Potential Anti-Inflammatory Effects of the Hydrophilic Fraction of Pomegranate (Punica granatum L.) Seed Oil on Breast Cancer Cell Lines



Abstract
:1. Introduction
2. Results and Discussion
2.1. Extraction of Hydrophilic Compounds from Pomegranate Seed Oil

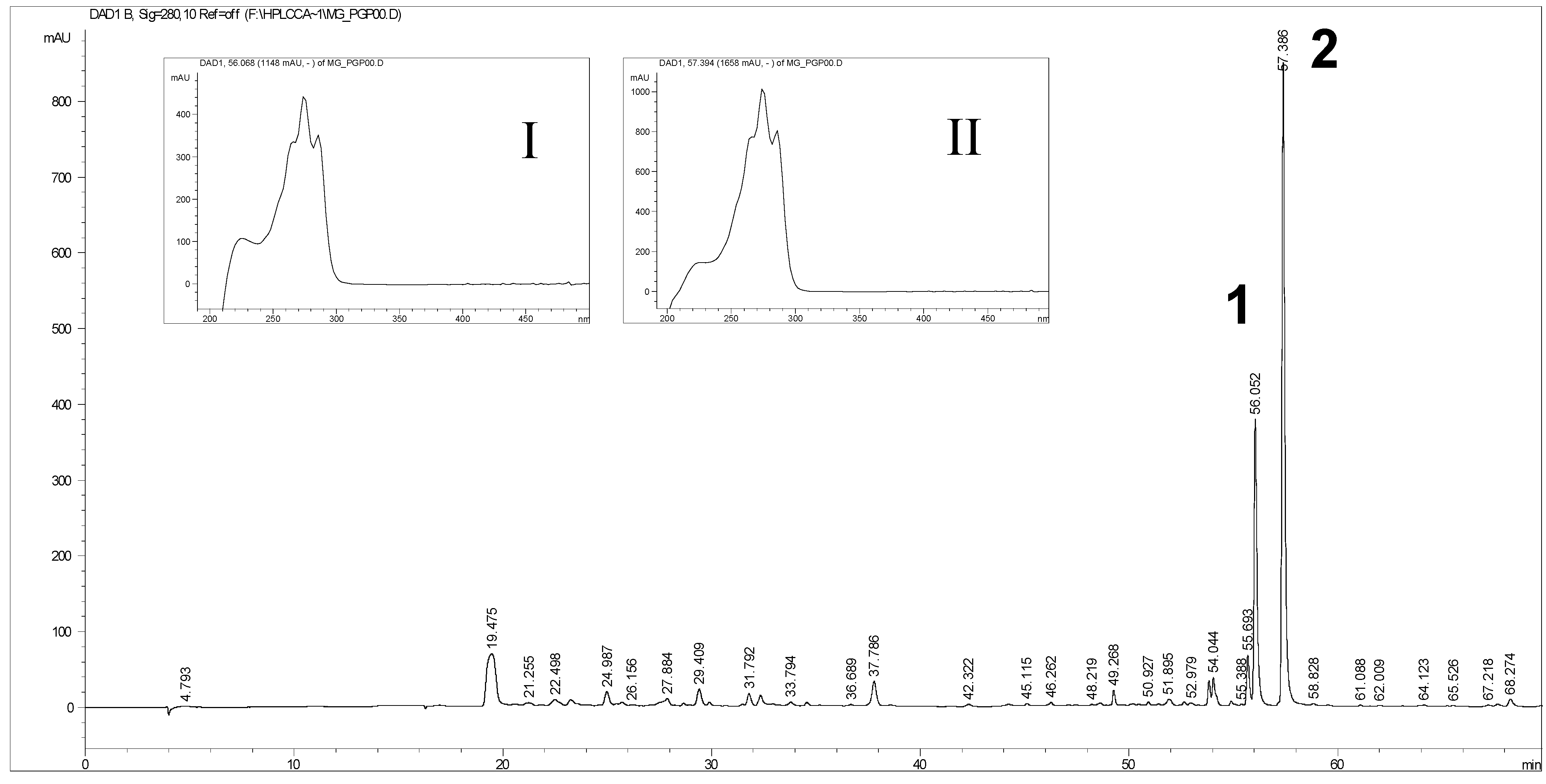{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polar Extracts From PSO | Yield (%) | TPCmg GAE/100 g oil | DPPH (I%) |
| 8.93 ± 1.02 | 23.07 ± 1.44 | 96.80 ± 8.93 |


2.2. Colorimetric Assay with Sulforhodamine B

2.3. Apoptosis Assay
- Live cells which are not undergoing to apoptosis: annexin V (−) and cells dead marker (−);
- Cells in early apoptosis: annexin V (+) and cells dead marker (−);
- Cells in adavanced apoptosis: annexin V (+) and cells dead marker (+);
- Dead cells which do not cross the apoptotic process (necrosis): annexin V (−) and cells dead marker (+).
