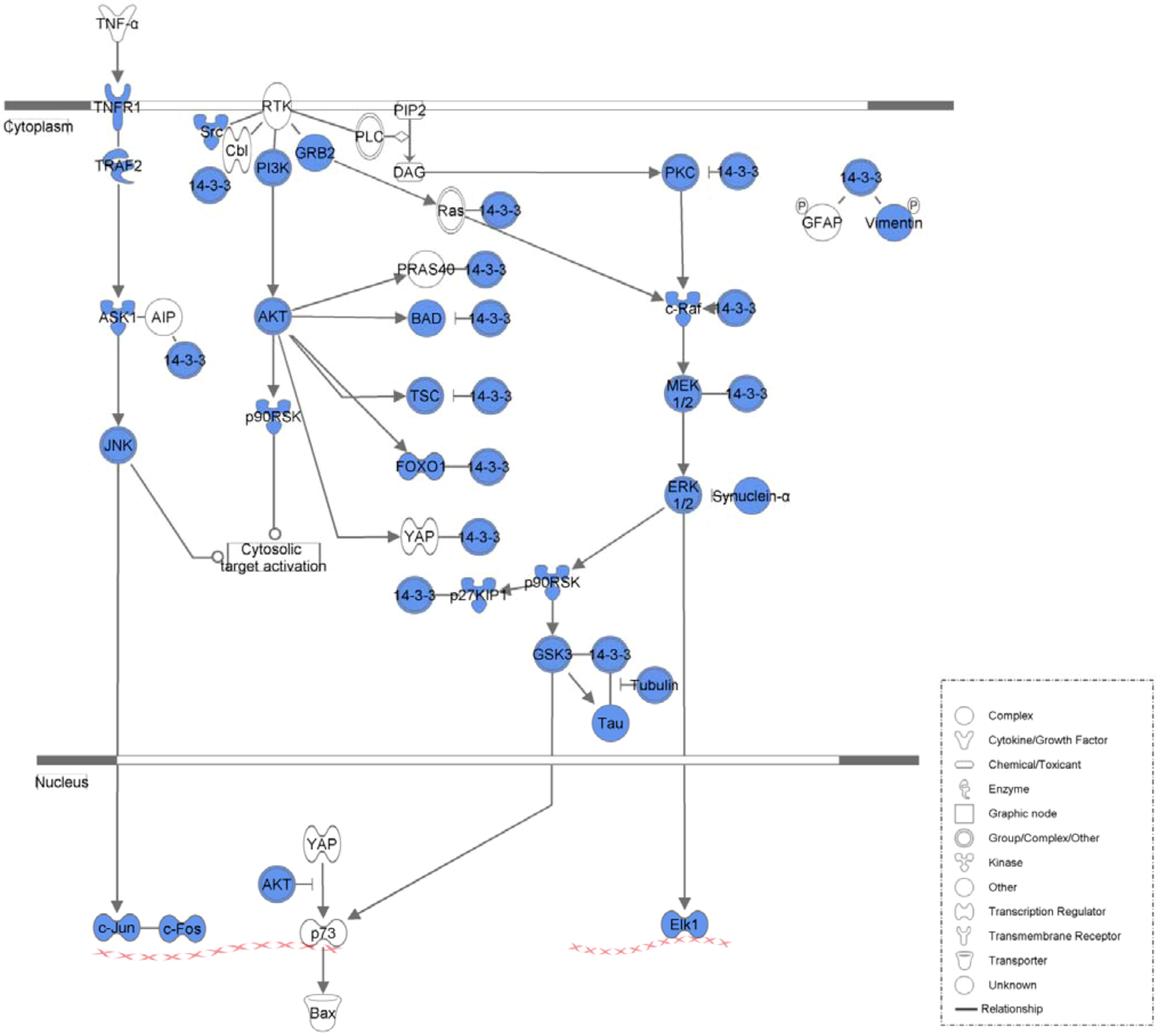
2. Results and Discussion
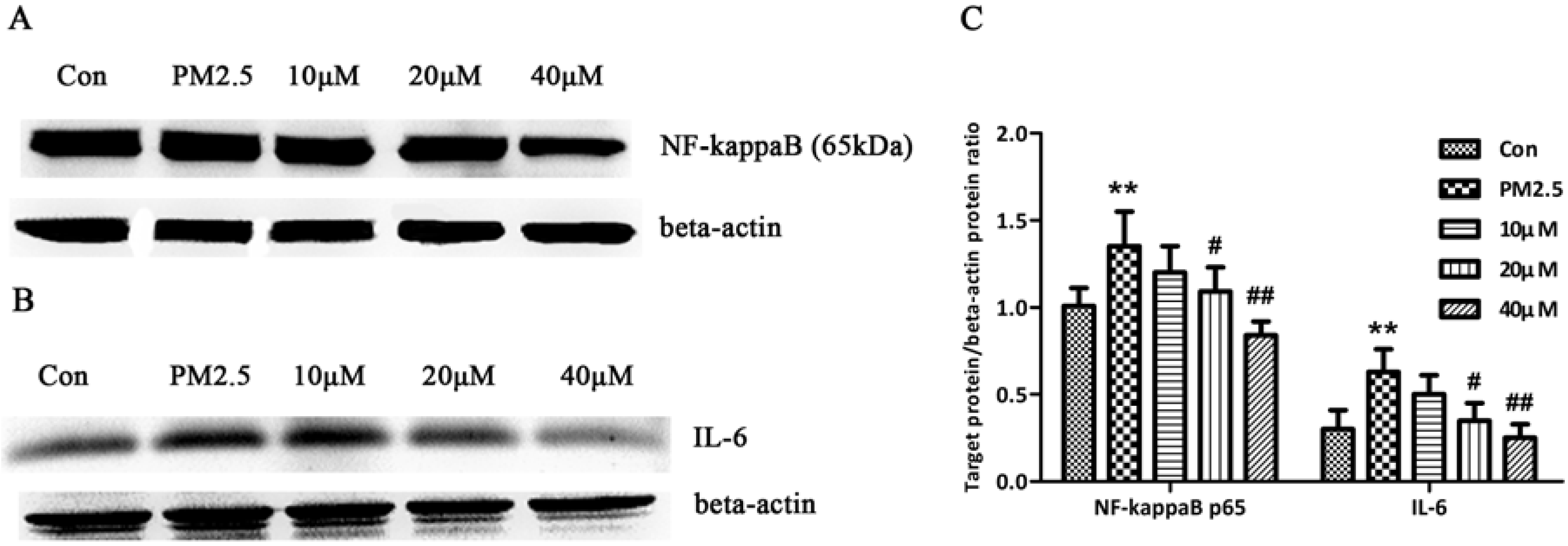
2.1. Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe Set ID | RefSeq ID | Gene Symbol | p-value | Fold Change | Regulation |
|---|---|---|---|---|---|
| 203665_at | NM_002133 | HMOX1 | 0.0045 | 24.58 | up |
| 209921_at | NM_014331 | SLC7A11 | 0.0004 | 10.45 | up |
| 202436_s_at | NM_000104 | CYP1B1 | 0.0015 | 7.02 | up |
| 201266_at | NM_003330 | TXNRD1 | 0.0014 | 6.96 | up |
| 205749_at | NM_000499 | CYP1A1 | 0.0185 | 5.54 | up |
| 203925_at | NM_002061 | GCLM | 0.0006 | 3.71 | up |
| 202923_s_at | NM_001498 | GCLC | 0.0110 | 3.29 | up |
| 201468_s_at | NM_000903 | NQO1 | 0.0139 | 2.92 | up |
| 204151_x_at | NM_001353 | AKR1C1 | 0.0042 | 2.91 | up |
| 206172_at | NM_000640 | IL13RA2 | 0.0110 | 2.72 | up |
| 211653_x_at | NM_001354 | AKR1C2 | 0.0083 | 2.70 | up |
| 209387_s_at | NM_014220 | TM4SF1 | 0.0472 | 2.47 | up |
| 210845_s_at | NM_002659 | PLAUR | 0.0104 | 2.18 | up |
| 206683_at | NM_003447 | ZNF165 | 0.0090 | 2.14 | up |
| 212907_at | NM_021194 | SLC30A1 | 0.0357 | 2.14 | up |
| 214211_at | NM_002032 | FTH1 | 0.0270 | 2.08 | up |
| 208963_x_at | NM_013402 | FADS1 | 0.0202 | 2.03 | up |
| 205767_at | NM_001432 | EREG | 0.0160 | 1.98 | up |
| 219475_at | NM_182981 | OSGIN1 | 0.0103 | 1.98 | up |
| 207675_x_at | NM_057091 | ARTN | 0.0313 | 1.97 | up |
| 202842_s_at | NM_012328 | DNAJB9 | 0.0309 | 1.96 | up |
| 202266_at | NM_016614 | TDP2 | 0.0001 | 1.95 | up |
| 201625_s_at | NM_005542 | INSIG1 | 0.0446 | 1.93 | up |
| 209882_at | NM_006912 | RIT1 | 0.0114 | 1.93 | up |
| 201489_at | NM_005729 | PPIF | 0.0093 | 1.92 | up |
| 213112_s_at | NM_003900 | SQSTM1 | 0.0191 | 1.91 | up |
| 204420_at | NM_005438 | FOSL1 | 0.0323 | 1.80 | up |
| 202284_s_at | NM_000389 | CDKN1A | 0.0324 | 1.77 | up |
| 206907_at | NM_003811 | TNFSF9 | 0.0032 | 1.74 | up |
| 219697_at | NM_006043 | HS3ST2 | 0.0291 | 1.72 | up |
| 204970_s_at | NM_002359 | MAFG | 0.0032 | 1.69 | up |
| 213187_x_at | NM_000146 | FTL | 0.0471 | 1.68 | up |
| 212717_at | NM_014798 | PLEKHM1 | 0.0319 | 1.66 | up |
| 206498_at | NM_000275 | OCA2 | 0.0221 | 1.66 | up |
