Chemistry and Antiviral Activity of Arrabidaea pulchra (Bignoniaceae)

Abstract
:1. Introduction
2. Results




{kind=link}
{kind=link}
{kind=link}
{kind=link}
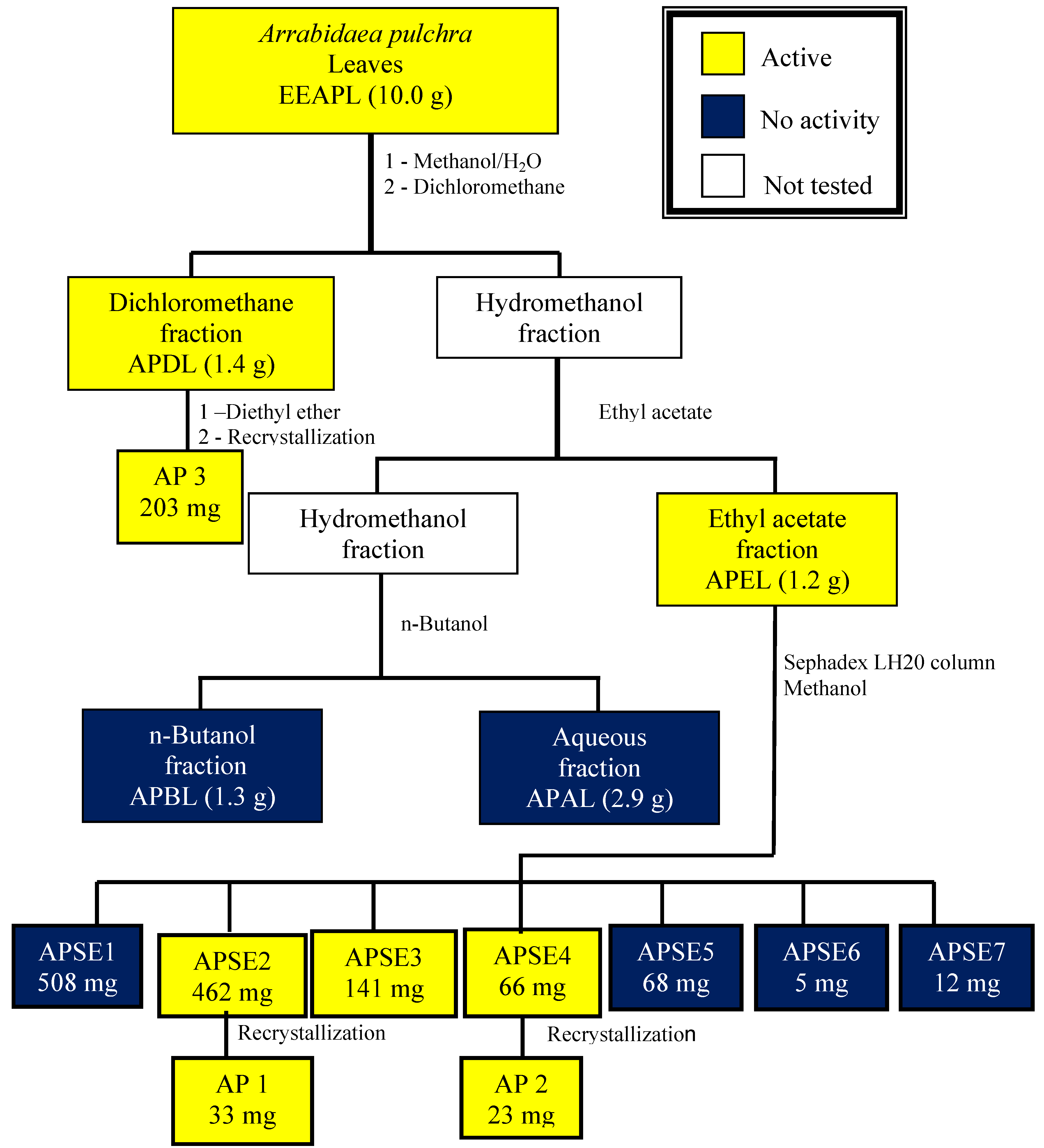{kind=link}
| Extract Fractions Compounds | Vero cells CC50µg/mL | LLCMK2 cells CC50µg/mL | a HSV-1 EC50 µg/mL | SI | b VACV-WR EC50 µg/mL | SI | c EMCV EC50 µg/mL | d DENV-2 EC50 µg/mL | SI |
|---|---|---|---|---|---|---|---|---|---|
| EEAPL | >500 | 124.4 ± 0.8 | 235.3 ± 9.7 | >2.1 | 245.2 ± 13.4 | >2.0 | NA | 46.8 ± 1.6 | 2.7 |
| APDL | 25.0 ± 0.3 | 19.6 ± 1.7 | NA | 18.6 ± 0.9 | 1.3 | NA | 15.4 ± 2.1 | 1.3 | |
| APEL | >200 | >200 | 121.9 ± 9.8 | >1.6 | 18.4 ± 1.9 | >10.9 | NA | 12.2 ± 1.6 | >16.3 |
| APSE2 | >200 | >200 | NA | 16.7 ± 1.8 | >12.0 | NA | 10.6 ± 2.1 | >18.9 | |
| APSE3 | >200 | >200 | NA | 18.1 ± 3.4 | >11.0 | NA | 11.9 ± 1.5 | >16.8 | |
| APSE4 | >200 | >200 | NA | 15.9 ± 2.3 | >12.3 | NA | 8.8 ± 0.7 | >22.7 | |
| AP 1 | >200 | 12.9 ± 1.1 | NA | NA | NA | 3.4 ± 0.4 | 3.8 | ||
| AP 2 | >200 | 56.1 ± 2.4 | NA | NA | NA | 2.8 ± 0.4 | 20.0 | ||
| AP 3 | 8.1 ± 0.9 | 9.9 ± 1.3 | 6.2 ± 0.1 | 1.6 | NA | NA | 3.2 + 0.6 | 3.1 | |
| Acyclovir | a 40 | ||||||||
| α-2a Interferon | ef 2.5 × 102 | ef 1.5 × 102 | ef 2.5 × 103 |

3. Discussion
4. Experimental
4.1. Plant Material
4.2. Extraction and Isolation

4.3. Structural Determination
4.4. Spectral Data
4.5. HPLC Analyses
4.6. Cell Culture and Virus
4.7. Cytotoxicity Assay
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Chattopadhyay, D.; Naik, T.N. Antivirals of ethnomedicinal origin: Structure-activity relationship and scope. Mini Rev. Med. Chem. 2007, 7, 275–301. [Google Scholar] [CrossRef]
- Mukhtar, M.; Arshad, M.; Ahmad, M.; Pomerantz, R.J.; Wigdahl, B.; Parveen, Z. Antiviral potentials of medicinal plants. Virus Res. 2008, 131, 111–120. [Google Scholar] [CrossRef]
- Selisco, B.; Guillemot, J.C.; Alvarez, K.; Canard, B. Opportunities in the Development of Anti-Dengue Drugs. Available online: http://www.tropika.net/svc/review/061001-Dengue_Drugs (accessed on 22 July 2013).
- Lohmann, L.G. Untangling the phylogeny of neotropical lianas (Bignonieae, Bignoniaceae). Am. J. Bot. 2006, 93, 304–318. [Google Scholar] [CrossRef]
- Gentry, A.H. A synopsis of Bignoniaceae ethnobotany and economic botany. Ann. Mo. Bot. Gard. 1992, 79, 53–64. [Google Scholar] [CrossRef]
- Fischer, E.; Theisen, I.; Lohmann, G.L. Bignoniaceae. In The Families and Genera of Vascular Plants; Kubitsky, K., Ed.; Springer-Verlag: Heidelberg, Germany, 2004; Volume VII, pp. 9–38. [Google Scholar]