| Cells | Live Cells (%) | Cells in Early Apoptosis (%) | Cells in Late Apoptosis (%) | Dead Cells (%) |
|---|---|---|---|---|
| MCF-7 non treated | 95.30 ± 1.02 | 0.95 ± 0.02 | 3.35% ± 1.21% | 0.40 ± 0.04 |
| MCF-7 treated | 92.85 ± 1.15 | 3.50 ± 0.77 | 2.65% ± 0.07% | 1.00 ± 0.09 |
| MDA-MB-231 non treated | 96.79 ± 1.09 | 1.31 ± 0.42 | 1.86% ± 0.54% | 0.05 ± 0.02 |
| MDA-MB-231 treated | 95.75 ± 2.04 | 2.95 ± 0.61 | 1.10% ± 0.05% | 0.20 ± 0.09 |
| Cells | G0/G1 (%) | S (%) | G2/M (%) |
|---|---|---|---|
| MCF-7 Non treated | 5.0 ± 1.2 | 94.2 ± 6.4 | 0.8 ± 0.1 |
| MCF-7 treated | 79.2 ± 5.3 | 7.9 ± 1.6 | 12.9 ± 2.1 |
| MDA-MB-231 non treated | 20.8 ± 3.6 | 18.9 ± 1.2 | 60.3 ± 4.6 |
| MDA-MB-231 treated | 67.5 ± 6.1 | 19.7 ± 2.1 | 12.8 ± 2.3 |
2.4. Cell Cycle Assay
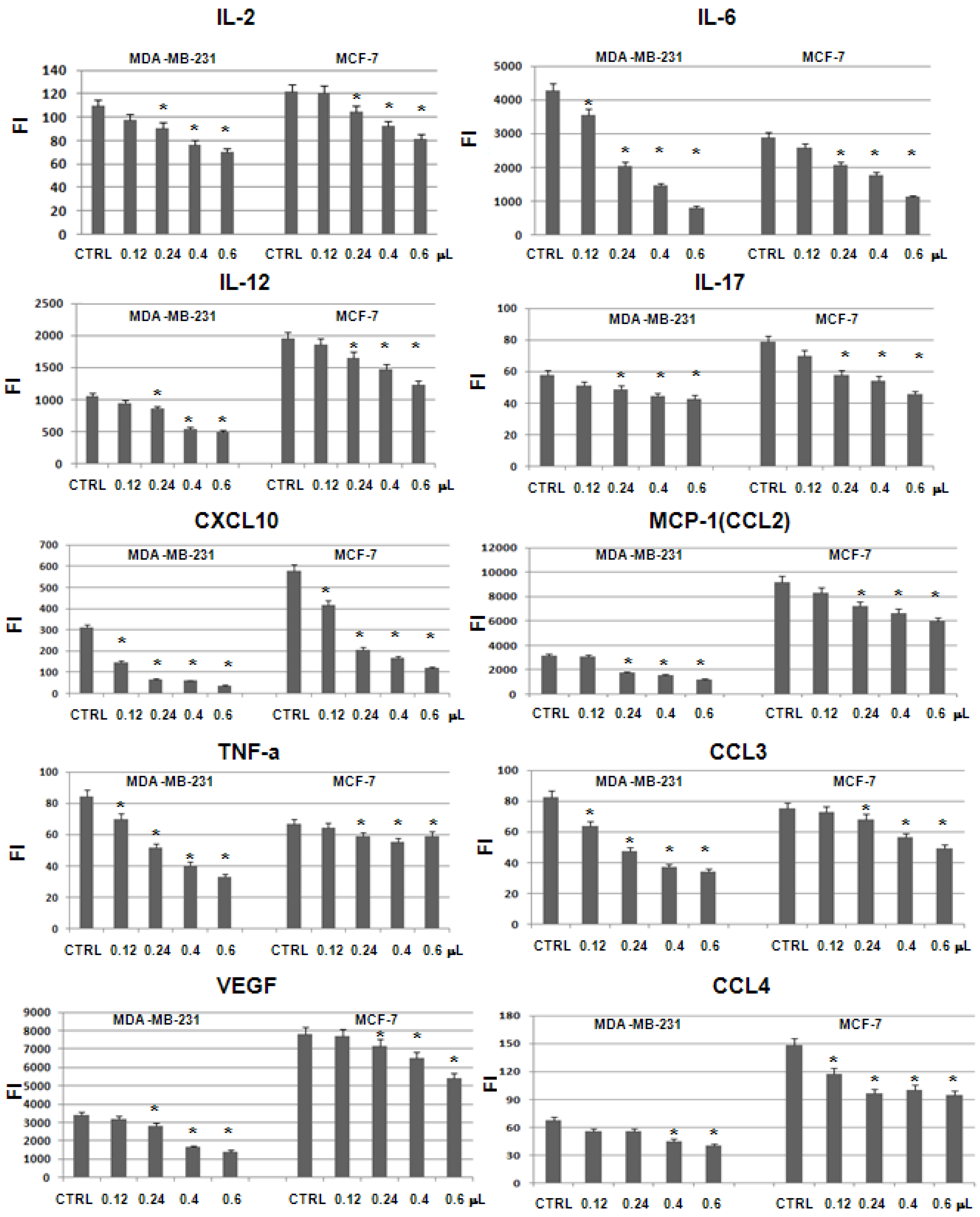
2.5. Evaluation of Cytokine Levels on MDA-MB-231 and MCF-7 Supernatants

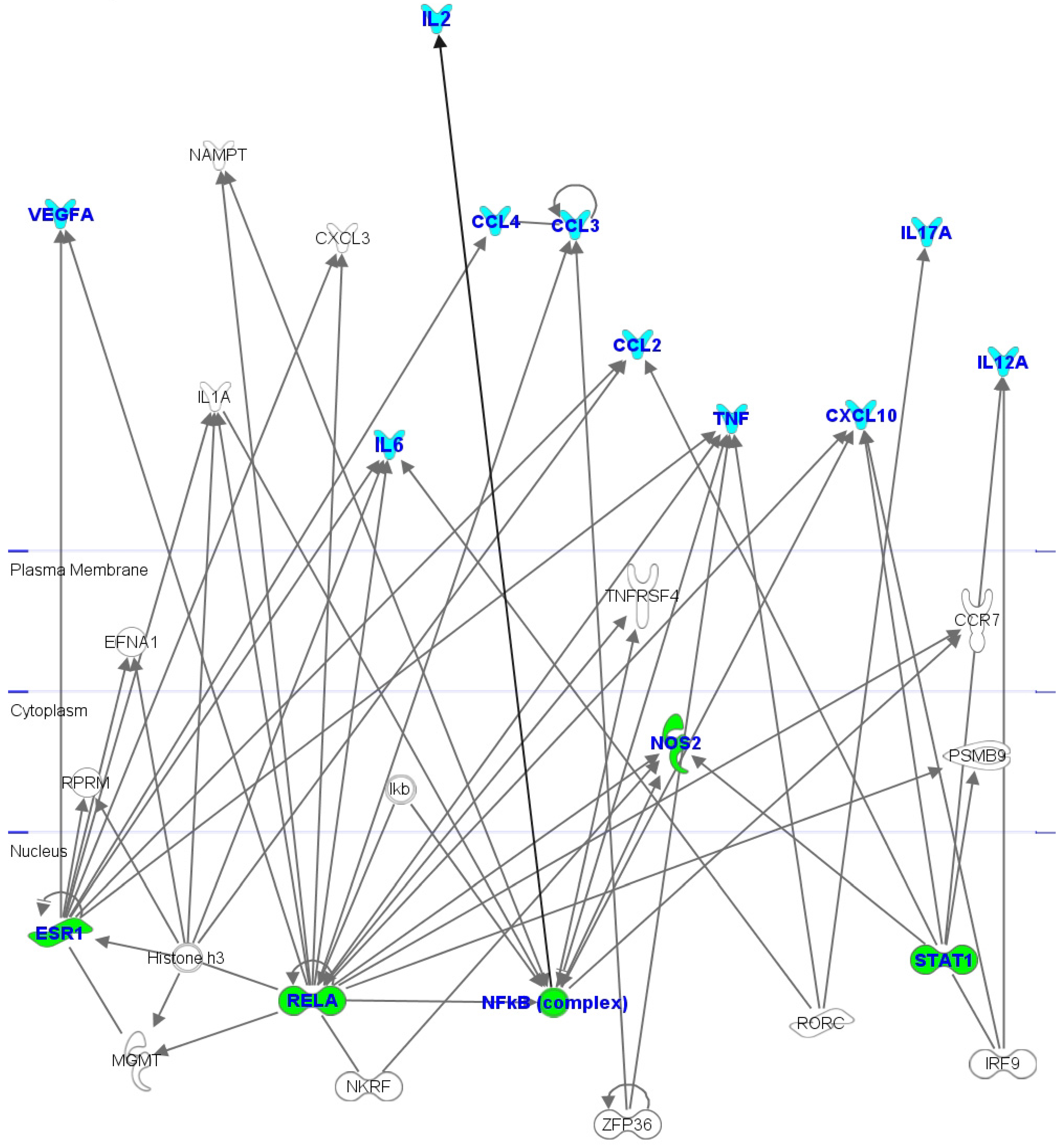
2.6. Interactomic Studies

3. Experimental
3.1. Samples and Chemicals
3.2. Oil Extraction
3.3. Extraction and Determination of Total Phenolics
3.4. RP-HPLC-DAD Analysis
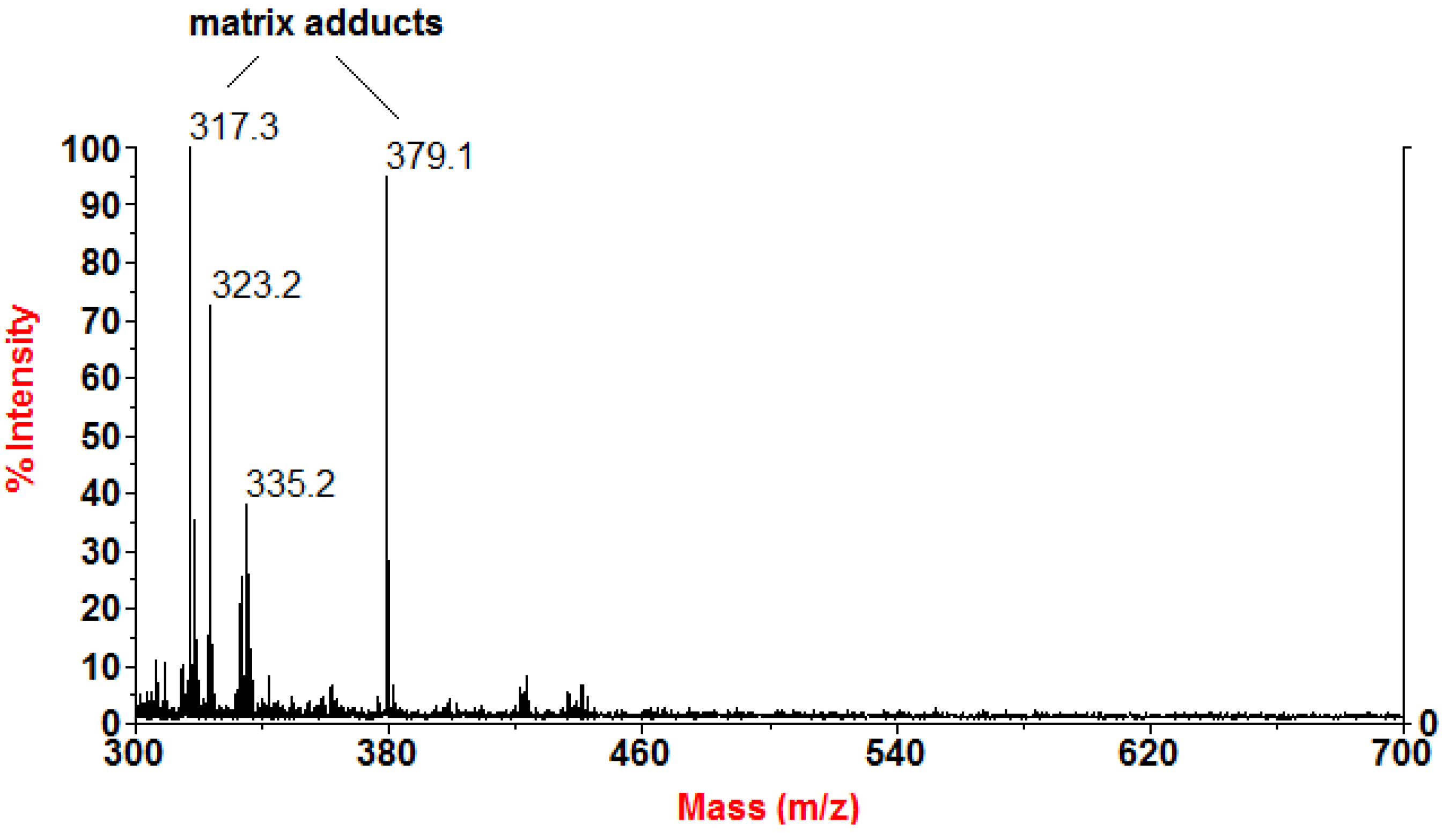
3.5. MALDI-TOF MS Analysis
3.6. Antioxidant Activity (DPPH Assay)
3.7. Cell Culture
3.8. Colorimetric Assay with Sulforhodamine B
3.9. Apoptosis Assay
3.10. Cell Cycle Assay
3.11. Bio-Plex Assay
3.12. Bioinformatics Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, Z.; Pan, Z.; Ma, H.; Atungulu, G.G. Extract of phenolics from pomegranate peels. Open Food Sci. J. 2011, 5, 17–25. [Google Scholar] [CrossRef]