| 202672_s_at | NM_001674 | ATF3 | 0.0041 | 1.57 | up |
| 202021_x_at | NM_005801 | EIF1 | 0.0460 | 1.55 | up |
| 202067_s_at | NM_000527 | LDLR | 0.0128 | 1.54 | up |
| 204958_at | NM_004073 | PLK3 | 0.0153 | 1.50 | up |
| 202207_at | NM_005737 | ARL4C | 0.0139 | 3.24 | down |
| 202887_s_at | NM_019058 | DDIT4 | 0.0123 | 2.66 | down |
| 201890_at | NM_001034 | RRM2 | 0.0097 | 2.26 | down |
| 211450_s_at | NM_000179 | MSH6 | 0.0486 | 2.26 | down |
| 201849_at | NM_004052 | BNIP3 | 0.0293 | 2.20 | down |
| 219250_s_at | NM_013281 | FLRT3 | 0.0425 | 2.10 | down |
| 209120_at | NM_021005 | NR2F2 | 0.0056 | 2.01 | down |
| 202464_s_at | NM_004566 | PFKFB3 | 0.0289 | 1.99 | down |
| 208808_s_at | NM_002129 | HMGB2 | 0.0462 | 1.99 | down |
| 203344_s_at | NM_002894 | RBBP8 | 0.0280 | 1.97 | down |
| 218718_at | NM_016205 | PDGFC | 0.0044 | 1.97 | down |
| 207173_x_at | NM_001797 | CDH11 | 0.0432 | 1.95 | down |
| 201669_s_at | NM_002356 | MARCKS | 0.0448 | 1.92 | down |
| 207826_s_at | NM_002167 | ID3 | 0.0285 | 1.84 | down |
| 204967_at | NM_001649 | SHROOM2 | 0.0141 | 1.80 | down |
| 202628_s_at | NM_000602 | SERPINE1 | 0.0486 | 1.77 | down |
| 212599_at | NM_015570 | AUTS2 | 0.0053 | 1.77 | down |
| 203274_at | NM_012151 | F8A1 | 0.0118 | 1.76 | down |
| 208673_s_at | NM_003017 | SRSF3 | 0.0188 | 1.76 | down |
| 203476_at | NM_006670 | TPBG | 0.0400 | 1.75 | down |
| 209189_at | NM_005252 | FOS | 0.0366 | 1.72 | down |
| 209784_s_at | NM_002226 | JAG2 | 0.0032 | 1.70 | down |
| 203625_x_at | NM_005983 | SKP2 | 0.0040 | 1.70 | down |
| 222036_s_at | NM_005914 | MCM4 | 0.0439 | 1.66 | down |
| 202219_at | NM_005629 | SLC6A8 | 0.0253 | 1.65 | down |
| 205449_at | NM_013299 | SAC3D1 | 0.0328 | 1.65 | down |
| 212168_at | NM_006047 | RBM12 | 0.0031 | 1.64 | down |
| 209286_at | NM_006449 | CDC42EP3 | 0.0060 | 1.63 | down |
| 204334_at | NM_003709 | KLF7 | 0.0105 | 1.63 | down |
| 208579_x_at | NM_017445 | H2BFS | 0.0173 | 1.62 | down |
| 204069_at | NM_002398 | MEIS1 | 0.0281 | 1.60 | down |
| 203797_at | NM_003385 | VSNL1 | 0.0172 | 1.58 | down |
| 203764_at | NM_014750 | DLGAP5 | 0.0181 | 1.58 | down |
