Selection of a Mimotope Peptide of S-adenosyl-l-homocysteine and Its Application in Immunoassays

Abstract
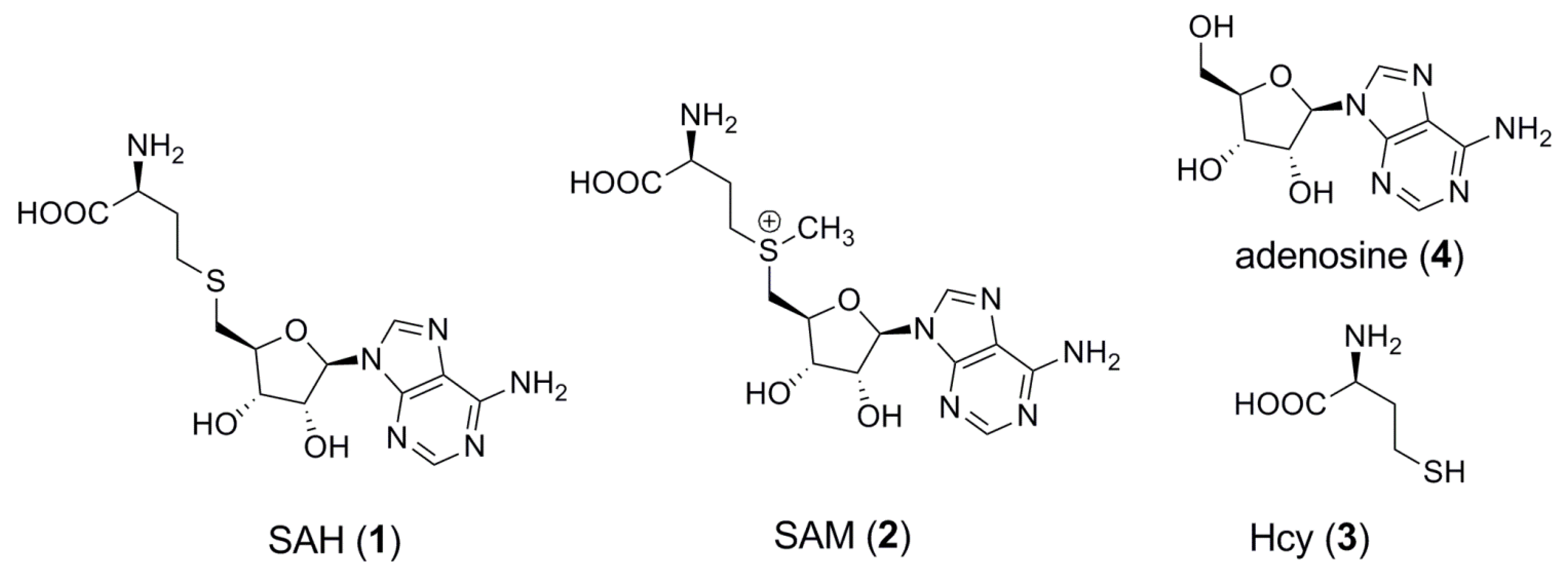
:1. Introduction

2. Results and Discussion
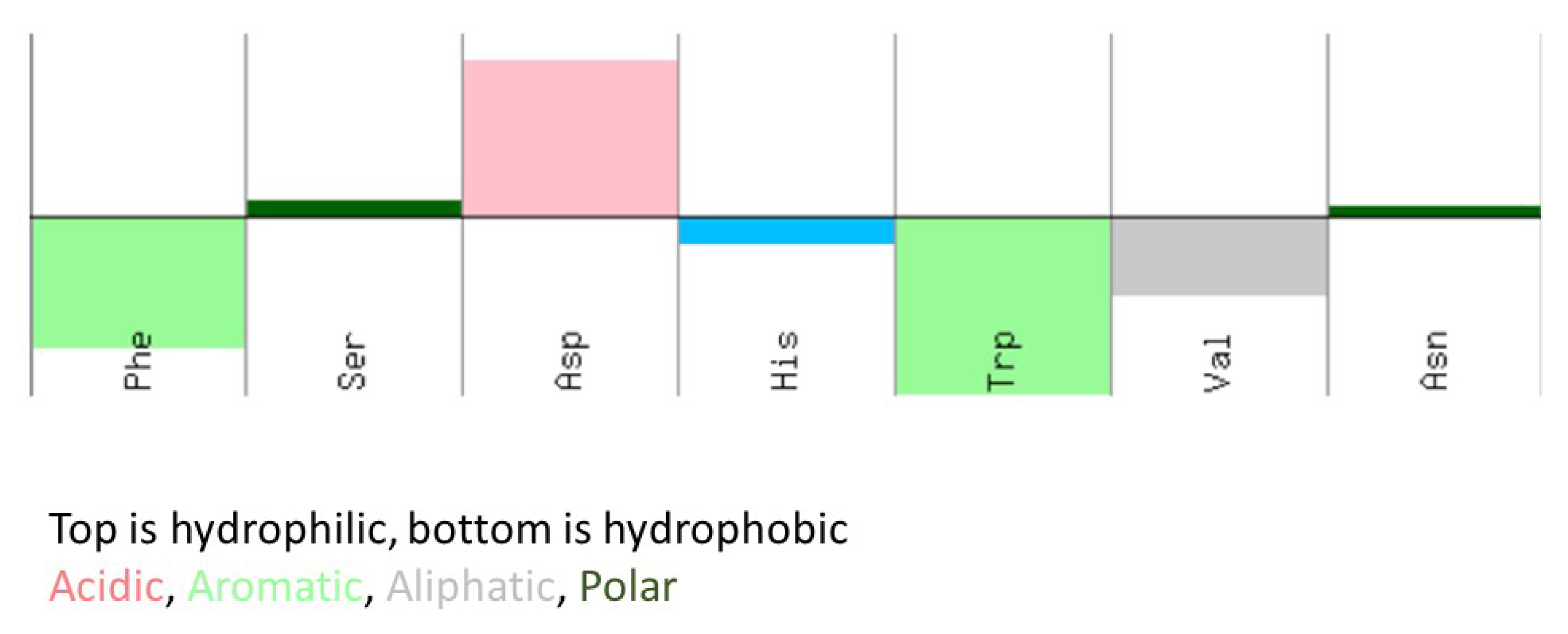
2.1. Panning-Elution Selection of Specific Phage

{kind=link}
{kind=link}
{kind=link}
| Ph.D-7 | Ph.D.-C7C |
|---|---|
| FSDHWVN | CVQMPAHSC |
| FSDHWVN | CPNSTHRNC |
| FSDHWVN | CMHTHSRMC |
| FSDHWVN | CNTGSPYEC |
| FSDHWVN | CFSGMSTDC |
| FSDHWVN | CDASRPATC |
| -- | CRGATPMSC |
| -- | CSEGLLNTC |
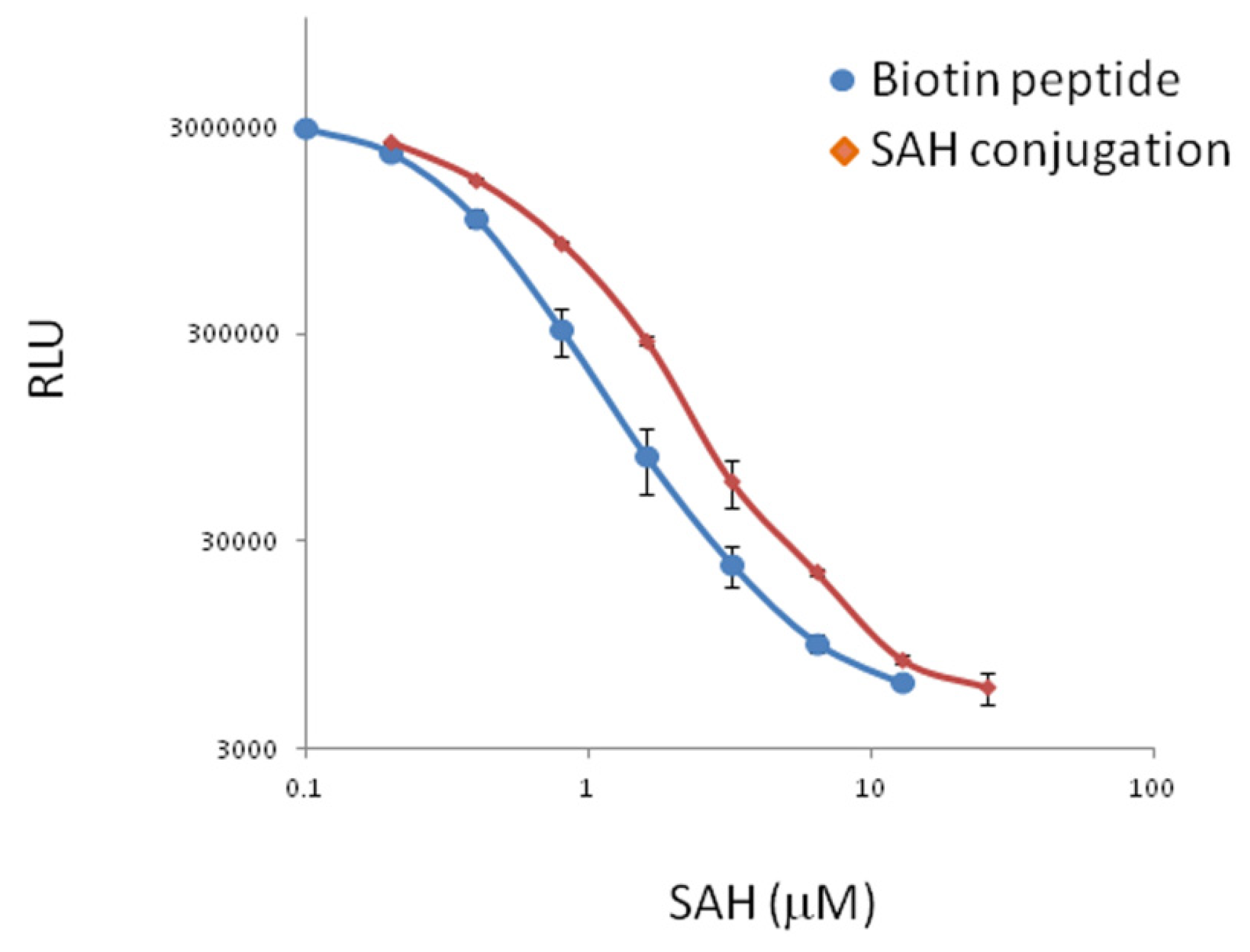
2.2. Use of the Discovered Mimotope Peptide in Immunoassay of SAH


3. Experimental
3.1. Materials
3.2. Selection of anti-SAH MAb Binding Peptides from Phage Library
3.3. Use of the Mimotope Peptide for Immunoassay of SAH
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Melnyk, S.; Pogribna, M.; Pogribny, I.P.; Yi, P.; James, S.J. Measurement of plasma and intracellular S-adenosylmethionine and S-adenosylhomocysteine utilizing coulometric electrochemical detection: Alterations with plasma homocysteine and pyridoxal 5'-phosphate concentrations. Clin. Chem. 2000, 46, 265–272. [Google Scholar]