- Lansky, E.P.; Newman, R.A. Punica granatum (pomegranate) and its potential for prevention and treatment of inflammation and cancer. J. Ethnopharmacol. 2007, 109, 177–206. [Google Scholar] [CrossRef]
- Rasheed, Z.; Akhtar, N.; Anbazhagan, N.A.; Ramamurthy, S.; Shukla, M.; Haqqi, T.M. Polyphenol-rich pomegranate fruit extract (POMx) suppresses PMACI induced expression of pro-inflammatory cytokines by inhibiting the activation of MAP Kinases and NF-κB in human KU812 cells. J. Inflam. 2009, 6, 1. [Google Scholar] [CrossRef]
- Larrosa, M.; González-Sarrías, A.; García-Conesa, M.T.; Francisco, A.T.B.; Espín, J.C. Urolithins ellagic acid-derived metabolites produced by human colonic microflora exhibit estrogenic and antiestrogenic activities. J. Agric. Food Chem. 2006, 54, 1611–1620. [Google Scholar] [CrossRef]
- Elfalleh, W.; Tlili, N.; Nasri, N.; Yahia, Y.; Hannachi, H.; Chaira, N.; Ying, M.; Ferchichi, A. Antioxidant capacities of phenolic compounds and tocopherols from tunisian pomegranate (Punica granatum) fruits. J. Food Sci. 2011, 76, 707–713. [Google Scholar] [CrossRef]
- Fadavi, A.; Barzegar, M.; Azizi, M.H. Determination of fatty acids and total lipid content in oilseed of 25 pomegranates varieties grown in Iran. J. Food Compos. Anal. 2006, 19, 676–680. [Google Scholar] [CrossRef]
- Kasimsetty, S.G.; Bialonska, D.; Reddy, M.K.; Ma, G.; Khan, S.I.; Ferreira, D. Colon cancer chemopreventive activities of pomegranate ellagitannins and urolithins. J. Agric. Food Chem. 2010, 58, 2180–2187. [Google Scholar] [CrossRef]
