Silymarin Induces Expression of Pancreatic Nkx6.1 Transcription Factor and β-Cells Neogenesis in a Pancreatectomy Model

Abstract
:1. Introduction
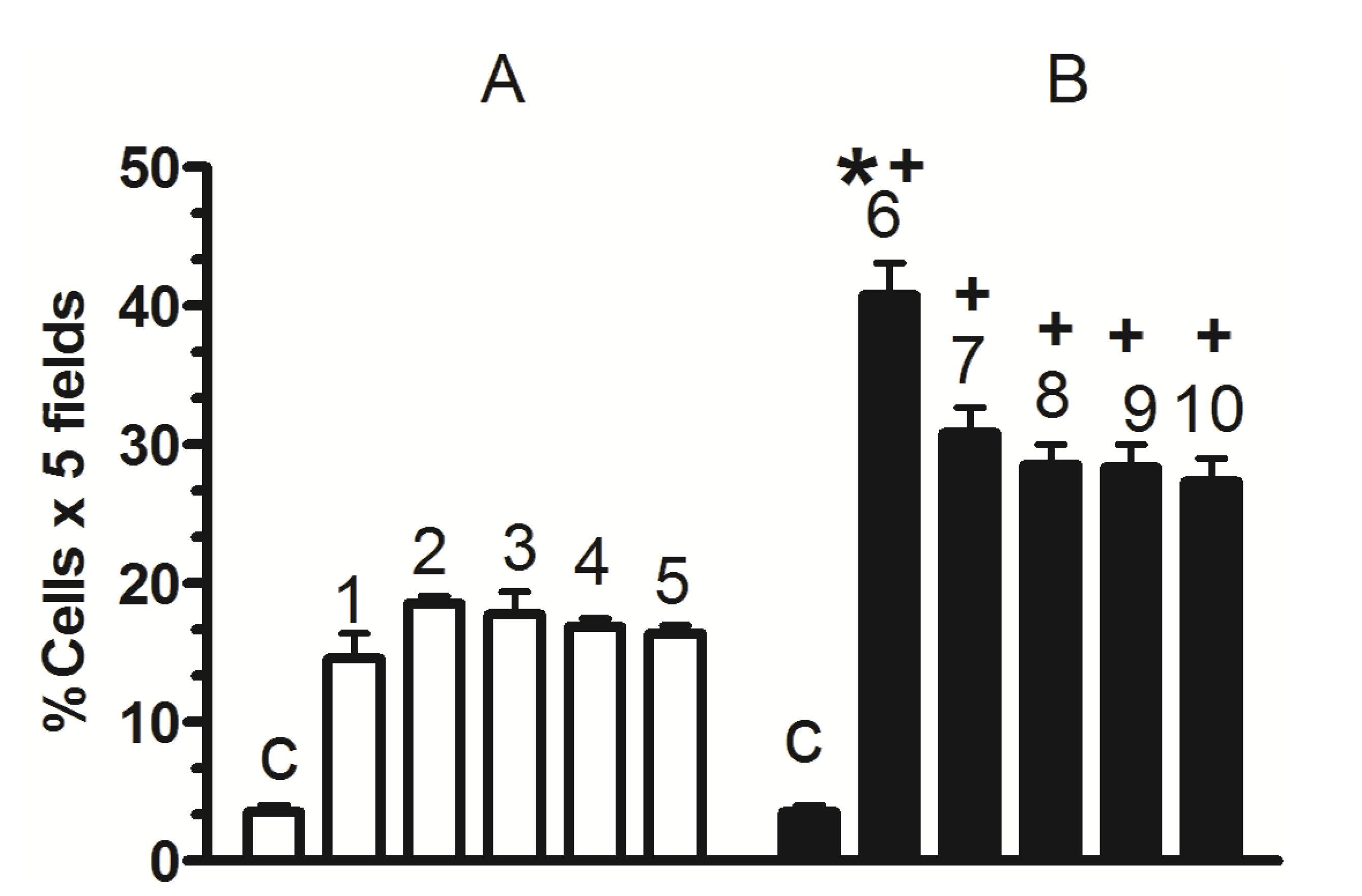
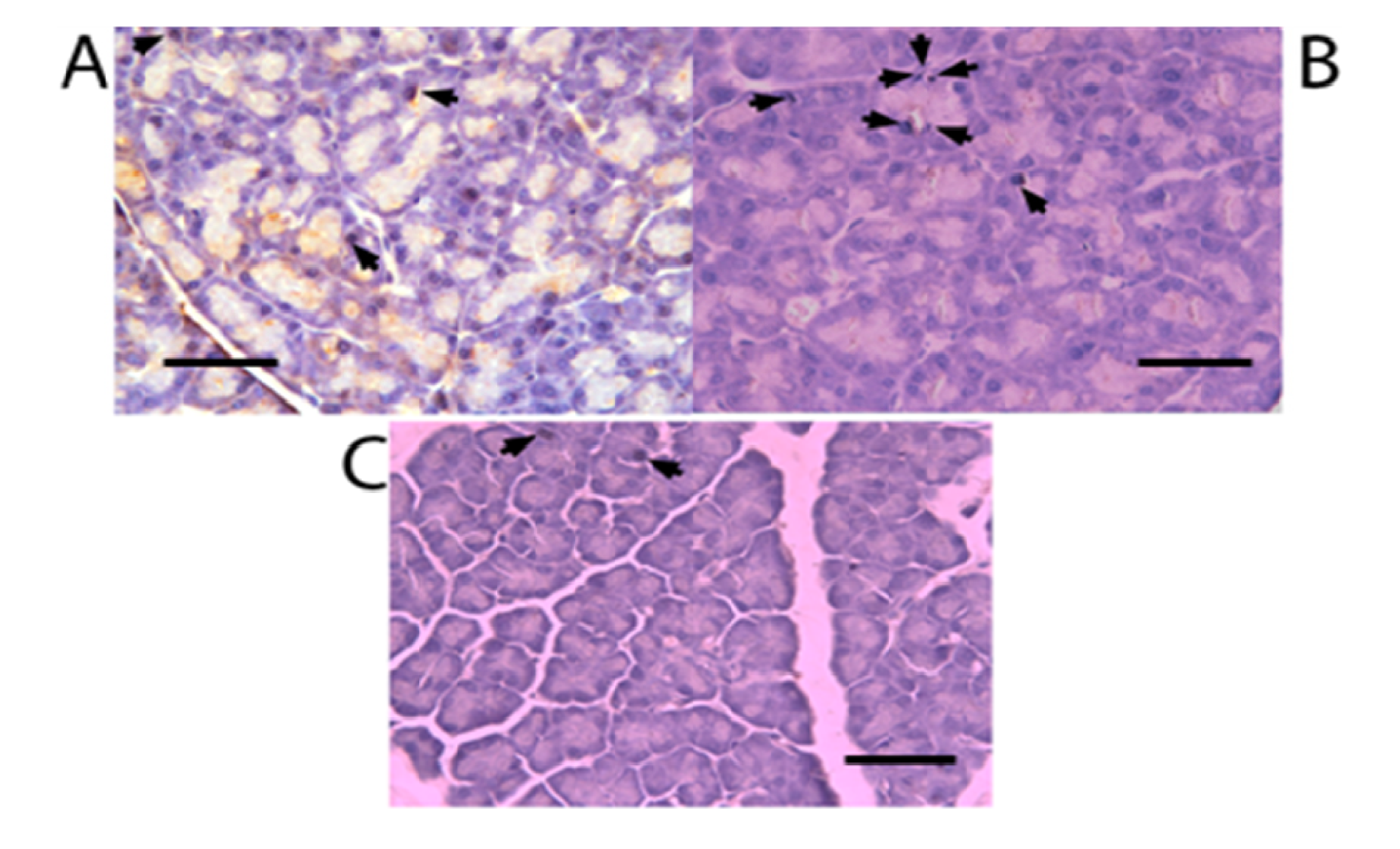
2. Results and Discussion
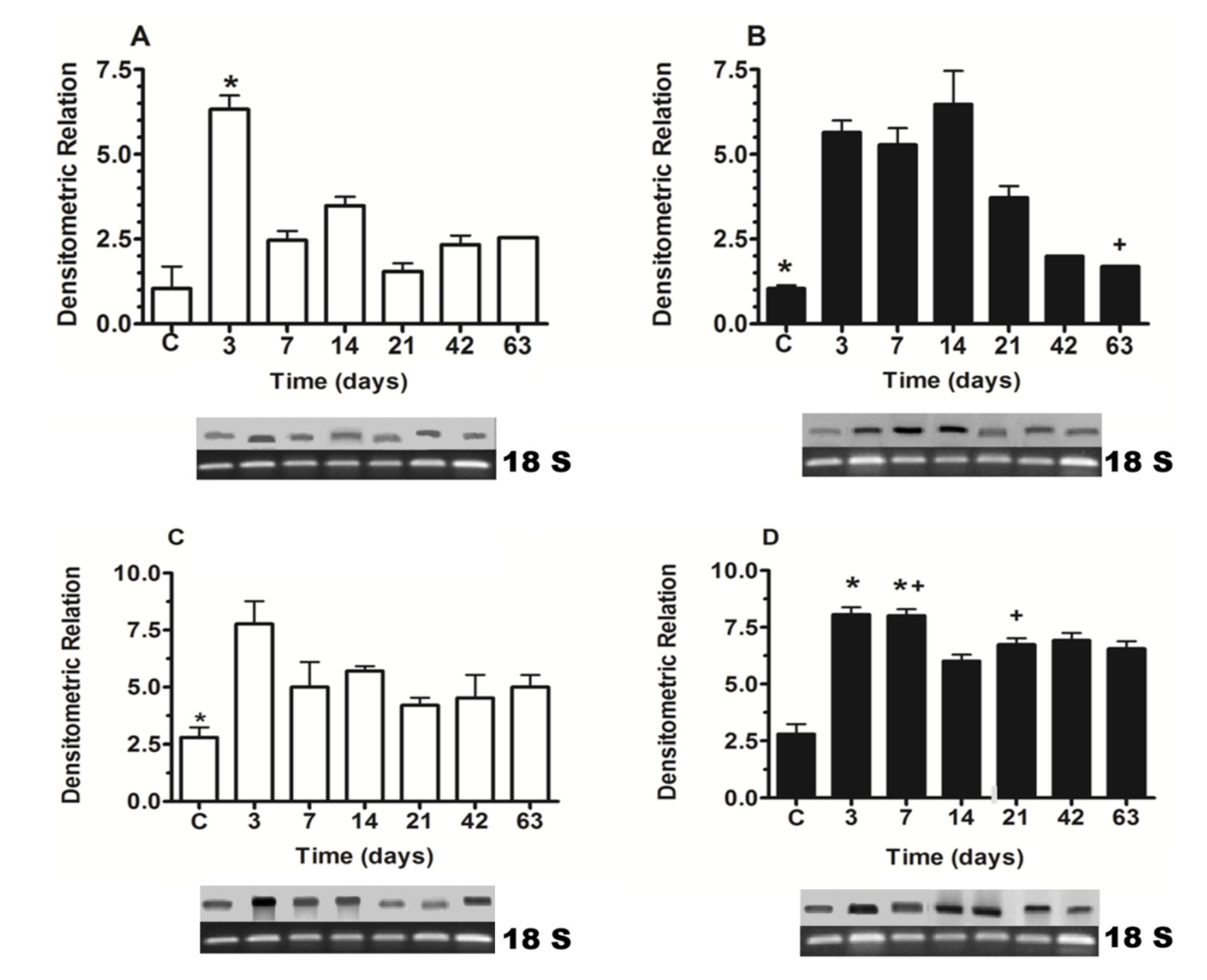
2.1. Nkx6.1 and Insulin Expression

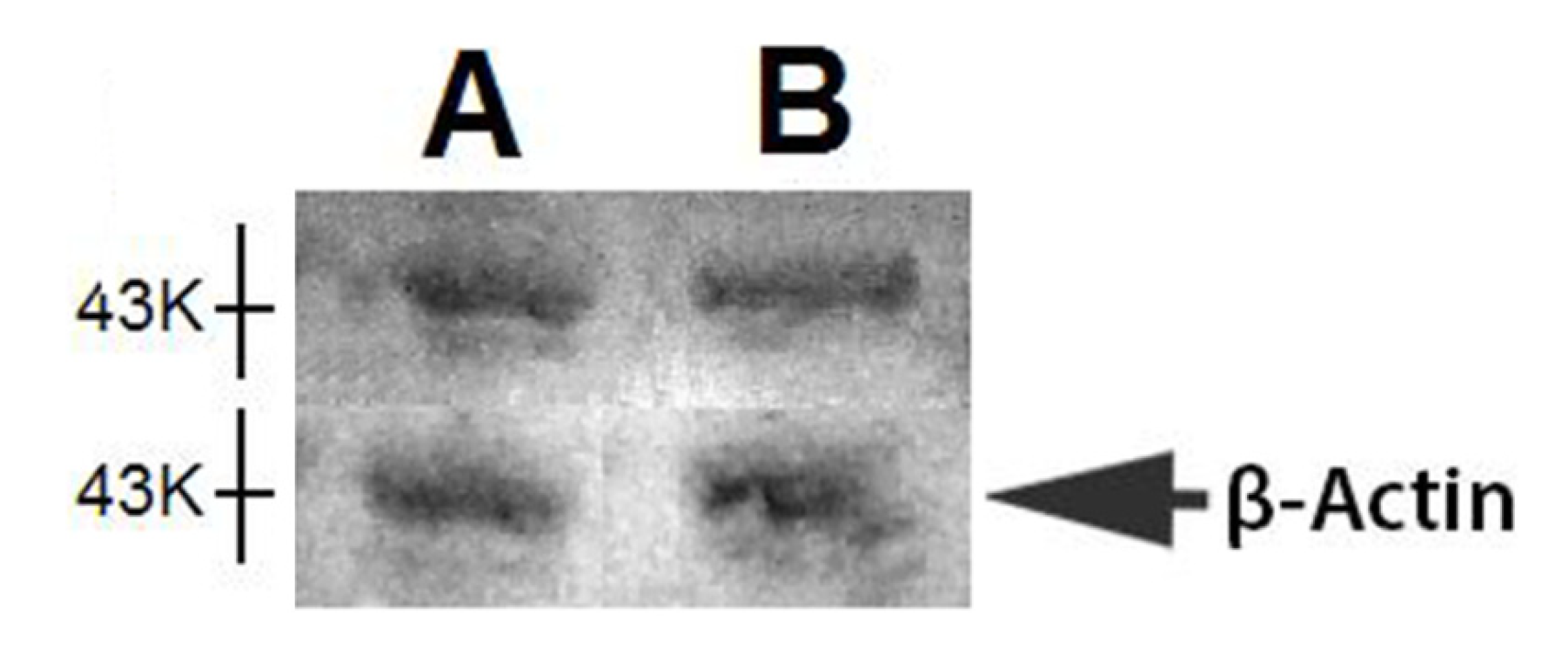
2.2. Western Blot of Nkx6.1

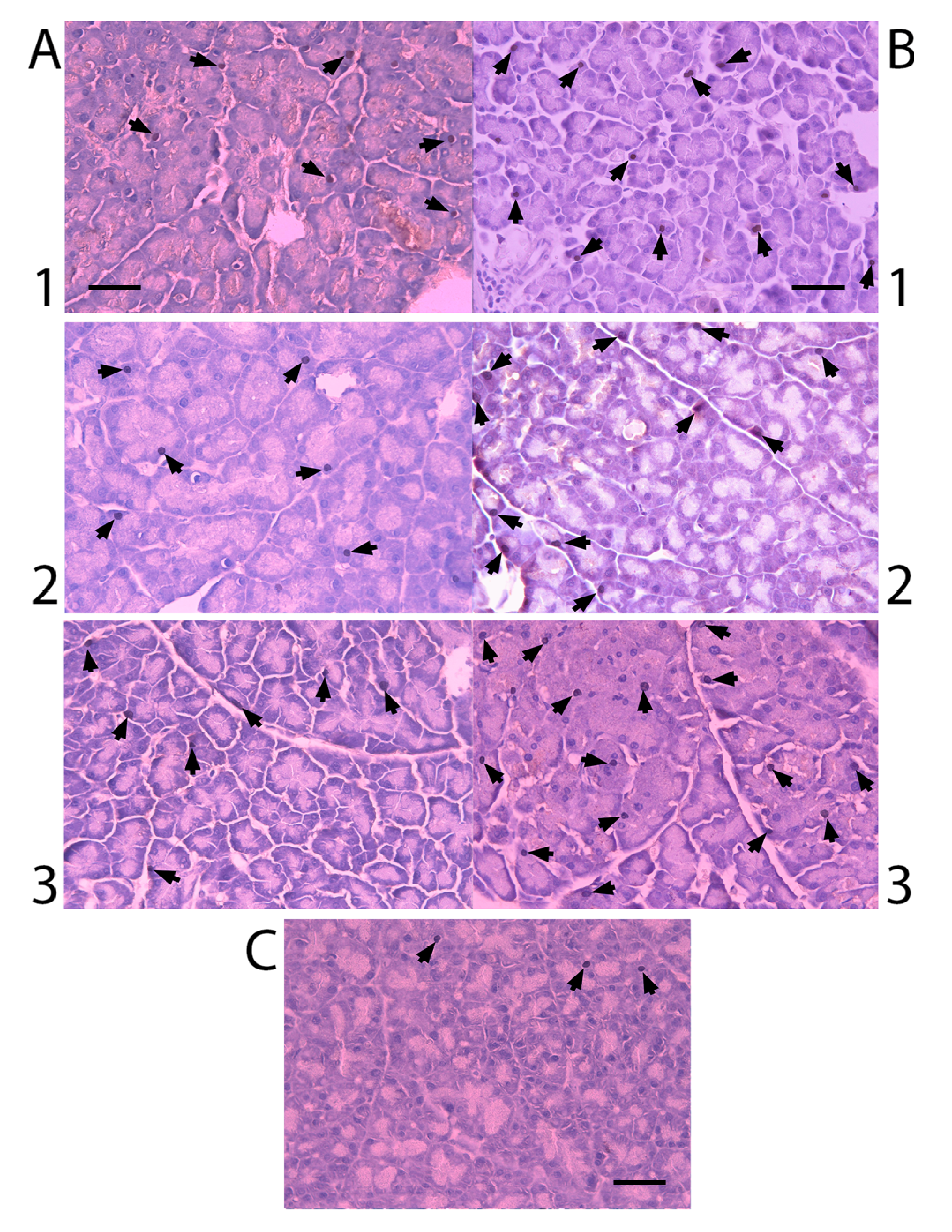
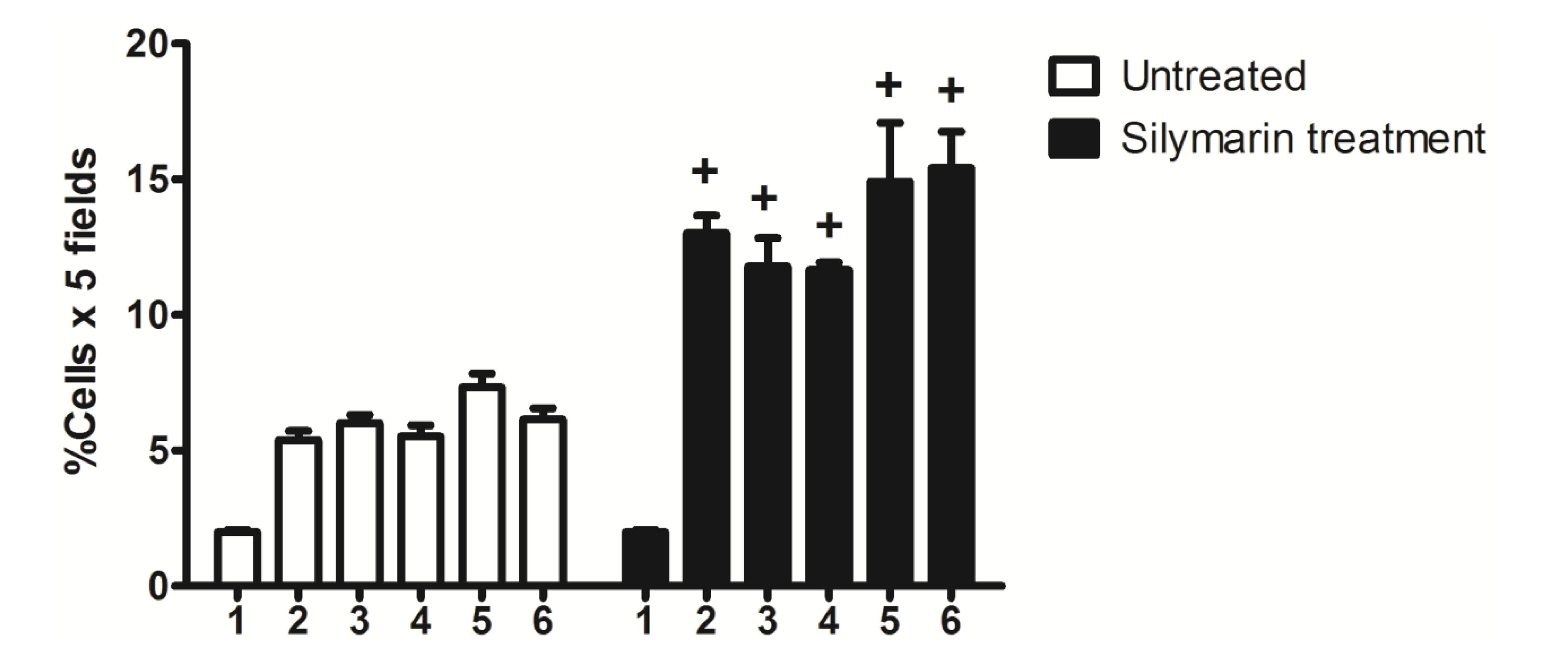
2.3. Nkx6.1 Immunolabelling




2.5. Serum Insulin Levels

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days after pancreatectomy | Serum insulin (ng/mL) Untreated group | Serum insulin (ng/mL) Silymarin treated group |
|---|---|---|
| Control | 1.28 ± 0.02 | 1.28 ± 0.02 |
| 3 | 0.350 ± 0.089 | 0.777 ± 0.122 |
| 7 | 1.158 ± 0.474 | 0.487 ± 0.023 |
| 14 | 0.293 ± 0.087 | 0.505 ± 0.018 |
| 21 | 0.175 ± 0.351 | 0.782 ± 0.202 + |
| 42 | 1.224 ± 0.292 | 2.79 ± 0.342 *,+ |
| 63 | 1.123 ± 0.357 | 3.206 ± 0.272 *,+ |
2.6. Serum Glucose Levels
| Days after pancreatectomy | Serum glucose (mM) Untreated group | Serum glucose (mM) Silymarin treated group |
|---|---|---|
| Control | 5.9 ± 0.004 | 5.9 ± 0.004 |
| 3 | 9.00 ± 1.575 | 6.2 ± 0.231 |
| 7 | 7.93 ± 0.493 | 9.00 ± 0.151 |
| 14 | 9.74 ± 0.788 | 7.2 ± 0.352 + |
| 21 | 7.54 ± 0.222 | 8.02 ± 1.2 |
| 42 | 10.55 ± 0.858 | 7.25 ± 0.256 + |
| 63 | 9.34 ± 0.457 | 6.35 ± 0.252 + |
2.7. Discussion
3. Experimental
3.1. Animal Treatments
3.2. Blood and Tissue Collection
3.3. RNA Isolation and RT–PCR Assay for Nkx6.1 and Insulin