| 213051_at | NM_020119 | ZC3HAV1 | 0.0104 | 1.58 | down |
| 208051_s_at | NM_006451 | PAIP1 | 0.0321 | 1.57 | down |
| 203405_at | NM_003720 | PSMG1 | 0.0304 | 1.57 | down |
| 211744_s_at | NM_001779 | CD58 | 0.0273 | 1.57 | down |
| 206277_at | NM_002564 | P2RY2 | 0.0179 | 1.56 | down |
| 204715_at | NM_015368 | PANX1 | 0.0375 | 1.56 | down |
| 201312_s_at | NM_003022 | SH3BGRL | 0.0383 | 1.55 | down |
| 213088_s_at | NM_015190 | DNAJC9 | 0.0253 | 1.55 | down |
| 203803_at | NM_016297 | PCYOX1 | 0.0350 | 1.54 | down |
| 201624_at | NM_001349 | DARS | 0.0225 | 1.54 | down |
| 214214_s_at | NM_001212 | C1QBP | 0.0468 | 1.54 | down |
| 212320_at | NM_178014 | TUBB | 0.0185 | 1.53 | down |
| 208405_s_at | NM_006016 | CD164 | 0.0465 | 1.51 | down |
| 213019_at | NM_012416 | RANBP6 | 0.0002 | 1.51 | down |
| 212922_s_at | NM_020197 | SMYD2 | 0.0002 | 1.50 | down |
| 209025_s_at | NM_006372 | SYNCRIP | 0.0481 | 1.50 | down |
| 201163_s_at | NM_001553 | IGFBP7 | 0.0458 | 1.50 | down |
| 214800_x_at | NM_001207 | BTF3 | 0.0036 | 1.50 | down |
| GI | UniProtKB ID |
|---|---|
| 4507949 | 1433B_HUMAN |
| 31542303 | ABHD5_HUMAN |
| 37622910 | ACM1_HUMAN |
| 21361176 | AL1A1_HUMAN |
| 4885057 | APJ_HUMAN |
| 47132611 | ATG4B_HUMAN |
| 6683500 | BAZ2B_HUMAN |
| 53832009 | CAC1H_HUMAN |
| 4502601 | CBR3_HUMAN |
| 37187860 | CCR6_HUMAN |
| 67551261 | CLK1_HUMAN |
| 153791372 | CLK3_HUMAN |
| 13435386 | CP3A4_HUMAN |
| 32307159 | CRFR2_HUMAN |
| 30219 | CRHBP_HUMAN |
| 4503383 | DRD1_HUMAN |
| 4503385 | DRD2_HUMAN |
| 10835013 | ESR2_HUMAN |
| 4885263 | GEM_HUMAN |
| 122921310 | HCD2_HUMAN |
| 155969707 | IDE_HUMAN |
| 98986450 | KC1G1_HUMAN |
| 153791733 | KC1G2_HUMAN |
| 325651834 | KCNH2_HUMAN |
| 221046486 | KD4DL_HUMAN |
| 22035600 | M4K2_HUMAN |
| 11386165 | MCL1_HUMAN |
| 89993689 | MDM2_HUMAN |
| 88702791 | MDM4_HUMAN |
| 20986531 | MK01_HUMAN |
| 4505209 | MMP13_HUMAN |
| 66911845 | MRGX1_HUMAN |
| 34577122 | NFKB1_HUMAN |
| 222080095 | OX1R_HUMAN |
| 32307152 | OXYR_HUMAN |
| 4505587 | PA1B3_HUMAN |
| 5031975 | PAK4_HUMAN |
| 31881630 | PE2R2_HUMAN |
| 31542939 | PGDH_HUMAN |
| 4505811 | PIM1_HUMAN |
| 42821112 | PIM2_HUMAN |