- Albrecht, M.; Jiang, W.; Kumi-Diaka, J.; Lansky, E.P.; Gommersall, L.M. Pomegranate extracts potently suppress proliferation Xenograft growth and invasion of human prostate cancer cells. J. Med. Food 2004, 7, 274–283. [Google Scholar]
- Adhami, V.M.; Khan, N.; Mukhtar, H. Cancer chemoprevention by pomegranate: Laboratory and clinical evidence. Nutr. Cancer 2009, 61, 811–815. [Google Scholar] [CrossRef]
- Afaq, F.; Zaid, M.A.; Khan, N.; Dreher, M.; Mukhtar, H. Protective effect of pomegranate-derived products on UVB-mediated damage in human reconstituted skin. Exp. Dermatol. 2009, 18, 553–561. [Google Scholar] [CrossRef]
- Schubert, S.Y.; Lansky, E.P.; Neeman, I. Antioxidant and eicosanoid enzyme inhibition properties of pomegranate seed oil and fermented juice flavonoids. J. Ethnopharmacol. 1999, 66, 11–17. [Google Scholar] [CrossRef]
- Straccia, M.C.; Siano, F.; Coppola, R; la Cara, F.; Volpe, M.G. Extraction and characterization of vegetable oils from cherry seed by different extraction processes. Chem. Eng. Trans. 2012, 27, 391–396. [Google Scholar]
- Kohno, H.; Suzuki, R.; Yasui, Y.; Hosokawa, M.; Miyashita, K.; Tanaka, T. Pomegranate seed oil rich in conjugated linolenic acid suppresses chemically induced colon carcinogenesis in rats. Cancer Sci. 2004, 95, 481–486. [Google Scholar] [CrossRef]
- Arzu, A.B.; Tulay, O.; Lutfiye, Y.E. The Therapeutic Potential of Pomegranate and Its Products for Prevention of Cancer Cancer Prevention-From Mechanisms to Translational Benefits; Tech–Open Access Company: Rijeka, Croatia, 2012; pp. 331–372. [Google Scholar]