- Poirier, L.A.; Wise, C.K.; Delongchamp, R.R.; Sinha, R. Blood determinations of S-adenosylmethionine, S-adenosylhomocysteine, and homocysteine: Correlations with diet. Cancer Epidemiol. Biomarkers Prev. 2001, 10, 649–655. [Google Scholar]
- Obeid, R.; Schadt, A.; Dillmann, U.; Kostopoulos, P.; Fassbender, K.; Herrmann, W. Methylation Status and Neurodegenerative Markers in Parkinson Disease. Clin. Chem. 2009, 55, 1852–1860. [Google Scholar] [CrossRef]
- Frantzen, F.; Faaren, A.L.; Alfheim, I.; Nordhei, A.K. Enzyme conversion immunoassay for determining total homocysteine in plasma or serum. Clin. Chem. 1998, 44, 311–316. [Google Scholar]
- Chun, B.-G.; Paik, W.K.; Kim, S. Simple separation of adenine and adenosyl-sulfur compounds by high performance liquid chromatography. J. Chromatogr. 1983, 264, 301–328. [Google Scholar]
- Phalipon, A.; Folgori, A.; Arondel, J.; Sgaramella, G.; Fortugno, P.; Cortese, R.; Sansonetti, P.J.; Felici, F. Induction of anti-carbohydrate antibodies by phage library-selected peptide mimics. Eur. J. Immunol. 1997, 27, 2620–2625. [Google Scholar] [CrossRef]