| 223718196 | PLIN1_HUMAN |
| 116734717 | PPBT_HUMAN |
| 4826962 | RAC3_HUMAN |
| 41281453 | SLK_HUMAN |
| 23943882 | STK33_HUMAN |
| 8400711 | TAU_HUMAN |
| 223468676 | TF65_HUMAN |
| 4507533 | TLR4_HUMAN |
| 8394456 | TLR9_HUMAN |
| 4507615 | TNNC1_HUMAN |
| 151101270 | TNNI3_HUMAN |
| 48255881 | TNNT2_HUMAN |
| 4507681 | TRFR_HUMAN |
| 118600387 | UBP1_HUMAN |
| 4502331 | V1AR_HUMAN |
| 4507883 | VDR_HUMAN |





| Top Bio Functions | p-value | Number of Molecules |
|---|---|---|
| Diseases and Disorders | ||
| Infectious Disease | 1.26E−12–4.25E−02 | 35 |
| Cancer | 3.45E–3.01E−02 | 8 |
| Genetic Disorder | 1.33E–3.01E−02 | 5 |
| Respiratory Disease | 1.33E–3.01E−02 | 6 |
| Inflammatory Response | 2.79E–2.79E−02 | 1 |
| Molecular and Cellular Functions | ||
| Cell Death | 9.91E−20–3.01E−02 | 31 |
| Cellular Growth and Proliferation | 5.64E−15–2.79E−02 | 32 |
| Cellular Development | 1.56E−08–2.79E−02 | 17 |
| Cell Cycle | 1.84E−07–2.79E−02 | 12 |
| Cellular Movement | 1.01E−04–2.79E−02 | 10 |
| Physiological System Development and Function | ||
| Organismal Survival | 2.02E−03–2.02E−03 | 4 |
| Respiratory System Development and Function | 2.28E−03–2.28E−03 | 2 |
| Tissue Development | 2.28E−03–2.79E−02 | 2 |
| Connective Tissue Development and Function | 1.94E−02–1.94E−02 | 2 |
| Tissue Morphology | 2.79E−02–2.79E−02 | 1 |
| Canonical Pathways | p-value | Ratio |
|---|---|---|
| Glucocorticoid Receptor Signaling | 2.57E−42 | 61/238 (0.256) |
| Molecular Mechanisms of Cancer | 6.68E−39 | 65/314 (0.207) |
| PI3K/AKT Signaling | 6.87E−36 | 41/110 (0.373) |
| NF-kappaB Signaling | 1.33E−30 | 41/143 (0.287) |
| 14-3-3-mediated Signaling | 1.37E−30 | 36/102 (0.353) |

2.2. Discussion
3. Experimental
3.1. Microarray Data Analysis
3.2. Target Proteins of Curcumin
3.3. Construction of PPI Networks and Detection of Common Network
3.4. Functional and Pathway Analysis of Common Network
3.5. Validating Experiment
3.5.1. Chemicals
3.5.2. Cell Culture
3.5.3. Preparation of Particles
3.5.4. Treatment of Cells with Curcumin and PM2.5
3.5.5. Western Blot
4. Conclusions
Supplementary Materials
Acknowledgments
- Sample Availability: Not Available.