- Hora, J.J.; Maydew, E.R.; Lansky, E.P.; Dwivedi, C. Chemopreventive effects of pomegranate seed oil on skin tumor development in CD1 mice. J. Med. Food 2003, 6, 157–161. [Google Scholar] [CrossRef]
- Faria, A.; Calhau, C. The Bioactivity of pomegranate: Impact on health and disease. CRC Crit. Rev. Food Sci. Nutr. 2011, 51, 626–634. [Google Scholar] [CrossRef]
- Caligiani, A; Bonzanini, F.; Palla, G.; Cirlini, M.; Bruni, R. Characterization of a potential nutraceutical ingredient: Pomegranate (Punica granatum L.) seed oil unsaponifiable fraction. Plant. Foods Hum. Nutr. 2010, 65, 277–283. [Google Scholar] [CrossRef]
- Adams, L.S.; Seeram, N.P.; Aggarwal, B.B.; Takada, Y.; Sand, D.; Heber, D. Pomegranate juice total pomegranate ellagitannins and punicalagin suppress inflammatory cell signaling in colon cancer Cells. J. Agric. Food Chem. 2006, 54, 980–985. [Google Scholar]
- Malik, A.; Afaq, F.; Sarfaraz, S.; Adhami, V.M.; Syed, D.N.; Mukhtar, H. Pomegranate fruit Juice for chemoprevention and chemotherapy of prostate cancer. Proc. Natl. Acad. Sci. USA 2005, 2, 14813–14818. [Google Scholar]
- Pande, G.; Akoh, C.C. Antioxidant capacity and lipid characterization of six Georgia-grown pomegranate cultivars. J. Agric. Food Chem. 2009, 57, 9427–9436. [Google Scholar] [CrossRef]
- Jing, P.; Tian, Y.; Shi, H.; Sheng, Y.; Slavin, M.; Gao, B.; Liud, L.; Yu, L. Antioxidant properties and phytochemical composition of China-grown pomegranate seeds. Food Chem. 2012, 132, 1457–1464. [Google Scholar] [CrossRef]
- Turtygin, A.V; Deineka, V.I.; Deineka, L.A. Determination of the triglyceride composition of pomegranate seed oil by reversed-phase HPLC and spectrophotometry. J. Anal. Chem. 2013, 68, 558–563. [Google Scholar]