3.4. Serum Insulin Levels
3.5. β-Cell Neogenesis Assessment
3.6. Pancreatic Tissue Nkx6.1 Immunoassay
3.7. Western Blot Analysis
3.8. Statistical Methods
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Weir, G.C.; Bonner-Weir, S. Five stages of evolving beta cell dysfunction during progression to diabetes. Diabetes 2004, 53, S16–S21. [Google Scholar] [CrossRef]
- Bernardo, A.S.; Hay, C.W.; Docherty, K. Pancreatic transcription factors and their role in the birth, life and survival of the pancreatic β cell. Mol. Cell Endocrinol. 2008, 294, 1–9. [Google Scholar] [CrossRef]
- Iype, T.; Taylor, D.G.; Ziesmann, S.M.; Garmey, J.C.; Watada, H.; Mirmira, R.G. The transcriptional repressor Nkx6.1 also functions as a deoxyribonucleic acid context-dependent transcriptional activator during pancreatic β-cell differentiation: Evidence for feedback activation of the nkx6.1 gene by Nkx6.1. Mol. Endocrinol. 2004, 18, 1363–1375. [Google Scholar] [CrossRef]
- Sander, M.; Sussel, L.; Conners, J.; Scheel, D.; Kalamaras, J.; Cruz, D.F.; Schwitzgebel, V.; Hayes-Jordan, A.; German, M. Homeobox gene Nkx6.1 lies downstream of Nkx2.2 in the major pathway of β-cell formation in the pancreas. Development 2000, 127, 5533–5540. [Google Scholar]
- Slack, J.M. Developmental biology of the pancreas. Development 1995, 121, 1569–1580. [Google Scholar]
- Oster, A.; Jensen, J.; Serup, P.; Galante, P.; Madsen, O.D.; Larson, L.I. Rat endocrine pancreatic development in relation to two homeobox gene products (Pdx-1 and Nkx 6.1). J. Histochem. Cytochem. 1998, 46, 707–715. [Google Scholar] [CrossRef]