References
- Valavanidis, A.; Fiotakis, K.; Vlachogianni, T. Airborne particulate matter and human health: toxicological assessment and importance of size and composition of particles for oxidative damage and carcinogenic mechanisms. J. Environ. Sci. Heal. C 2008, 26, 339–362. [Google Scholar]
- Sacks, J.D.; Stanek, L.W.; Luben, T.J.; Johns, D.O.; Buckley, B.J.; Brown, J.S.; Ross, M. Particulate matter-induced health effects: who is susceptible? Environ. Health Perspect. 2011, 119, 446–454. [Google Scholar]
- Polichetti, G.; Cocco, S.; Spinali, A.; Trimarco, V.; Nunziata, A. Effects of particulate matter (PM(10), PM(2.5) and PM(1)) on the cardiovascular system. Toxicology 2009, 261, 1–8. [Google Scholar] [CrossRef]
- Hatcher, H.; Planalp, R.; Cho, J.; Torti, F.M.; Torti, S.V. Curcumin: From ancient medicine to current clinical trials. Cell. Mol. Life Sci. 2008, 65, 1631–1652. [Google Scholar] [CrossRef]
- Zhou, H.; Beevers, C.S.; Huang, S. The targets of curcumin. Curr. Drug Targets 2011, 12, 332–347. [Google Scholar]
- Wolberg, J.R. Designing Quantitative Experiments : Prediction Analysis; Springer: Berlin, Germany, 2010. [Google Scholar]
- PubChem. Available online: http://pubchem.ncbi.nlm.nih.gov (accessed on 16 October 2012).
- Wang, Y.; Xiao, J.; Suzek, T.O.; Zhang, J.; Wang, J.; Zhou, Z.; Han, L.; Karapetyan, K.; Dracheva, S.; Shoemaker, B.A.; et al. PubChem’s BioAssay Database. Nucleic Acids Res. 2012, 40, D400–D412. [Google Scholar]
- Raman, K. Construction and analysis of protein-protein interaction networks. Autom. Exp. 2010, 2, 2. [Google Scholar] [CrossRef]
- Phung, T.T.; Sugamata, R.; Uno, K.; Aratani, Y.; Ozato, K.; Kawachi, S.; Thanh Nguyen, L.; Nakayama, T.; Suzuki, K. Key role of regulated upon activation normal T-cell expressed and secreted, nonstructural protein1 and myeloperoxidase in cytokine storm induced by influenza virus PR-8 (A/H1N1) infection in A549 bronchial epithelial cells. Microbiol. Immunol. 2011, 55, 874–884. [Google Scholar] [CrossRef]
- Dergham, M.; Lepers, C.; Verdin, A.; Billet, S.; Cazier, F.; Courcot, D.; Shirali, P.; Garcon, G. Prooxidant and proinflammatory potency of air pollution particulate matter (PM(2).(5)(-)(0).(3)) produced in rural, urban, or industrial surroundings in human bronchial epithelial cells (BEAS-2B). Chem. Res. Toxicol. 2012, 25, 904–919. [Google Scholar] [CrossRef]
- Koarai, A.; Sugiura, H.; Yanagisawa, S.; Ichikawa, T.; Minakata, Y.; Matsunaga, K.; Hirano, T.; Akamatsu, K.; Ichinose, M. Oxidative stress enhances toll-like receptor 3 response to double-stranded RNA in airway epithelial cells. Am. J. Respir. Cell Mol. Biol. 2010, 42, 651–660. [Google Scholar] [CrossRef]
- Alfaro-Moreno, E.; Torres, V.; Miranda, J.; Martinez, L.; Garcia-Cuellar, C.; Nawrot, T.S.; Vanaudenaerde, B.; Hoet, P.; Ramirez-Lopez, P.; Rosas, I.; et al. Induction of IL-6 and inhibition of IL-8 secretion in the human airway cell line Calu-3 by urban particulate matter collected with a modified method of PM sampling. Environ. Res. 2009, 109, 528–535. [Google Scholar] [CrossRef]
- Veranth, J.M.; Moss, T.A.; Chow, J.C.; Labban, R.; Nichols, W.K.; Walton, J.C.; Watson, J.G.; Yost, G.S. Correlation of in vitro cytokine responses with the chemical composition of soil-derived particulate matter. Environ. Health Perspect. 2006, 114, 341–349. [Google Scholar]
- Baulig, A.; Sourdeval, M.; Meyer, M.; Marano, F.; Baeza-Squiban, A. Biological effects of atmospheric particles on human bronchial epithelial cells. Comparison with diesel exhaust particles. Toxicol. In Vitro 2003, 17, 567–573. [Google Scholar] [CrossRef]
- Reibman, J.; Hsu, Y.; Chen, L.C.; Kumar, A.; Su, W.C.; Choy, W.; Talbot, A.; Gordon, T. Size fractions of ambient particulate matter induce granulocyte macrophage colony-stimulating factor in human bronchial epithelial cells by mitogen-activated protein kinase pathways. Am. J. Respir. Cell Mol. Biol. 2002, 27, 455–462. [Google Scholar]