- Scudeller, V.V.; Carvalho-Okano, R.M. A tribo Bignonieae Spreng (Bignoniaceae) no Parque Estadual do Rio Doce (MG). Iheringia Série Botânica 1998, 51, 79–133. [Google Scholar]
- Alvarenga, T.A.; Bêdo, T.R.O.; Braguine, C.G.; Gonçalves, U.O.; Magalhães, L.G.; Rodrigues, V.; Gimenez, V.M.M.; Groppo, M.; Silva, M.L.A.; Cunha, W.R.; Januário, A.H.; Pauletti, P.M. Evaluation of Cuspidaria pulchra and its isolated compounds against Schistosoma mansoni Adult Worms. Int. J. Biotechn. Wellness Ind. 2012, 1, 122–127. [Google Scholar]
- Brandão, G.C.; Kroon, E.G.; Santos, J.R.; Stehmann, J.R.; Lombardi, J.A.; Oliveira, A.B. Antiviral activities of plants occurring in the state of Minas Gerais, Brazil. Part 2. Screening Bignoniaceae species. Braz. J. Pharmacogn. 2010, 20, 742–750. [Google Scholar]
- Brandão, G.C.; Kroon, E.G.; Santos, J.R.; Stehmann, J.R.; Lombardi, J.A; Oliveira, A.B. Antiviral activity of Bignoniaceae species occurring in the State of Minas Gerais (Brazil): Part 1. Letters Appl. Microbiol. 2010, 51, 469–476. [Google Scholar] [CrossRef]
- Lima, C.S.A.; Amorim, E.L.C.; Sena, K.X.F.R.; Chiappeta, A.A.; Nunes, X.P.; Agra, M.F; Cunha, E.V.L.; Silva, M.S.; Barbosa-Filho, J.M. Antimicrobial activity of a mixture of two isomeric phenylpropanoid glycosides from Arrabidaea harleyi A.H. Gentry (Bignoniaceae). Braz. J. Pharm. Sci. 2003, 39, 77–81. [Google Scholar]
- Challice, J.S.; Williams, A.H. Phenolic compounds of the genus Pyrus. I. Occurrence of flavones and phenolic acid derivatives of 3,4-dihydroxybenzyl alcohol 4-glucoside in Pyrus calleryana. Phytochemistry 1968, 7, 119–130. [Google Scholar] [CrossRef]
- Zheng, Q.Y.; Li, P.P.; Jin, F.S.; Yao, C.; Zhang, G.H.; Zang, T.; Ai, X. Ursolic acid induces ER stress response to activate ASK1-JNK signaling and induce apoptosis in human bladder cancer T24 cells. Cell Signal. 2013, 25, 206–213. [Google Scholar] [CrossRef]
- Yang, H.; Dou, Q.P. Targeting Apoptosis Pathway with Natural Terpenoids: Implications for Treatment of Breast and Prostate Cancer. Curr. Drug Targets 2010, 11, 733–744. [Google Scholar] [CrossRef]
- Ghisalberti, E.L. Lantana camara L. Verbenaceae. Fitoterapia 2000, 71, 467–486. [Google Scholar] [CrossRef]
- Kernan, M.R.; Amarquaye, A.; Chen, J.L.; Chan, J.; Sesin, D.F.; Parkinson, N.; Ye, Z.; Barrett, M.; Bales, C.; Stoddart, C.A. Antiviral phenylpropanoid glycosides from the medicinal plant Markhamia lutea. J. Nat. Prod. 1998, 61, 564–570. [Google Scholar] [CrossRef]
- Martins, F.O.; Esteves, P.F.; Mendes, G.S.; Barbi, N.S.; Menezes, F.S.; Romanos, M.T. Verbascoside isolated from Lepechinia speciosa has inhibitory activity against HSV-1 and HSV-2 in vitro. Nat. Prod. Commun. 2009, 4, 1693–1696. [Google Scholar]