- Sander, M.; German, M.S. The beta cell transcription factors and development of the pancreas. J. Mol. Med. 1997, 75, 327–340. [Google Scholar] [CrossRef]
- Schwitzgebel, V.M.; Scheel, D.W.; Conners, J.R.; Kalamaras, J.; Lee, J.E.; Anderson, D.J.; Sussel, L.; Johnson, J.D.; German, M.S. Expression of neurogenin3 reveal an islet cell precursor population in the pancreas. Development 2000, 127, 3533–3542. [Google Scholar]
- Ahlgren, U.; Jonson, J.; Jonson, L.; Simu, K.; Edlund, H. Beta cell specific inactivation of the mouse Ipf1/Pdx1 gene results in loss of the beta-cell phenotype and maturity onset diabetes. Genes Dev. 1998, 12, 1763–1768. [Google Scholar] [CrossRef]
- Schaffer, A.E.; Taylor, B.L.; Benthuysen, J.R.; Liu, J.; Thorel, F.; Yuan, W.; Jiao, Y.; Kaetsner, K.H.; Herrera, P.L.; Magnuson, M.A.; et al. Nkx6.1 controls a gene regulatory network required for establishing and maintaining pancreatic beta cell. PloS Genet. 2013, 9, 1–14. [Google Scholar]
- Soto, C.; Mena, R.; Luna, J.; Cerbón, M.; Larrieta, E.; Vital, P.; Uría, E.; Sánchez, M.; Recoba, R.; Barrón, H.; et al. Silymarin induces recovery of pancreatic function after alloxan damage in rats. Life Sci. 2004, 75, 2167–2180. [Google Scholar] [CrossRef]
- Althagafy, H.S.; Meza-Aviña, M.E.; Oberlies, N.H.; Croatt, M.P. Mechanistic Study of the Biomimetic Synthesis of Flavonolignan Diastereoisomers in Milk Thistle. J. Org. Chem. 2013, 78, 7594–7600. [Google Scholar] [CrossRef]
- Abenavoli, L.; Capasso, R.; Milic, N.; Capasso, F. Milk thistle in liver diseases: Past, present, future. Phytother. Res. 2010, 24, 1423–1432. [Google Scholar] [CrossRef]