- Liu, X.; Togo, S.; Al-Mugotir, M.; Kim, H.; Fang, Q.; Kobayashi, T.; Wang, X.; Mao, L.; Bitterman, P.; Rennard, S. NF-kappaB mediates the survival of human bronchial epithelial cells exposed to cigarette smoke extract. Respir. Res. 2008, 9, 66. [Google Scholar] [CrossRef]
- Rennolds, J.; Malireddy, S.; Hassan, F.; Tridandapani, S.; Parinandi, N.; Boyaka, P.N.; Cormet-Boyaka, E. Curcumin regulates airway epithelial cell cytokine responses to the pollutant cadmium. Biochem. Biophys. Res. Commun. 2012, 417, 256–261. [Google Scholar] [CrossRef]
- Turner, M.C.; Krewski, D.; Pope, C.A., 3rd; Chen, Y.; Gapstur, S.M.; Thun, M.J. Long-term ambient fine particulate matter air pollution and lung cancer in a large cohort of never-smokers. Am. J. Respir. Crit. Care Med. 2011, 184, 1374–1381. [Google Scholar] [CrossRef]
- Pope, C.A., 3rd.; Burnett, R.T.; Thun, M.J.; Calle, E.E.; Krewski, D.; Ito, K.; Thurston, G.D. Lung cancer, cardiopulmonary mortality, and long-term exposure to fine particulate air pollution. JAMA 2002, 287, 1132–1141. [Google Scholar] [CrossRef]
- Chen, H.W.; Lee, J.Y.; Huang, J.Y.; Wang, C.C.; Chen, W.J.; Su, S.F.; Huang, C.W.; Ho, C.C.; Chen, J.J.; Tsai, M.F.; et al. Curcumin inhibits lung cancer cell invasion and metastasis through the tumor suppressor HLJ1. Cancer Res. 2008, 68, 7428–7438. [Google Scholar]
- Lin, S.S.; Lai, K.C.; Hsu, S.C.; Yang, J.S.; Kuo, C.L.; Lin, J.P.; Ma, Y.S.; Wu, C.C.; Chung, J.G. Curcumin inhibits the migration and invasion of human A549 lung cancer cells through the inhibition of matrix metalloproteinase-2 and -9 and Vascular Endothelial Growth Factor (VEGF). Cancer Res. 2009, 285, 127–133. [Google Scholar]
- Alexandrow, M.G.; Song, L.J.; Altiok, S.; Gray, J.; Haura, E.B.; Kumar, N.B. Curcumin: A novel Stat3 pathway inhibitor for chemoprevention of lung cancer. Eur. J. Cancer Prev. 2012, 21, 407–412. [Google Scholar] [CrossRef]
- Gescher, A.J.; Sharma, R.A.; Steward, W.P. Cancer chemoprevention by dietary constituents: a tale of failure and promise. Lancet Oncol. 2001, 2, 371–379. [Google Scholar] [CrossRef]
- Agopyan, N.; Head, J.; Yu, S.; Simon, S.A. TRPV1 receptors mediate particulate matter-induced apoptosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 286, L563–l572. [Google Scholar] [CrossRef]
- Nel, A.E.; Diaz-Sanchez, D.; Li, N. The role of particulate pollutants in pulmonary inflammation and asthma: Evidence for the involvement of organic chemicals and oxidative stress. Curr. Opin. Pulm. Med. 2001, 7, 20–26. [Google Scholar] [CrossRef]
- Gualtieri, M.; Ovrevik, J.; Mollerup, S.; Asare, N.; Longhin, E.; Dahlman, H.J.; Camatini, M.; Holme, J.A. Airborne urban particles (Milan winter-PM2.5) cause mitotic arrest and cell death: Effects on DNA, mitochondria, AhR binding and spindle organization. Mutat. Res. 2011, 713, 18–31. [Google Scholar] [CrossRef]
- Zhang, J.; Ghio, A.J.; Gao, M.; Wei, K.; Rosen, G.D.; Upadhyay, D. Ambient particulate matter induces alveolar epithelial cell cycle arrest: role of G1 cyclins. FEBS Lett. 2007, 581, 5315–5320. [Google Scholar] [CrossRef]
- Soberanes, S.; Panduri, V.; Mutlu, G.M.; Ghio, A.; Bundinger, G.R.; Kamp, D.W. p53 mediates particulate matter-induced alveolar epithelial cell mitochondria-regulated apoptosis. Am. J. Respir. Crit. Care Med. 2006, 174, 1229–1238. [Google Scholar] [CrossRef]
- Dagher, Z.; Garcon, G.; Billet, S.; Gosset, P.; Ledoux, F.; Courcot, D.; Aboukais, A.; Shirali, P. Activation of different pathways of apoptosis by air pollution particulate matter (PM2.5) in human epithelial lung cells (L132) in culture. Toxicology 2006, 225, 12–24. [Google Scholar] [CrossRef]
- Siddiqui, M.A.; Ahamed, M.; Ahmad, J.; Majeed Khan, M.A.; Musarrat, J.; Al-Khedhairy, A.A.; Alrokayan, S.A. Nickel oxide nanoparticles induce cytotoxicity, oxidative stress and apoptosis in cultured human cells that is abrogated by the dietary antioxidant curcumin. Food Chem. Toxicol. 2012, 50, 641–647. [Google Scholar] [CrossRef]