- Lui, J. Pharmacology of oleanolic acid and ursolic acid. J. Ethnopharmacol. 1995, 49, 57–68. [Google Scholar] [CrossRef]
- Simões, L.R.; Maciel, G.M.; Brandão, G.C.; Kroon, E.G.; Castilho, R.O.; Oliveira, A.B. Antiviral activity of Distictella elongata (Vahl) Urb. (Bignoniaceae), a potentially useful source of anti-dengue drugs from the state of Minas Gerais, Brazil. Lett. Appl. Microbiol. 2011, 23, 602–607. [Google Scholar]
- Sarkhail, A.; Monsef-Esfehani, H.R.; Amin, G.; Surmaghi, M.H.S.; Shafiee, A. Phytochemical Study of Phlomis olivieri Benth. and Phlomis persica Boiss. DARU 2006, 14, 115–121. [Google Scholar]
- Rodriguez, D.J.; Chulia, J.; Simões, C.M.O.; Amoros, M.; Mariotte, A.M.; Girre, L. Search for in vitro antiviral activity of a new isoflavonic glycoside from Ulex europaeus. Planta Med. 1990, 56, 59–62. [Google Scholar] [CrossRef]
- Betancur-Galvis, L.A.; Saez; Granados, J.H.; Salazar, A.; Ossa, J.E. Antitumor and antiviral activity of Colombian medicinal plants extracts. Mem. Inst. Oswaldo Cruz. 1999, 94, 531–535. [Google Scholar] [CrossRef]
- Twentyman, P.R.; Luscombe, M. A study of some variables in a tetrazolium dye (MTT) based assay for cell growth and chemosensitivity. Br. J. Cancer. 1987, 56, 279–285. [Google Scholar] [CrossRef]
- Oliveira, A.B.; Kroon, E.G.; Brandão, G.C. Composições farmacêuticas antivirais contendo extrato, frações e/ou compostos isolados de Arrabidaea pulchra e uso. Brazilian Patent PI BR 014110003098 (INPI/MG), 27 November 2011. [Google Scholar]
- Sample Availability: Samples of the compounds AP 1, AP 2 and AP 3 are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Brandão, G.C.; Kroon, E.G.; Souza, D.E.R.; Filho, J.D.S.; Oliveira, A.B. Chemistry and Antiviral Activity of Arrabidaea pulchra (Bignoniaceae). Molecules 2013, 18, 9919-9932. https://doi.org/10.3390/molecules18089919
Brandão GC, Kroon EG, Souza DER, Filho JDS, Oliveira AB. Chemistry and Antiviral Activity of Arrabidaea pulchra (Bignoniaceae). Molecules. 2013; 18(8):9919-9932. https://doi.org/10.3390/molecules18089919
Chicago/Turabian StyleBrandão, Geraldo Célio, Erna G. Kroon, Danielle E.R. Souza, José D. Souza Filho, and Alaíde Braga Oliveira. 2013. "Chemistry and Antiviral Activity of Arrabidaea pulchra (Bignoniaceae)" Molecules 18, no. 8: 9919-9932. https://doi.org/10.3390/molecules18089919
APA StyleBrandão, G. C., Kroon, E. G., Souza, D. E. R., Filho, J. D. S., & Oliveira, A. B. (2013). Chemistry and Antiviral Activity of Arrabidaea pulchra (Bignoniaceae). Molecules, 18(8), 9919-9932. https://doi.org/10.3390/molecules18089919