- Schaffer, A.E.; Freude, K.K.; Nelson, S.B.; Sander, M. Nkx6.1 transcription factor and Ptf1a function as antagonistic lineage determinants in multipotent pancreatic progenitors. Dev. Cell 2010, 18, 1022–1029. [Google Scholar] [CrossRef]
- Nelson, S.B.; Schaffer, A.E.; Sander, M. The transcription factors Nkx6.1 and Nkx6.2 possess equivalent activities promoting beta-cell fate specification in Pdx1+ pancreatic progenitor cells. Development 2007, 134, 2491–2500. [Google Scholar] [CrossRef]
- Wang, H.; Maechler, P.; Ritz-Laser, B.; Hagenfeldt, K.A.; Ishihara, H.; Philippe, J.; Wollheim, C.B. Pdx1 level defines pancreatic gene expression pattern and cell lineage differentiation. J. Biol. Chem. 2001, 276, 25279–25286. [Google Scholar]
- Schisler, J.C.; Jensen, P.B.; Taylor, D.G.; Becker, T.C.; Knop, F.K.; Takekawa, S.; German, M.; Weir, G.C.; Lu, D.; Mirmira, R.G.; et al. The Nkx6.1 homeodomain transcription factor directly suppress glucagon expression and regulates glucose stimulated insulin secretion in islet b-cells. Proc. Natl. Acad. Sci. USA 2005, 102, 7297–7302. [Google Scholar]
- Mirmira, R.G.; Watada, H.; German, M.S. Beta-cell differentiation factor Nkx6.1 contains distinct DNA binding interference and transcriptional repression domains. J. Biol. Chem. 2000, 275, 14743–14751. [Google Scholar] [CrossRef]
- Taylor, D.G.; Babu, D.; Mirmira, R.G. The C-terminal domain of the β cell homeodomain factor Nkx6.1 enhances sequence selective DNA binding at the insulin promoter. Biochemistry 2005, 44, 11269–11278. [Google Scholar] [CrossRef]