- Hontecillas, R.; O’Shea, M.; Einerhand, A.; Diguardo, M.; Bassaganya-Riera, J. Activation of PPAR gamma and alpha by punicic acid ameliorates glucose tolerance and suppresses obesity-related inflammation. J. Am. Coll. Nutr. 2009, 28, 184–195. [Google Scholar] [CrossRef]
- Bassaganya-Riera, J.; DiGuardo, M.; Climent, M.; Vives, C.; Carbo, A.; Jouni, Z.E.; Einerhand, A.W.; O’Shea, M.; Hontecillas, R. Activation of PPARγ and δ by dietary punicic acid ameliorates intestinal inflammation in mice. Br. J. Nutr. 2011, 106, 878–886. [Google Scholar] [CrossRef]
- Dehqanzada, Z.A.; Storrer, C.E.; Hueman, M.T.; Foley, R.J.; Harris, K.A.; Jama, Y.H.; Shriver, C.D.; Ponniah, S.; Peoples, G.E. Assessing serum cytokine profiles in breast cancer patients receiving a HER2/neu vaccine using Luminex technology. Oncol. Rep. 2007, 17, 687–694. [Google Scholar]
- Salcedo, R.; Ponce, M.L.; Young, H.A.; Wasserman, K.; Ward, J.M.; Kleinman, H.K.; Oppenheim, J.J.; Murphy, W.J. Human endothelial cells express CCR2 and respond to MCP-1: Direct role of MCP-1 in angiogenesis and tumor progression. Blood 2000, 196, 34–40. [Google Scholar]
- Kolls, J.K.; Linden, A. Interleukin-17 family members and infiammation. Immunity 2004, 21, 467–476. [Google Scholar] [CrossRef]
- Fabregat, I. Dysregulation of apoptosis in hepatocellular carcinoma cells. World J. Gastroenterol. 2009, 15, 513–520. [Google Scholar] [CrossRef]