- Ramachandran, C.; You, W. Differential sensitivity of human mammary epithelial and breast carcinoma cell lines to curcumin. Breast Cancer Res. Treat. 1999, 54, 269–278. [Google Scholar] [CrossRef]
- Pujolsa, L.; Mullol, J.; Picado, C. Glucocorticoid receptor in human respiratory epithelial cells. Neuroimmunomodulation 2009, 16, 290–299. [Google Scholar] [CrossRef]
- Verheggen, M.M.; Adriaansen-Soeting, P.W.; Berrevoets, C.A.; van Hal, P.T.; Brinkmann, A.O.; Hoogsteden, H.C.; Versnel, M.A. Glucocorticoid receptor expression in human bronchial epithelial cells: effects of smoking and COPD. Mediat. Inflamm. 1998, 7, 275–281. [Google Scholar] [CrossRef]
- Barnes, P.J. Histone deacetylase-2 and airway disease. Ther. Adv. Respir. Dis. 2009, 3, 235–243. [Google Scholar] [CrossRef]
- Marwick, J.A.; Ito, K.; Adcock, I.M.; Kirkham, P.A. Oxidative stress and steroid resistance in asthma and COPD: pharmacological manipulation of HDAC-2 as a therapeutic strategy. Expert Opin. Ther. Tar. 2007, 11, 745–755. [Google Scholar] [CrossRef]
- Ito, K.; Caramori, G.; Adcock, I.M. Therapeutic potential of phosphatidylinositol 3-kinase inhibitors in inflammatory respiratory disease. J. Pharmacol. Exp. Ther. 2007, 321, 1–8. [Google Scholar] [CrossRef]
- Barnes, P.J. Targeting the epigenome in the treatment of asthma and chronic obstructive pulmonary disease. Proc. Am. Thorac. Soc. 2009, 6, 693–696. [Google Scholar] [CrossRef]
- Watterson, T.L.; Hamilton, B.; Martin, R.S.; Coulombe, R.A., Jr. Urban particulate matter activates Akt in human lung cells. Arch. Toxicol. 2012, 86, 121–135. [Google Scholar] [CrossRef]
- Syed, D.N.; Afaq, F.; Kweon, M.H.; Hadi, N.; Bhatia, N.; Spiegelman, V.S.; Mukhtar, H. Green tea polyphenol EGCG suppresses cigarette smoke condensate-induced NF-kappaB activation in normal human bronchial epithelial cells. Oncogene 2007, 26, 673–682. [Google Scholar] [CrossRef]
- Yu, H.; Li, Q.; Kolosov, V.P.; Perelman, J.M.; Zhou, X. Regulation of cigarette smoke-induced mucin expression by neuregulin1beta/ErbB3 signalling in human airway epithelial cells. Basic Clin. Pharmacol. Toxicol. 2011, 109, 63–72. [Google Scholar] [CrossRef]
- Moriyuki, K.; Sekiguchi, F.; Matsubara, K.; Nishikawa, H.; Kawabata, A. Curcumin Inhibits the proteinase-activated receptor-2-triggered prostaglandin E2 production by suppressing cyclooxygenase-2 upregulation and Akt-dependent activation of nuclear factor-kappaB in human lung epithelial cells. J. Pharmacol. Sci. 2010, 114, 225–229. [Google Scholar] [CrossRef]
- Lee, C.W.; Lin, C.C.; Lin, W.N.; Liang, K.C.; Luo, S.F.; Wu, C.B.; Wang, S.W.; Yang, C.M. TNF-alpha induces MMP-9 expression via activation of Src/EGFR, PDGFR/PI3K/Akt cascade and promotion of NF-kappaB/p300 binding in human tracheal smooth muscle cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L799–l812. [Google Scholar]
- Mantecca, P.; Farina, F.; Moschini, E.; Gallinotti, D.; Gualtieri, M.; Rohr, A.; Sancini, G.; Palestini, P.; Camatini, M. Comparative acute lung inflammation induced by atmospheric PM and size-fractionated tire particles. Toxicol. Lett. 2010, 198, 244–254. [Google Scholar] [CrossRef]
- Dagher, Z.; Garcon, G.; Billet, S.; Verdin, A.; Ledoux, F.; Courcot, D.; Aboukais, A.; Shirali, P. Role of nuclear factor-kappa B activation in the adverse effects induced by air pollution particulate matter (PM2.5) in human epithelial lung cells (L132) in culture. J. Appl. Toxicol. 2007, 27, 284–290. [Google Scholar] [CrossRef]
- Zhao, Y.; Usatyuk, P.V.; Gorshkova, I.A.; He, D.; Wang, T.; Moreno-Vinasco, L.; Geyh, A.S.; Breysse, P.N.; Samet, J.M.; Spannhake, E.W.; et al. Regulation of COX-2 expression and IL-6 release by particulate matter in airway epithelial cells. Am. J. Respir. Cell Mol. Biol. 2009, 40, 19–30. [Google Scholar]