- Peshvaria, M.; Larmie, B.L.; Lausier, J.; Satish, B.; Habibovic, A.; Roskens, V.; LaRock, K.; Everill, B.; Leahy, J.L.; Jetton, T.L. Regulation of pancreatic β-cell regeneration in the normoglycemic 60% partial pancreatectomy mouse. Diabetes 2006, 55, 3289–3298. [Google Scholar] [CrossRef]
- Li, W.C.; Rukstalis, J.M.; Nishimura, W.; Tchipashvil, V.; Habener, J.F.; Sharma, A.; Bonner-Weir, S. Activation of pancreatic-duct-derived progenitor cells during pancreas regeneration in adult rats. J. Cell Sci. 2010, 123, 2792–2802. [Google Scholar] [CrossRef]
- Soto, C.; Raya, L.; Juárez, J.; Pérez, J.; González, I. Effect of Silymarin in Pdx-1 expression and the proliferation of pancreatic β-cells in a pancreatectomy model. Phytomedicine 2014, 21, 233–239. [Google Scholar] [CrossRef]
- Röhrdanz, E.; Ohler, S.; Tran-Thi, Q.H.; Kahl, R. The phytoestrogen Daidzein affects the antioxidant enzyme system of rat hepatoma H4IIE cells. J. Nutr. 2002, 132, 370–375. [Google Scholar]
- Peck, M.C.; Fisher, R.F.; Long, S.R. Diverse flavonoids stimulate NodD1 binding to nod gene promoters in Sinorhizobium meliloti. J. Bacteriol. 2006, 188, 5417–5427. [Google Scholar] [CrossRef]
- Schisler, J.C.; Fueger, P.T.; Babu, D.A.; Hohmeier, H.E.; Tessem, J.S.; Lu, D.; Becker, C.T.; Naziruddin, B.; Levy, M.; Mirmira, R.G.; et al. Stimulation of human and rat islet β-cell proliferation with retention of function by the homeodomain transcription factor Nkx6.1. Mol. Cell Biol. 2008, 28, 3465–3476. [Google Scholar] [CrossRef]