- Feurino, L.W.; Zhang, Y.; Bharadwaj, U.; Zhang, R.; Li, F.; Fisher, W.E.; Brunicardi, F.C.; Chen, C.; Yao, Q.; Min, L. IL-6 stimulates Th2 type cytokine secretion and upregulates VEGF and NRP-1 expression in pancreatic cancer cells. Cancer Biol. Ther. 2007, 6, 1096–1100. [Google Scholar] [CrossRef]
- Ren, M.; Guo, Q.; Guo, L.; Lenz, M.; Qian, F.; Koenen, R.R.; Xu, H.; Schilling, A.B.; Weber, C.; Ye, R.D.; et al. WJPolymerization of MIP-1 chemokine (CCL3 and CCL4) and clearance of MIP-1 by insulin-degrading enzyme. EMBO J. 2010, 129, 3952–3966. [Google Scholar]
- Waldmann, T.A. The biology of interleukin-2 and interleukin-15: Implications for cancer therapy and vaccine design. Nat. Rev. Immunol. 2006, 6, 595–601. [Google Scholar] [CrossRef]
- Dun, X.; Li, X.Q.; Li, L.; Xu, Y.Y.; Feng, Y.M. The detection of ESR1/PGR/ERBB2 mRNA levels by RT-QPCR: A better approach for subtyping breast cancer and predicting prognosis. Breast Cancer Res. Treat. 2013, 138, 59–67. [Google Scholar] [CrossRef]
- Glynn, S.A.; Boersma, B.J.; Dorsey, T.H.; Yi, M.; Yfantis, H.G.; Ridnour, L.A.; Martin, D.N.; Switzer, C.H.; Hudson, R.S.; Wink, D.A.; et al. Increased NOS2 predicts poor survival in estrogen receptor-negative breast cancer patients. J. Clin. Invest. 2010, 120, 3843–3854. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticult. 1965, 16, 144–158. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Rusolo, F.; Pucci, B.; Colonna, G.; Capone, F.; Guerriero, E.; Milone, M.R.; Nazzaro, M.; Volpe, M.G.; di Bernardo, G.; Castello, G.; et al. Evaluation of selenite effects on selenoproteins and cytokinome in human hepatoma cell lines. Molecules 2013, 18, 2549–2562. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the pomegranate seeds and their extracts are available from the authors.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Costantini, S.; Rusolo, F.; De Vito, V.; Moccia, S.; Picariello, G.; Capone, F.; Guerriero, E.; Castello, G.; Volpe, M.G. Potential Anti-Inflammatory Effects of the Hydrophilic Fraction of Pomegranate (Punica granatum L.) Seed Oil on Breast Cancer Cell Lines. Molecules 2014, 19, 8644-8660. https://doi.org/10.3390/molecules19068644
Costantini S, Rusolo F, De Vito V, Moccia S, Picariello G, Capone F, Guerriero E, Castello G, Volpe MG. Potential Anti-Inflammatory Effects of the Hydrophilic Fraction of Pomegranate (Punica granatum L.) Seed Oil on Breast Cancer Cell Lines. Molecules. 2014; 19(6):8644-8660. https://doi.org/10.3390/molecules19068644
Chicago/Turabian StyleCostantini, Susan, Fabiola Rusolo, Valentina De Vito, Stefania Moccia, Gianluca Picariello, Francesca Capone, Eliana Guerriero, Giuseppe Castello, and Maria Grazia Volpe. 2014. "Potential Anti-Inflammatory Effects of the Hydrophilic Fraction of Pomegranate (Punica granatum L.) Seed Oil on Breast Cancer Cell Lines" Molecules 19, no. 6: 8644-8660. https://doi.org/10.3390/molecules19068644
APA StyleCostantini, S., Rusolo, F., De Vito, V., Moccia, S., Picariello, G., Capone, F., Guerriero, E., Castello, G., & Volpe, M. G. (2014). Potential Anti-Inflammatory Effects of the Hydrophilic Fraction of Pomegranate (Punica granatum L.) Seed Oil on Breast Cancer Cell Lines. Molecules, 19(6), 8644-8660. https://doi.org/10.3390/molecules19068644