- Bottger, V.; Peters, L.E.; Micheel, B. Identification of peptide mimotopes for the fluorescein hapten binding of monoclonal antibody B13-DE1. J. Mol. Recognit. 1999, 12, 191–197. [Google Scholar] [CrossRef]
- Yuan, Q.P.; Pestka, J.J.; Hespenheide, B.M.; Kuhn, L.A.; Linz, J.E.; Hart, L.P. Identification of mimotope peptides which bind to the mycotoxin deoxynivalenol-specific monoclonal antibody. Appl. Environ. Microbiol. 1999, 65, 3279–3286. [Google Scholar]
- Wang, Y.R.; Wang, H.; Li, P.W.; Zhang, Q.; Kim, H.J.; Gee, S.J.; Hammock, B.D. Phage-displayed peptide that mimics aflatoxins and its application in immunoassay. J. Agric. Food. Chem. 2013, 61, 2426–2433. [Google Scholar]
- Schmidt, T.G.M.; Koepke, J.; Frank, R.; Skerra, A. Molecular interaction between the Strep-tag affinity peptide and its cognate target, streptavidin. J. Mol. Biol. 1996, 255, 753–766. [Google Scholar] [CrossRef]
- Verdolino, V.; Cammi, R.; Munk, B.H.; Schlegel, H.B. Calculation of pK values of nucleobases and the guanine oxidation products guanidinohydantoin and spiroiminodihydantoin using density functional theory and a polarizable continuum model. J. Phys. Chem. B 2008, 112, 16860–16873. [Google Scholar]
- Sample Availability: Samples of materials and compounds that are not commercially available can be obtained from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, C.; Tzertzinis, G. Selection of a Mimotope Peptide of S-adenosyl-l-homocysteine and Its Application in Immunoassays. Molecules 2013, 18, 13020-13026. https://doi.org/10.3390/molecules181013020
Wu C, Tzertzinis G. Selection of a Mimotope Peptide of S-adenosyl-l-homocysteine and Its Application in Immunoassays. Molecules. 2013; 18(10):13020-13026. https://doi.org/10.3390/molecules181013020
Chicago/Turabian StyleWu, Chun, and George Tzertzinis. 2013. "Selection of a Mimotope Peptide of S-adenosyl-l-homocysteine and Its Application in Immunoassays" Molecules 18, no. 10: 13020-13026. https://doi.org/10.3390/molecules181013020
APA StyleWu, C., & Tzertzinis, G. (2013). Selection of a Mimotope Peptide of S-adenosyl-l-homocysteine and Its Application in Immunoassays. Molecules, 18(10), 13020-13026. https://doi.org/10.3390/molecules181013020