- Freeman, A.K.; Morrison, D.K. 14–3-3 Proteins: diverse functions in cell proliferation and cancer progression. Semin. Cell Dev. Biol. 2011, 22, 681–687. [Google Scholar] [CrossRef]
- Gardino, A.K.; Yaffe, M.B. 14–3-3 proteins as signaling integration points for cell cycle control and apoptosis. Semin. Cell Dev. Biol. 2011, 22, 688–695. [Google Scholar] [CrossRef]
- Qi, W.; Liu, X.; Chen, W.; Li, Q.; Martinez, J.D. Overexpression of 14–3-3gamma causes polyploidization in H322 lung cancer cells. Mol. Carcinog. 2007, 46, 847–856. [Google Scholar] [CrossRef]
- Kawamoto, S.; Iemura, N.; Inoue, Y.; Katakura, Y.; Shirahata, S. Effect of 14–3-3 protein induction on cell proliferation of A549 human lung adenocarcinoma. Cytotechnology 2000, 33, 253–257. [Google Scholar]
- Wang, J.B.; Qi, L.L.; Zheng, S.D.; Wang, H.Z.; Wu, T.X. Curcumin suppresses PPARdelta expression and related genes in HT-29 cells. World J. Gastroenterol. 2009, 15, 1346–1352. [Google Scholar] [CrossRef]
- Miyata, R.; van Eeden, S.F. The innate and adaptive immune response induced by alveolar macrophages exposed to ambient particulate matter. Toxicol. Appl. Pharmacol. 2011, 257, 209–226. [Google Scholar] [CrossRef]
- GSE7010. Available online: http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE7010 (accessed on 16 October 2012).
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef]
- Huang, Y.C.; Karoly, E.D.; Dailey, L.A.; Schmitt, M.T.; Silbajoris, R.; Graff, D.W.; Devlin, R.B. Comparison of gene expression profiles induced by coarse, fine, and ultrafine particulate matter. J. Toxicol. Environ. Health A 2011, 74, 296–312. [Google Scholar]
- Wang, Y.; Xiao, J.; Suzek, T.O.; Zhang, J.; Wang, J.; Bryant, S.H. PubChem: A public information system for analyzing bioactivities of small molecules. Nucleic acids Res. 2009, 37, W623–w633. [Google Scholar]
- Canny, S.A.; Cruz, Y.; Southern, M.R.; Griffin, P.R. PubChem promiscuity: A web resource for gathering compound promiscuity data from PubChem. Bioinformatics 2012, 28, 140–141. [Google Scholar] [CrossRef]
- PubChem Promiscuity. Available online: http://chemutils.florida.scripps.edu:8080/pcpromiscuity (Accessed on 16 October 2012).
- Real-Chicharro, A.; Ruiz-Mostazo, I.; Navas-Delgado, I.; Kerzazi, A.; Chniber, O.; Sanchez-Jimenez, F.; Medina, M.A.; Aldana-Montes, J.F. Protopia: A protein-protein interaction tool. BMC Bioinformatics 2009, 10 Suppl. 12, S17. [Google Scholar]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Martin, A.; Ochagavia, M.E.; Rabasa, L.C.; Miranda, J.; Fernandez-de-Cossio, J.; Bringas, R. BisoGenet: A new tool for gene network building, visualization and analysis. BMC Bioinformatics 2010, 11, 91. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, Z.; Niu, X.; Lu, C.; Jiang, M.; Xiao, G.G.; Lu, A. The Effect of Curcumin on Human Bronchial Epithelial Cells Exposed to Fine Particulate Matter: A Predictive Analysis. Molecules 2012, 17, 12406-12426. https://doi.org/10.3390/molecules171012406
Zhang Z, Niu X, Lu C, Jiang M, Xiao GG, Lu A. The Effect of Curcumin on Human Bronchial Epithelial Cells Exposed to Fine Particulate Matter: A Predictive Analysis. Molecules. 2012; 17(10):12406-12426. https://doi.org/10.3390/molecules171012406
Chicago/Turabian StyleZhang, Zhiguo, Xuyan Niu, Cheng Lu, Miao Jiang, Gary G. Xiao, and Aiping Lu. 2012. "The Effect of Curcumin on Human Bronchial Epithelial Cells Exposed to Fine Particulate Matter: A Predictive Analysis" Molecules 17, no. 10: 12406-12426. https://doi.org/10.3390/molecules171012406
APA StyleZhang, Z., Niu, X., Lu, C., Jiang, M., Xiao, G. G., & Lu, A. (2012). The Effect of Curcumin on Human Bronchial Epithelial Cells Exposed to Fine Particulate Matter: A Predictive Analysis. Molecules, 17(10), 12406-12426. https://doi.org/10.3390/molecules171012406