- Gerrish, K.; Cisell, M.; Stein, R. The role of hepatic nuclear factor 1a and PDX-1 transcriptional regulation of the pdx-1 gene. J. Biol. Chem. 2001, 275, 47775–47784. [Google Scholar]
- Smith, S.; Watada, H.; Scheel, D.; Mrejen, C.; German, M. Autorregulation and maturity onset diabetes of the young transcription factors control the human PAX4 promoter. J. Biol. Chem. 2000, 275, 36910–36919. [Google Scholar] [CrossRef]
- Aota, S.; Nakajima, N.; Sakamoto, R.; Watanabe, S.; Ibaraki, N.; Okazaki, K. Pax6 autorregulation mediated by direct interaction of Pax6 protein with the head Surface ectoderm-specific enhancer of the mouse Pax6 gene. Dev. Biol. 2003, 257, 1–13. [Google Scholar] [CrossRef]
- Smith, S.; Watada, H.; German, M. Neurogenin 3 activates the islet differentiation program while its own expression. Mol. Endocrinol. 2004, 18, 142–149. [Google Scholar] [CrossRef]
- Jorgensen, M.; Vestergard, P.; Petersen, H.; Ericson, J.; Madsen, O.; Serup, P. Cloning and DNA-binding properties of the rat pancreatic β-cell-specific factor Nkx6.1. FEBS Lett. 1999, 461, 287–294. [Google Scholar] [CrossRef]
- Stringer, K.F.; Ingles, C.J.; Greenblatt, J. Direct and selective binding of an acidic transcriptional to the TATA-box factor TFIID. Nature 1990, 345, 783–786. [Google Scholar] [CrossRef]
- Roberts, S.G.; Ha, I.; Maldonado, E.; Reinberg, D.; Green, M.R. Interaction between anacidic activator and transcription factor TFIIB is required for transcriptionactivation. Nature 1993, 363, 741–744. [Google Scholar] [CrossRef]
- Guo, S.; Dai, C.; Guo, M.; Taylor, B.; Harmon, J.S.; Sander, M.; Robertson, P.R.; Powers, A.C.; Stein, R. Inactivation of specific β cell transcription factors in type 2 diabetes. J. Clin. Invest. 2013, 123, 3305–3316. [Google Scholar] [CrossRef]
- Soto, C.P.; Pérez, B.L.; Favari, L.P.; Reyes, J.L. Prevention of alloxan-induced diabetes mellitus in the rat by silymarin. Comp. Biochem. Physiol. Part C 1998, 119, 125–129. [Google Scholar] [CrossRef]
- Soto, C.P.; Recoba, R.; Barrón, H.; Alvarez, C.; Favari, L. Silymarin increases antioxidant enzymes in alloxan-induced diabetes mellitus in rat pancreas. Comp. Biochem. Physiol. Part C 2003, 136, 205–212. [Google Scholar] [CrossRef]
- Muriel, P.; Garciapiña, T.; Perez-Alvarez, V.; Mourelle, M. Silymarin protects against paracetamol-induced lipid peroxidation and liver damage. J. Appl. Toxicol. 1992, 12, 439–442. [Google Scholar] [CrossRef]
- Tsai, J.H.; Liu, J.Y.; Wu, T.T.; Ho, P.C.; Huang, C.Y.; Shyu, J.C.; Hsieh, Y.S.; Tsai, C.C.; Liu, Y.C. Effects of silymarin on the resolution of liver fibrosis induced by carbon tetrachloride in rats. J. Viral Hepat. 2008, 15, 508–514. [Google Scholar] [CrossRef]
- Olansky, L.; Welling, C.; Giddings, S.; Adler, S.; Bourey, R.; Dowse, G.; Serjeantson, S.; Zimmet, P.; Permut, M. A variant insulin promoter in non-insulin-dependent diabetes mellitus. J. Clin. Invest. 1992, 89, 1596–1602. [Google Scholar] [CrossRef]
- Schmittgen, T.; Zakrajsek, B. Effect of experimental treatment on housekeeping gene expression: Validation by real-time, quantitative RT-PCR. J. Biochem. Biophys. Methods 2000, 46, 69–81. [Google Scholar] [CrossRef]
- Montanya, E.; Téllez, N. Pancreatic remodeling: Beta-cell apoptosis, proliferation and neogenesis, and the measurement of beta-cell mass and of individual beta-cell size. In Type 2 Diabetes, Methods in Molecular Biology; Stocker, C., Ed.; Humana Press: New York, NY, USA, 2009; Series volume 560, Charpter 11; pp. 137–158. [Google Scholar]
- Weir, G.; Bonner-Weir, S.; Leahy, J. Islet mass and function in diabetes and transplantation. Diabetes 1990, 39, 401–405. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Soto, C.; Raya, L.; Pérez, J.; González, I.; Pérez, S. Silymarin Induces Expression of Pancreatic Nkx6.1 Transcription Factor and β-Cells Neogenesis in a Pancreatectomy Model. Molecules 2014, 19, 4654-4668. https://doi.org/10.3390/molecules19044654
Soto C, Raya L, Pérez J, González I, Pérez S. Silymarin Induces Expression of Pancreatic Nkx6.1 Transcription Factor and β-Cells Neogenesis in a Pancreatectomy Model. Molecules. 2014; 19(4):4654-4668. https://doi.org/10.3390/molecules19044654
Chicago/Turabian StyleSoto, Claudia, Luis Raya, Julia Pérez, Imelda González, and Salud Pérez. 2014. "Silymarin Induces Expression of Pancreatic Nkx6.1 Transcription Factor and β-Cells Neogenesis in a Pancreatectomy Model" Molecules 19, no. 4: 4654-4668. https://doi.org/10.3390/molecules19044654
APA StyleSoto, C., Raya, L., Pérez, J., González, I., & Pérez, S. (2014). Silymarin Induces Expression of Pancreatic Nkx6.1 Transcription Factor and β-Cells Neogenesis in a Pancreatectomy Model. Molecules, 19(4), 4654-4668. https://doi.org/10.3390/molecules19044654